בשלב הזה הקורס עובר מגסטרולציה לאורגנוגנזה.
יוער כי יש ביניהן חפיפה מסוימת: עוד לפני שהגסטרולציה מסתיימת, מתחילים בד בבד תהליכים ראשונים של יצירת איברים ומערכות.
נתמקד בשתי מערכות מודל:
| מערכת | מה נלמד בה |
|---|---|
| מערכת העצבים | יצירת צינור העצבים בתהליך נוירולציה, ובהמשך התמיינות נוירונים בתהליך נוירוגנזה |
| סומיטים | יצירת סומיטים מה־Paraxial mesoderm, ובהמשך התמיינותם לחוליות, שרירים, דרמיס וגידים |
העיקרון המרכזי דומה למה שנלמד בגסטרולציה: תאים ורקמות מתפתחים בעזרת מידע שהם מקבלים מהרקמות שסביבם, בעיקר דרך סיגנלים וגרדיאנטים. בשיעור הנוכחי זה ייבלוט במיוחד בשלושה הקשרים:
- הנוטוכורד משפיע על האקטודרם ומשרה את יצירת Neural Plate.
- הנוטוכורד, ה־Floor plate, ה־Roof plate, ה־Surface ectoderm וה־Lateral plate mesoderm משפיעים על התמיינות צינור העצבים והסומיטים.
- ה־Tailbud (בעברית: ״ניצן הזנב״), המזודרם הפרה־סומיטי, והשעון הסגמנטלי קובעים מתי ואיפה ייווצר סומיט חדש.
נוטוכורד ואינדוקציית ה־Neural Plate
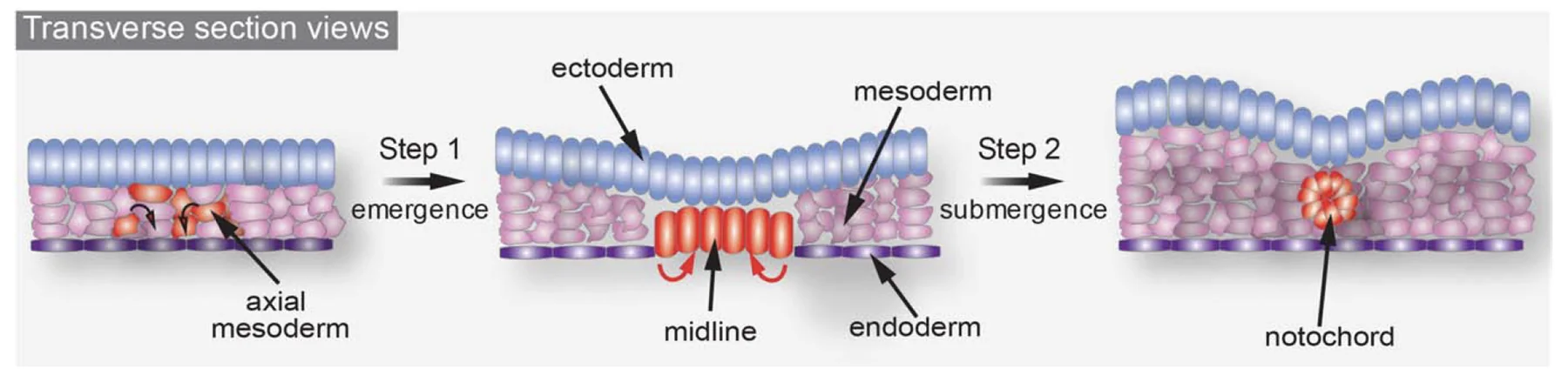
בסוף הגסטרולציה נוצרת רצועת Axial mesoderm בקו האמצע (midline), ממנה נוצר הנוטוכורד (Notochord).
הנוטוכורד מתחיל להיווצר בצד האנטריורי. בזמן שה־Node נע לכיוון פוסטריורי, תאים חדשים נשארים מאחור, נכנסים פנימה לשכבת המזודרם, ומאריכים את הנוטוכורד לאורך הציר.
הנוטוכורד משרה את יצירת מערכת העצבים המרכזית; האקטודרם שנמצא מעליו (ויוצר אותה), מקבל אינדוקציה והופך ל־Neural Plate. לכן:
- מעל הנוטוכורד נוצר Neural Plate.
- במקום שבו אין נוטוכורד, לא תתחיל יצירה תקינה של מערכת העצבים המרכזית.
זו הסיבה שיצירת ה־Axial mesoderm והנוטוכורד בסוף הגסטרולציה היא תנאי לתחילת האורגנוגנזה של מערכת העצבים.
התחומים הראשוניים באקטודרם
כשמסתכלים על האקטודרם מהצד הדורסלי, אפשר לחלק אותו לשלושה אזורים עיקריים.
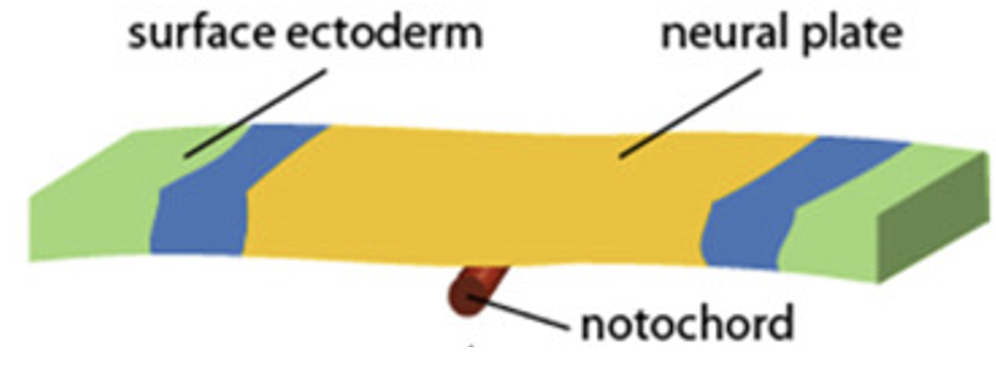
| אזור באקטודרם | מיקום | גורל עיקרי |
|---|---|---|
Neural Plate | מעל הנוטוכורד, בקו האמצע | יהפוך ל־Neural Tube |
Neural Plate Border | בין ה־Neural Plate ל־Surface ectoderm | ייתן את תאי הרכס העצבי (Neural Crest cells) |
Surface Ectoderm | לטרלית יותר | ייתן את האפידרמיס של העור |
בשלב הזה מדובר בעיקר על הצד הדורסלי של העובר (הצד הוונטרלי וצינור העיכול - Gut tube - עדיין לא מאורגנים). מתחת לנוטוכורד יש Defenitive endoderm.
נוירולציה: יצירת צינור העצבים
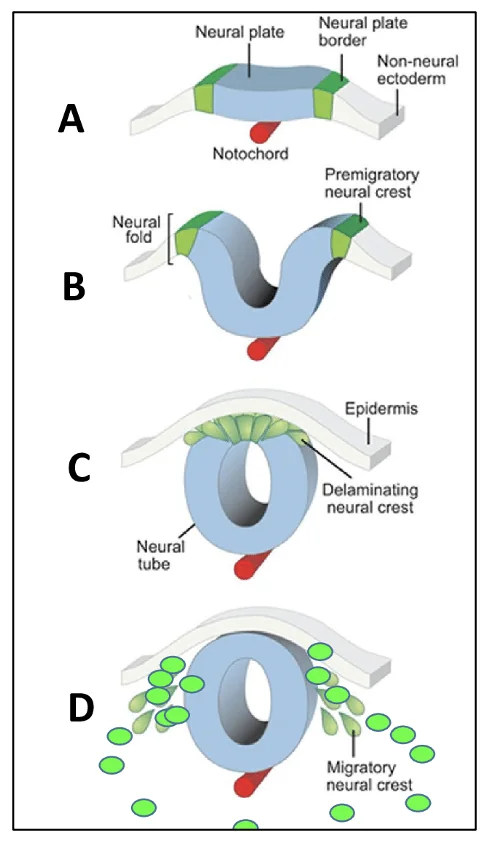
נוירולציה (Neurulation) היא התהליך שבו ה־Neural Plate מתקפל, עובר איחוי (fusion) וסגירה (closure), ולבסוף יוצר את צינור העצבים (Neural Tube).
כאמור, השלב הראשון תלוי בנוטוכורד: הנוטוכורד הוא זה שמשרה את יצירת מערכת העצבים. לכן יצירת ה־Axial mesoderm, שממנו נוצר הנוטוכורד, חשובה כל כך - בלי הנוטוכורד לא תתבצע אינדוקציית מערכת העצבים.
הרצף הכללי:
-
ה־Neural Plate נמצא מעל הנוטוכורד ומתחיל להתקפל. הקיפול נוצר בעקבות Apical constriction באזור העמוק ביותר של ה־Neural Plate, קרוב לנוטוכורד (MHP - ראו בהמשך). בגלל הכיווץ האפיקלי, משטח האקטודרם העצבי (Neuroectoderm) מתקפל ונוצרים Neural folds.
-
בהמשך נוצרים גם אזורי הכיפוף הדורסו־לטרליים DLHP (Dorsolateral Hinge Points), שעוזרים לקרב את שני ה־Neural folds זה לזה (ראו להלן). אזורי ה־Neural Plate Border מתקרבים זה לזה בהדרגה.
-
בסוף התהליך שני הצדדים מתאחים ונוצר צינור העצבים.
-
במקביל, גם ה־Surface ectoderm שמעל לצינור העצבים עובר איחוי ונסגר (החלק הלבן באיור למעלה). הוא יתפתח בהמשך לאפידרמיס של העור.
-
באזור הדורסלי של האקטודרם העצבי, תאים יוצאים מהשכבה האפיתליאלית באמצעות EMT ו־ingression, ונודדים לצדדים. אלה תאי הרכס העצבי (Neural crest), והם יתרמו בהמשך לרכיבים של מערכת העצבים הפריפרית, למשל Dorsal Root Ganglia (DRG).
חשוב להפריד בין שני דברים:
- צינור העצבים נשאר במקומו ויהפוך לחלק המרכזי של מערכת העצבים.
- תאי הרכס העצבי יוצאים מהאזור הדורסלי של האקטודרם העצבי, עוברים EMT, נודדים, ויתרמו למבנים פריפריים.
בנוסף, ה־Surface ectoderm יוצר רק את האפידרמיס של העור. הוא לא יוצר את כל העור. הדרמיס הוא שכבה מזנכימלית, ונמצאים בה מרכיבים כמו זקיקי שיער, בלוטות חלב, עצבים, בלוטות זיעה ורכיבים נוספים.
בשלב הזה מדובר בעיקר על הצד הדורסלי של העובר. הצד הוונטרלי עדיין לא מוגדר באופן מלא, ועדיין אין מערכת עיכול סגורה או gut tube. לכן הדיון כאן מתייחס בעיקר לעור, ולמבנים הדורסליים שמעל צינור העצבים.
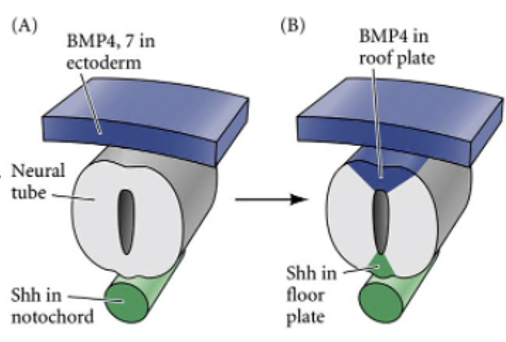
Paraxial mesoderm, ה־Roof plate וה־Floor plate
בסכמות הפשוטות רואים רק Neural Plate, Surface Ectoderm ונוטוכורד. בפועל, משני צדדי צינור העצבים נמצא גם ה־Paraxial mesoderm (הגלילים האדומים באיור למטה), שממנו ייווצרו בהמשך הסומיטים.
לאחר יצירת צינור העצבים מבחינים בו בין שני אזורים מרכזיים:
| אזור | מיקום | חשיבות |
|---|---|---|
| Roof Plate | הצד הדורסלי של צינור העצבים | קשור לסיגנלים דורסליים ולמקור תאי הרכס העצבי |
| Floor Plate | הצד הוונטרלי של צינור העצבים, מעל הנוטוכורד | קשור לסיגנלים ונטרליים כמו Shh |
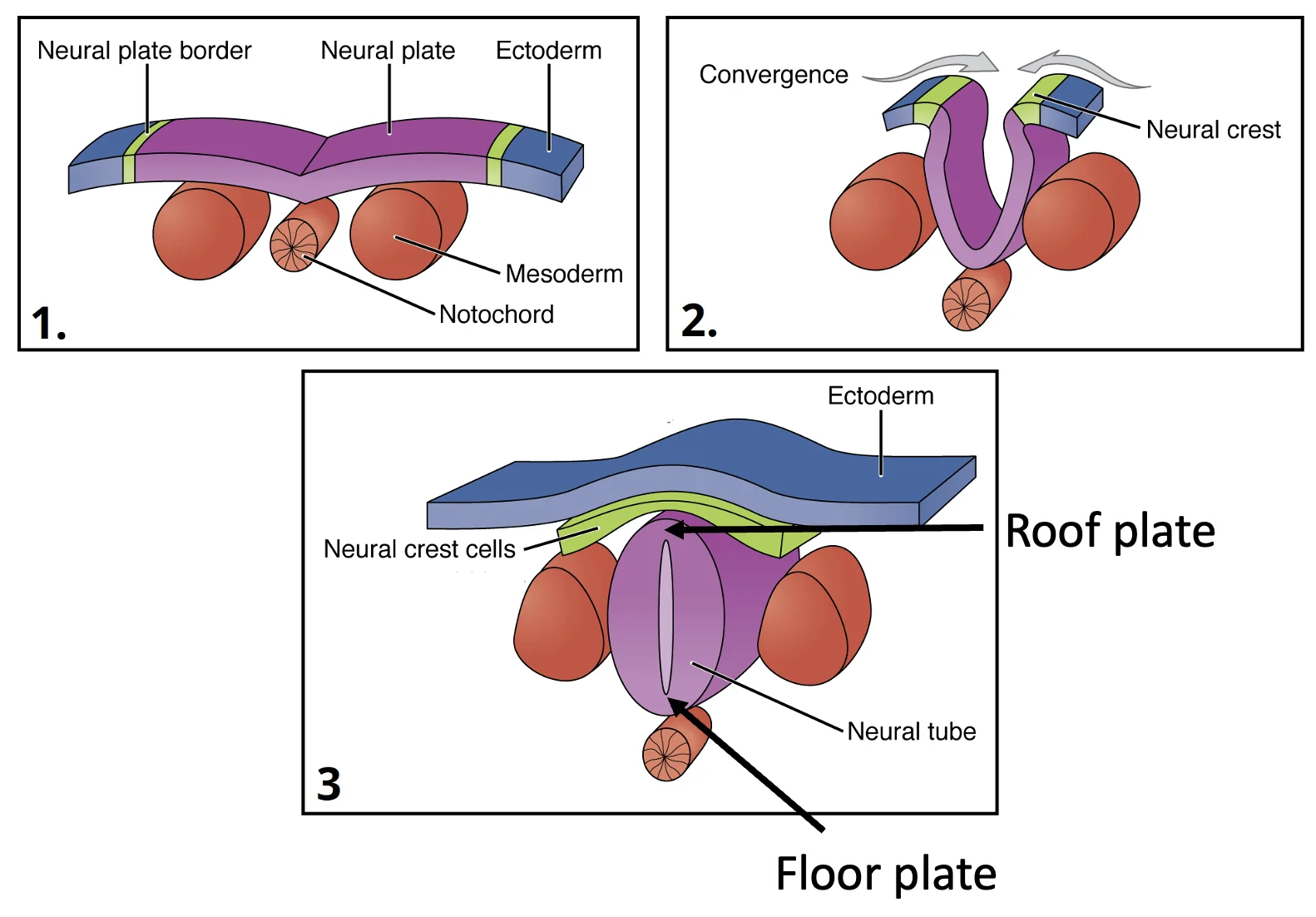 | 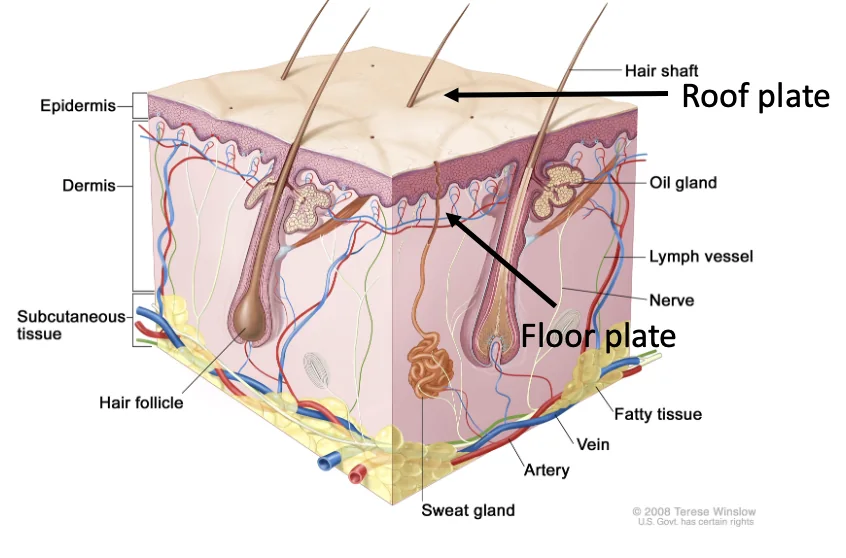 |
הנוטוכורד נמצא מתחת ל־Floor Plate. לכן שני המבנים האלה יוצרים יחד מקור חשוב לסיגנלים ונטרליים במהלך התמיינות חוט השדרה.
Hinge points ו־Apical constriction
במהלך קיפול ה־Neural Plate נוצרים אזורי כיפוף מוגדרים שבהם התאים משנים צורה.
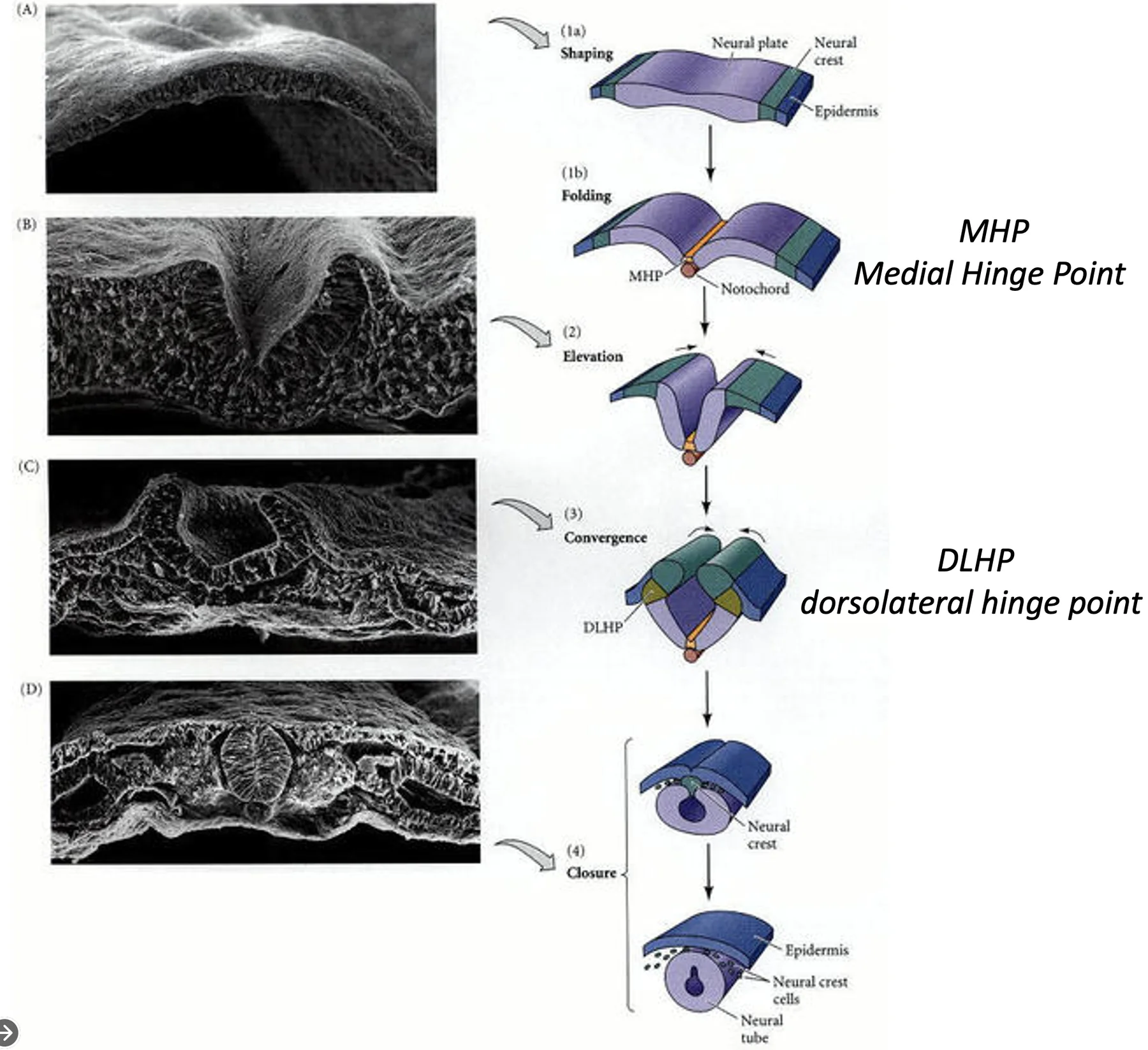
| Hinge point | מיקום | מה קורה בו |
|---|---|---|
| Medial Hinge Point (MHP) | בקו האמצע, מעל הנוטוכורד | התאים עוברים Apical constriction ויוצרים את הכיפוף הראשוני |
| Dorsolateral Hinge Points (DLHP) | בצדדים הדורסולטרליים | התאים נעשים wedge-shaped ועוזרים לקרב את שני הקפלים עד לאיחוי |
המנגנון המרכזי הוא Apical constriction: הצד האפיקלי של התא מתכווץ, התא מקבל צורת יתד, והשכבה כולה מתכופפת.
ב־MHP הכיווץ מתרחש מעל הנוטוכורד. בהמשך, נוצרים גם DLHP משני הצדדים, כדי להביא את הקפלים למגע ולסגירה.
צביעות בזמן קיפול וסגירת צינור העצבים
 | 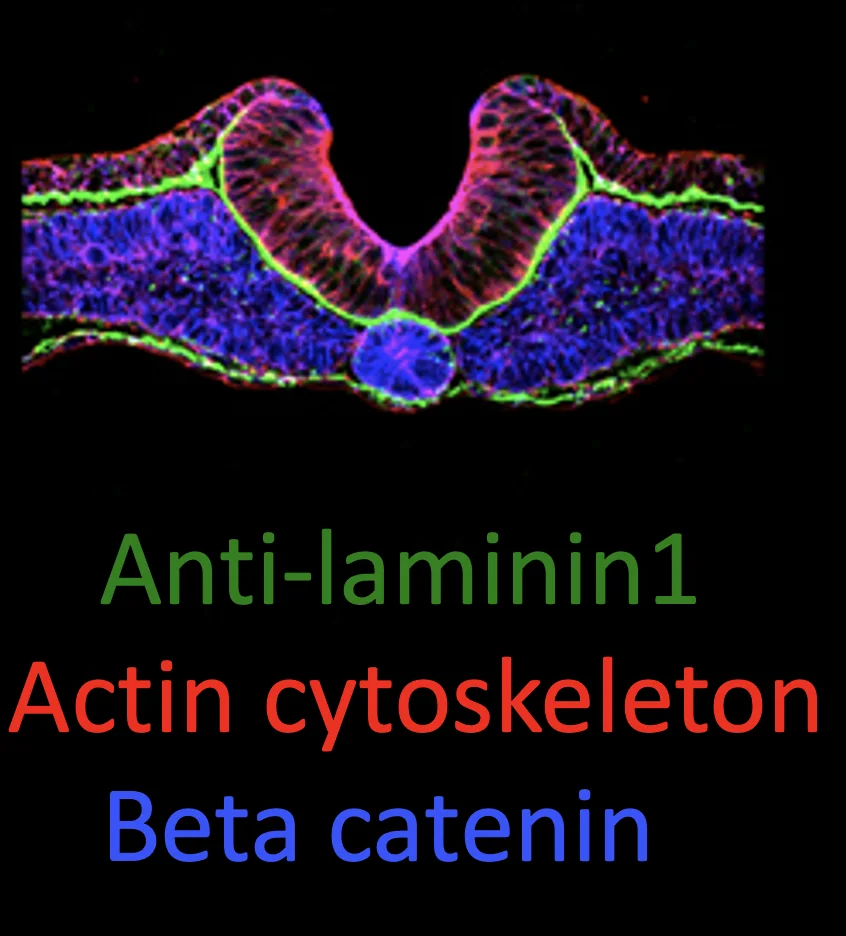 |
בחתכים דרך ה־Neural ectoderm אפשר לזהות מספר רכיבים:
| צביעה / רכיב | מה היא מדגישה |
|---|---|
Laminin | הממברנה הבזלית שעוטפת את האקטודרם |
Actin filaments | ריכוז בצד האפיקלי של תאים פולריים, מתאים למנגנון של כיווץ וקיפול |
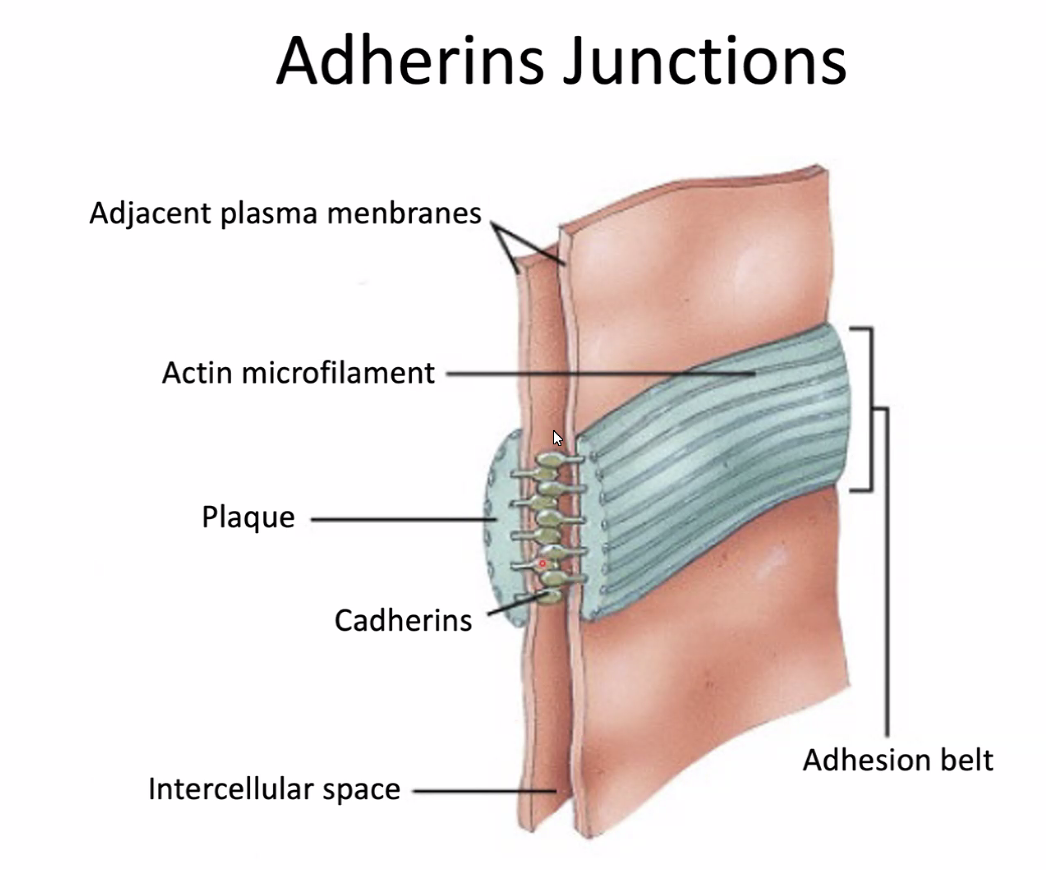
β-catenin | אזורי Adherens junctions וקשרים בין תאים |
| DAPI | גרעינים, כאשר הכחול בתמונה הוא DAPI |
בנוירולציה, הממברנה הבזלית נשארת שלמה בזמן הקיפול. זו נקודה חשובה ביחס לשיעור הקודם: ב־Primitive streak, פירוק מקומי של הממברנה הבזלית היה תנאי ל־Ingression; כאן, לעומת זאת, הממברנה השלמה עוזרת לשמור על מבנה ופולריות בזמן קיפול האפיתל.
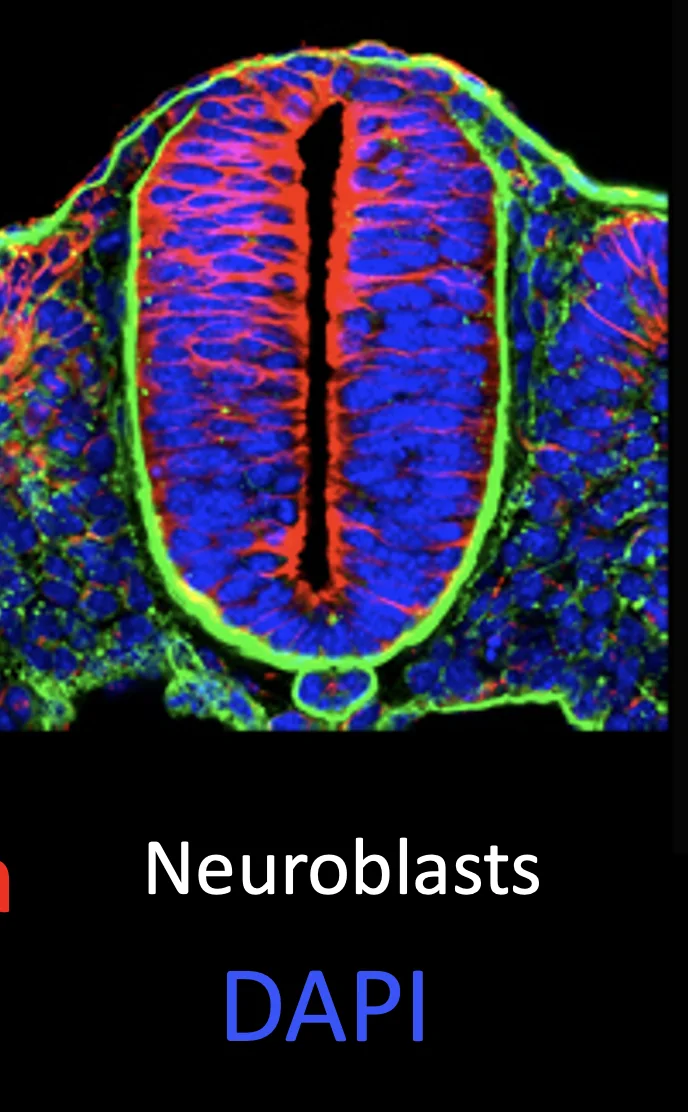
לאחר האיחוי מתקבל צינור העצבים, שבנוי מתאי עצב ראשוניים. בשלב הזה אין עדיין נוירונים בשלים, אלא נוירובלסטים (Neuroblasts) - תאים שנכנסו למסלול עצבי אבל טרם השלימו התמיינות.
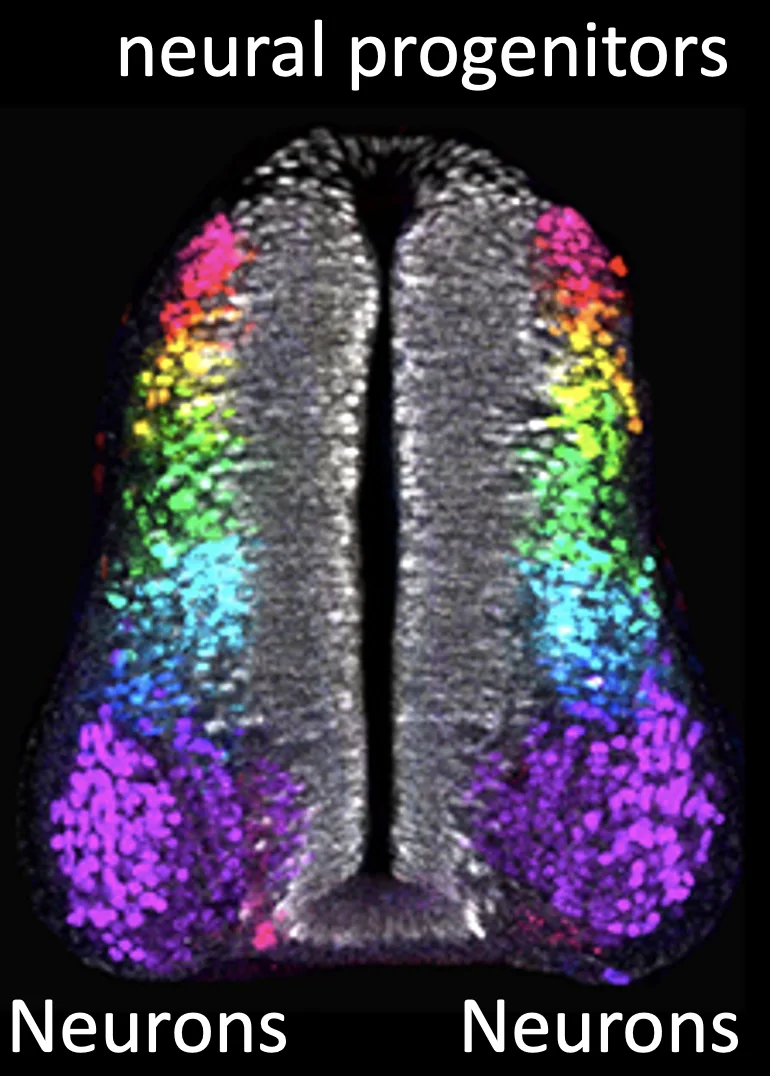
בשלב מאוחר יותר מתחילים לראות נוירונים. כאשר רואים תאים עצביים בצד הוונטרלי של חוט השדרה, זה מתאים להופעת נוירונים מוטוריים (סגול בתמונה), משום שנוירונים מוטוריים נוצרים בצד הוונטרלי.
נוירולציה לעומת נוירוגנזה
| תהליך | מה קורה בו | אופי התהליך |
|---|---|---|
| נוירולציה | Neural Plate מתקפל ונסגר לצינור העצבים | תהליך מורפוגנטי - יצירת צורה |
| נוירוגנזה | נוירובלסטים מתמיינים לנוירונים ושולחים אקסונים | תהליך התמיינות |
סגירת צינור העצבים, Neuropores ומומים בצינור העצבים
לאחר יצירת צינור העצבים, חלקו הקדמי מתרחב ומתחיל ליצור את אזורי המוח: המוח הקדמי, המוח האמצעי והמוח האחורי (Forebrain, Midbrain ו־Hindbrain, בהתאמה).
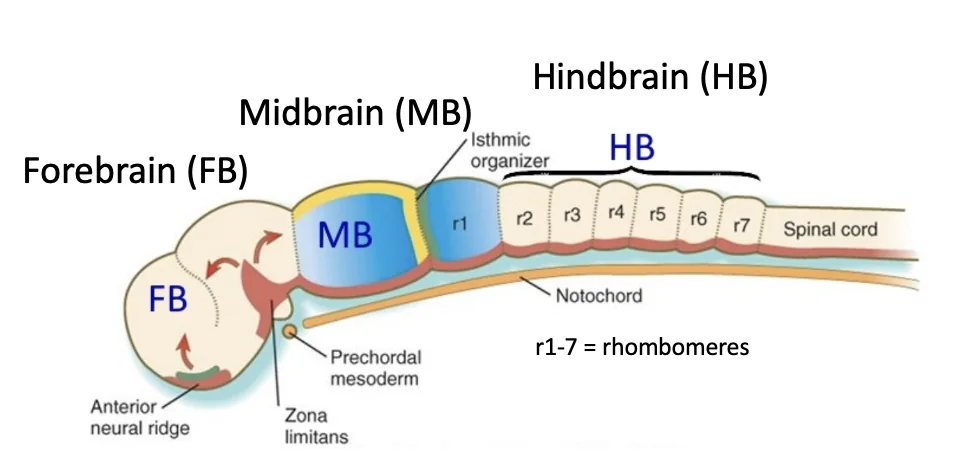
במוח האחורי יש חלוקה לשבעה Rhombomeres. לכאורה אין צורך לזכור כאן את המספרים, אלא להבין שהחלק הקדמי של צינור העצבים לא נשאר צינור אחיד: הוא עובר התנפחות, חלוקה אזורית והתמיינות. בהמשך (מצד שמאל) ניתן לראות את חוט השדרה.
נוירולציה אינה מסונכרנת לאורך כל העובר
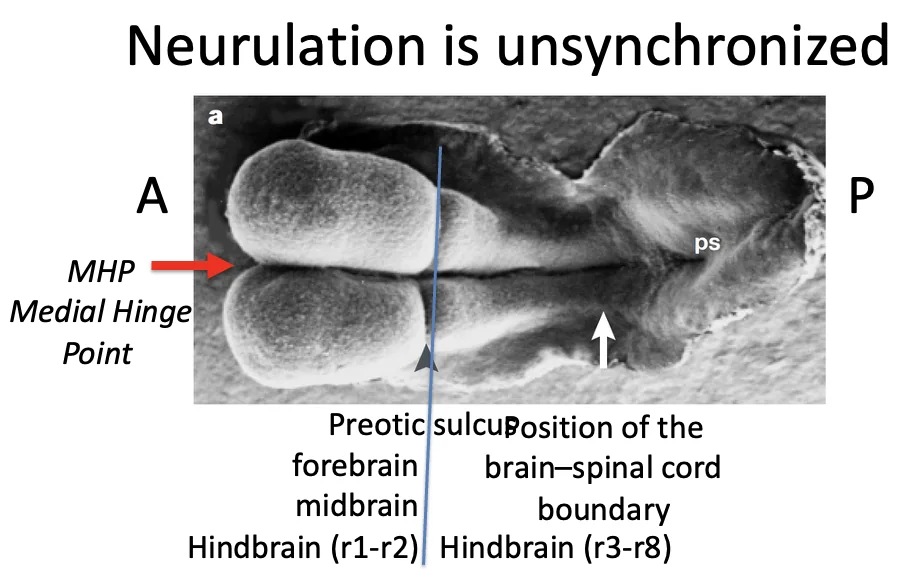
בשלבים מוקדמים העובר עסוק בעיקר ביצירת הראש והמוח; בחלק הפוסטריורי לא תמיד יש נוטוכורד מלא בשלב הזה, ולכן עדיין אין שם אינדוקציה מלאה של רקמה עצבית.
יצירת צינור העצבים מתחילה תמיד בצד האנטריורי, בהתאם למיקום הנוטוכורד.
המשמעות:
- גסטרולציה - מתקדמת בעיקר מהצד הפוסטריורי לכיוון אנטריורי.
- נוירולציה - מתחילה בעיקר בצד האנטריורי וממשיכה לכיוון פוסטריורי.
לכן, באותו עובר אפשר לראות אזורים שבהם צינור העצבים כבר נסגר, ואזורים אחרים, שבהם הקפלים עדיין פתוחים.
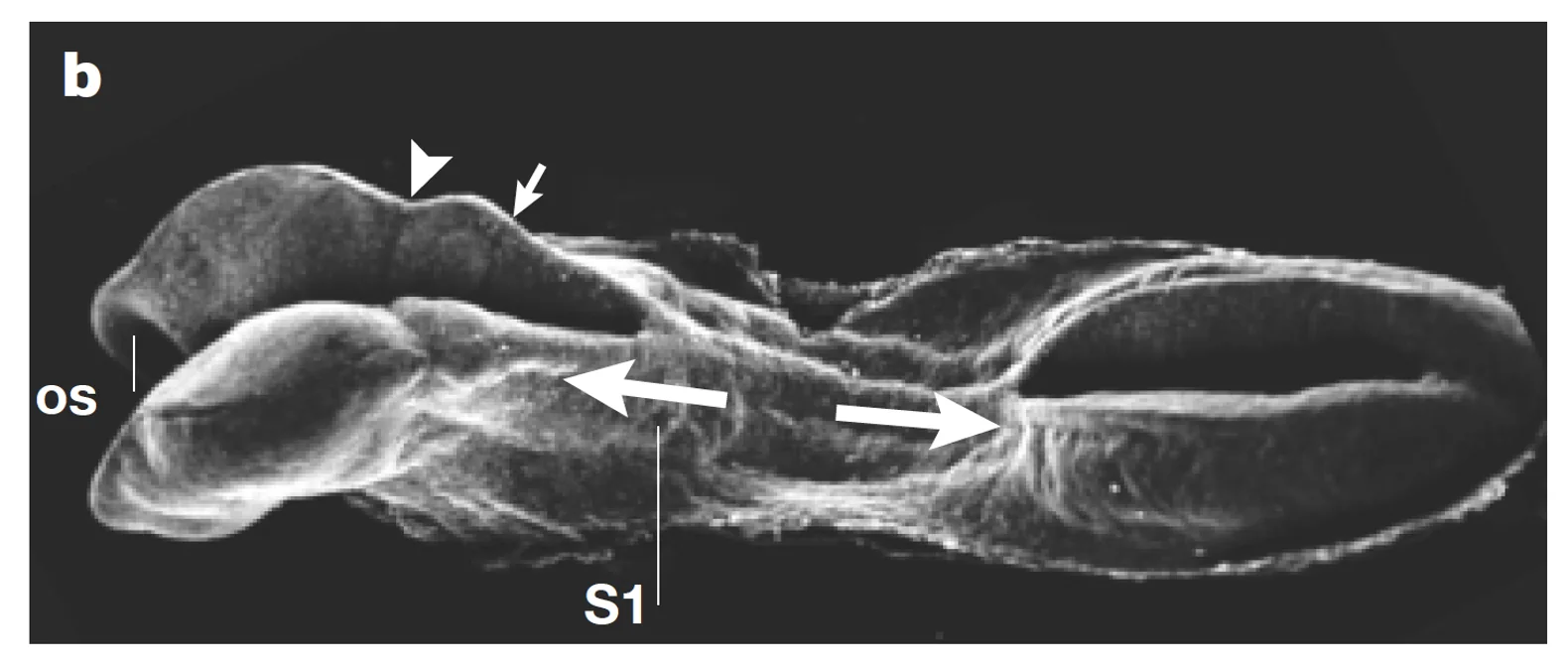
בתמונה העליונה ניתן לראות שלב מתקדם יותר.
Closure points ו־Neuropores
בעכבר מתוארים שלושה מוקדי סגירה עיקריים - אזורי איחוי במסגרת תהליך הנוירולציה. הם לא תלויים אחד בשני, והם נמצאים לאורך ציר הציר האנטריורי-פוסטריורי.
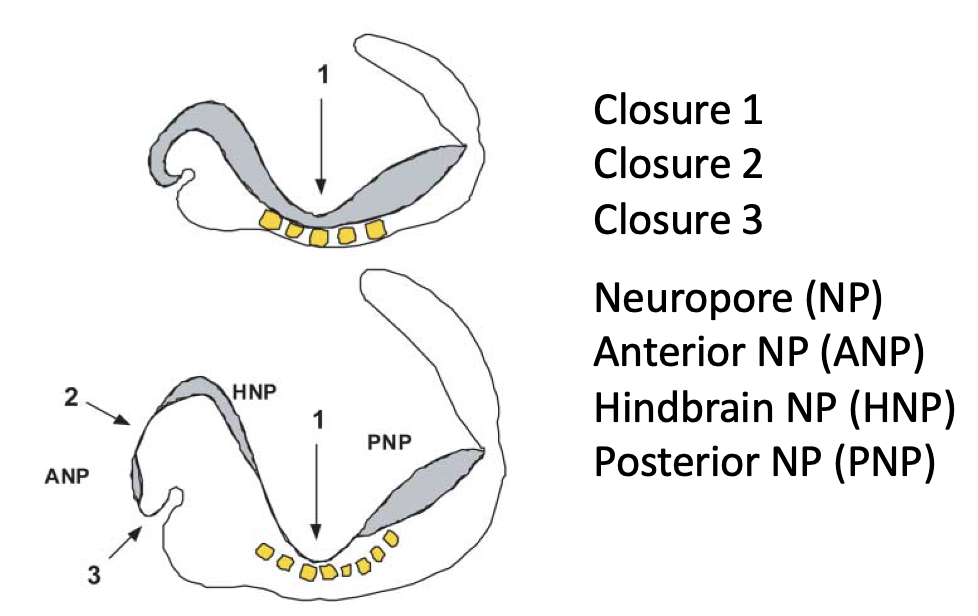
| Closure | מיקום כללי | משמעות |
|---|---|---|
| Closure 1 | אזור קדמי-אמצעי יחסית, ליד המקום שבו כבר מתחילים להופיע סומיטים | נקודת סגירה ראשונה |
| Closure 2 | אזור אנטריורי יותר | נקודת סגירה נוספת בעכבר |
| Closure 3 | קדמי מאוד | נקודת סגירה קדמית |
האזורים שנשארים פתוחים בין מוקדי הסגירה (טרם עברו “fusion”) נקראים Neuropores. ראו בתמונה מעלה.
| Neuropore | מיקום |
|---|---|
| Anterior neuropore (ANP) | בין Closure 3 ל־Closure 2 |
| Hindbrain neuropore (HNP) | בין Closure 2 ל־Closure 1 |
| Posterior neuropore (PNP) | האזור הפוסטריורי שטרם נסגר |
במצב תקין כל ה־Neuropores נסגרים בסוף התהליך. הם שלבי ביניים, לא מבנים שנשארים פתוחים.
באדם העיקרון דומה, אבל מתוארים שני מוקדי Closure מרכזיים:
- Closure 1
- Closure 3
את Closure 2, שמופיע בעכבר, לא מזהים באדם.
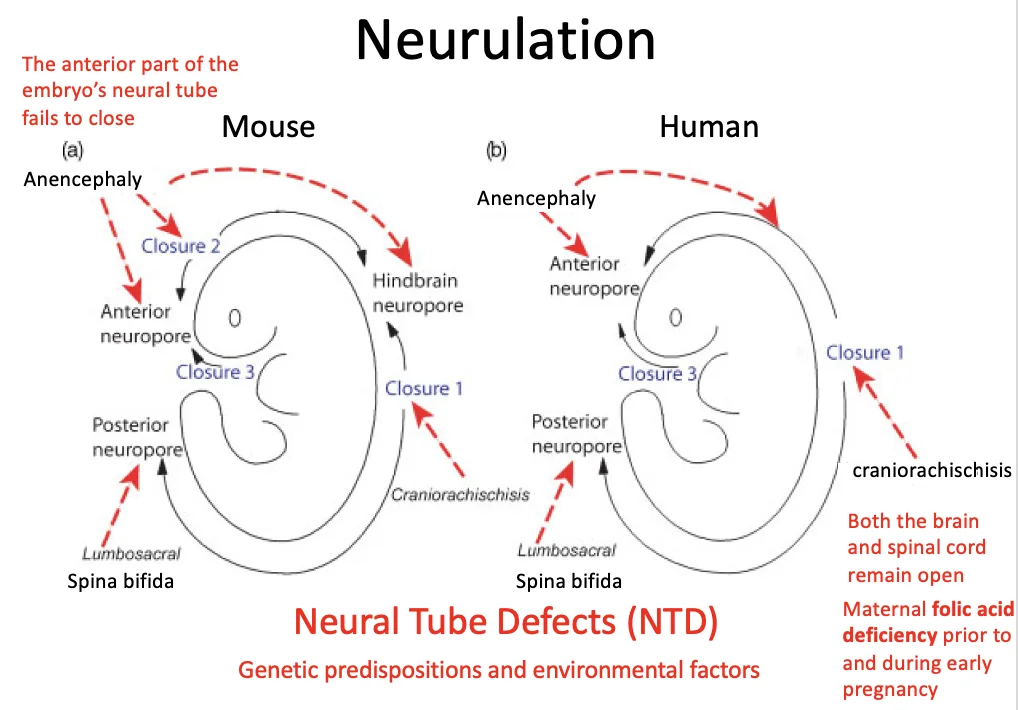
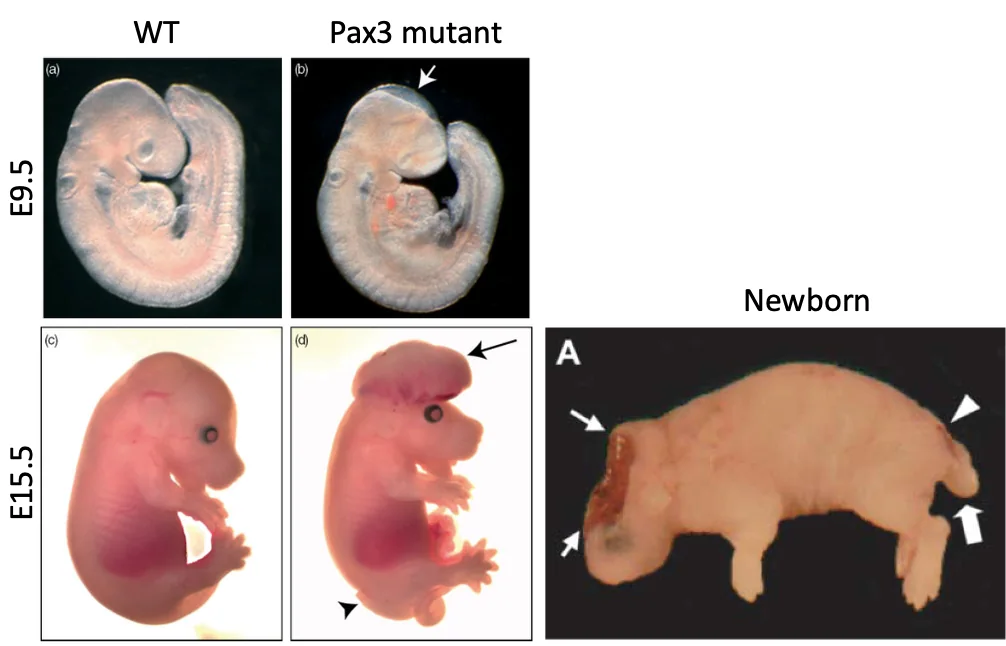
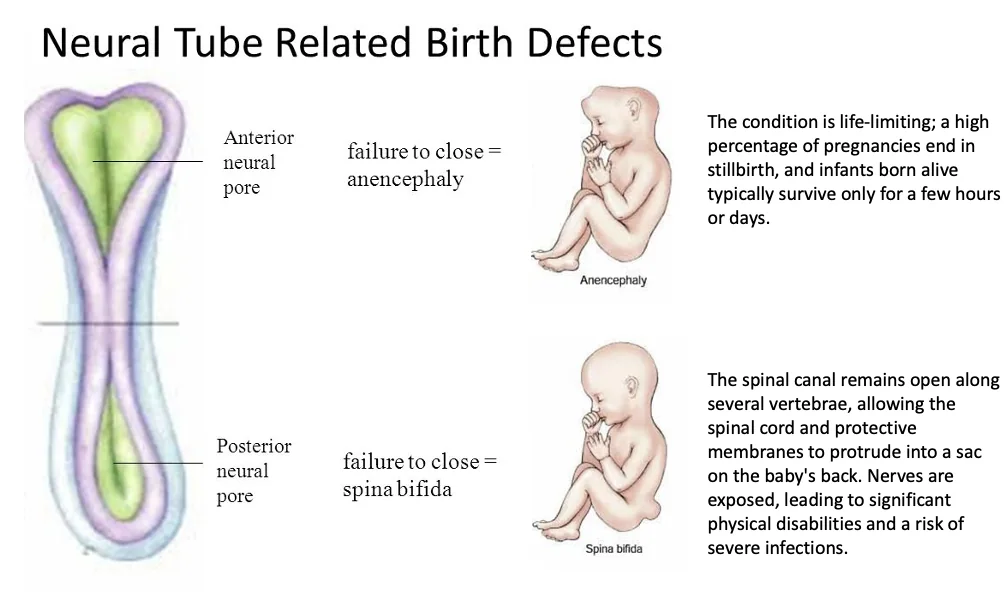
מומים בצינור העצבים (NTD)
Neural Tube Defects הם פגמים בסגירת צינור העצבים. הם יכולים לנבוע מרקע גנטי או מגורמים סביבתיים, כמו מחסור בחומצה פולית.
| פגם | מה נפגע | משמעות |
|---|---|---|
| Anencephaly | סגירת האזור האנטריורי/קרניאלי | פגיעה קשה ביצירת המוח והגולגולת; מצב חמור מאוד |
| Craniorachischisis | כשל נרחב מאוד בסגירת צינור העצבים | הצורה החמורה ביותר שהוזכרה |
| Spina bifida | סגירת האזור הפוסטריורי / spinal region | חומרה משתנה; פגיעה בחוט השדרה ובעצבים, ולכן בעיות מוטוריות אפשריות |
ב־Anencephaly הבעיה היא בסגירת האזור הקדמי. בעכבר אפשר לראות קודם כשל בסגירה הקרניאלית, ובהמשך התפתחות לא תקינה של הרקמה העצבית הקדמית.
ב־Spina bifida הבעיה היא בסגירת האזור הפוסטריורי. החומרה משתנה, אבל צפויה פגיעה בעיקר באזור חוט השדרה והעצבים שיוצאים ממנו.
נוירוגנזה של חוט השדרה
אחרי שנוצר צינור העצבים, התאים בתוכו מתחילים לעבור התמיינות עצבית. המוח מורכב יותר, ולכן נתמקד בעיקר בנוירוגנזה של חוט השדרה (Spinal Cord Neurogenesis).
Ventricular zone ו־Mantle zone
 |  |
בתוך צינור העצבים מבחינים בין אזורים תפקודיים.
| אזור | תאים | המשך |
|---|---|---|
| Mantle zone (בצדדים) | נוירונים פוסט-מיטוטיים (post-mitotic), קרי, לא מתחלקים | תאים שכבר התמיינו, כעת מייצרים אקסונים כדי להגיע לאזורים שונים בגוף |
| Ventricular zone | תאי אב (progenitor cells) שמתחלקים, רבים מהם מבטאים גורמים כמו Sox2 | מקור לתאים שימשיכו להתחלק ולתאים שיתחילו התמיינות |
בתאי האב יכולה להתרחש חלוקה א-סימטרית: תא אחד שומר על תכונות של תא אב ונשאר קרוב לחלל, ותא שני מתחיל התמיינות, נע לצידי צינור העצבים, ומצטרף ל־Mantle zone. בהמשך הוא יתחיל לשלוח אקסונים.
גרדיאנטים של Shh, BMP ו־Wnt
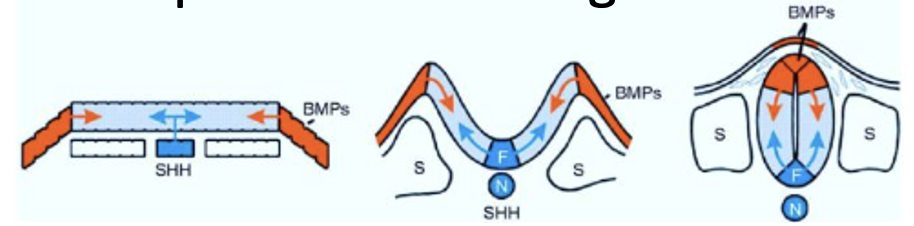
ההתמיינות לאורך הציר הדורסלי-ונטרלי של חוט השדרה תלויה בגרדיאנטים של סיגנלים.
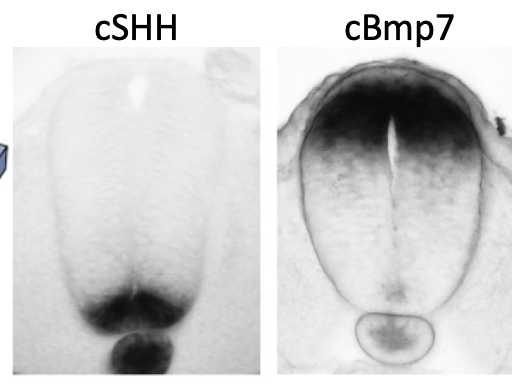 | 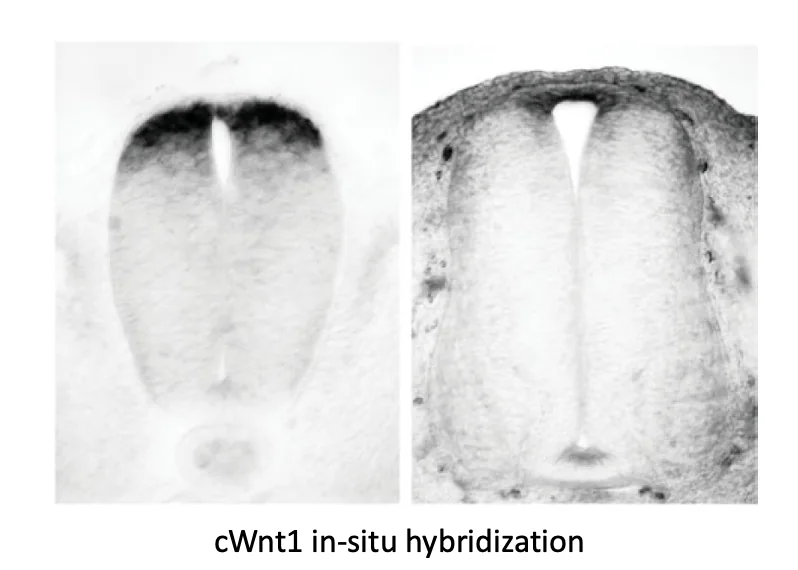 |
Sonic Hedgehog (Shh) מתבטא תחילה בנוטוכורד, ובהמשך גם ב־Floor Plate של צינור העצבים. כך נוצר מקור ונטרלי של Shh.
בצד הדורסלי פועלים סיגנלים אחרים, בעיקר BMP ו־Wnt.
העיקרון הוא שכל תא בצינור העצבים רואה שילוב אחר של סיגנלים לפי מיקומו:
| מיקום בצינור העצבים | רמות יחסיות של סיגנלים | תוצאה כללית |
|---|---|---|
| דורסלי, קרוב ל־Surface ectoderm/Roof Plate | BMP/Wnt גבוהים יותר | זהויות דורסליות |
| אזורי ביניים | שילובים שונים של Shh/BMP/Wnt | דומיינים שונים של פקטורי שעתוק |
| ונטרלי, ממש מעל הנוטוכורד/Floor Plate | Shh גבוה יותר | זהויות ונטרליות, כולל נוירונים מוטוריים |
לכן לאורך הציר הדורסלי-ונטרלי נוצרים דומיינים של פקטורי שעתוק. כל דומיין כזה תורם לספציפיקציה של אוכלוסיית נוירונים אחרת.
הוזכר גם ש־Primary cilia חשובים בהקשר של Shh signaling.
Shh והפרדה בין ימין ושמאל במוח
Shh מתבטא בקו האמצע הוונטרלי. בהקשר של המוח, הוא שומר על הפרדה בין הדומיין הימני לדומיין השמאלי (הוא לא ״מגדיר״ את ציר ימין-שמאל, אלא מונע איחוי בין שני הצדדים).
כאשר Shh נפגע, עלול להופיע איחוי בין ההמיספרות (Holoprosencephaly). Shh מתפקד כאן כמו חיץ לאורך ה־midline, ומונע איחוי בין שני הצדדים.
סומיטוגנזה: יצירת סומיטים
במקביל לנוירולציה, משני צידי צינור העצבים נמצא ה־Paraxial mesoderm. ממנו נוצרים הסומיטים.
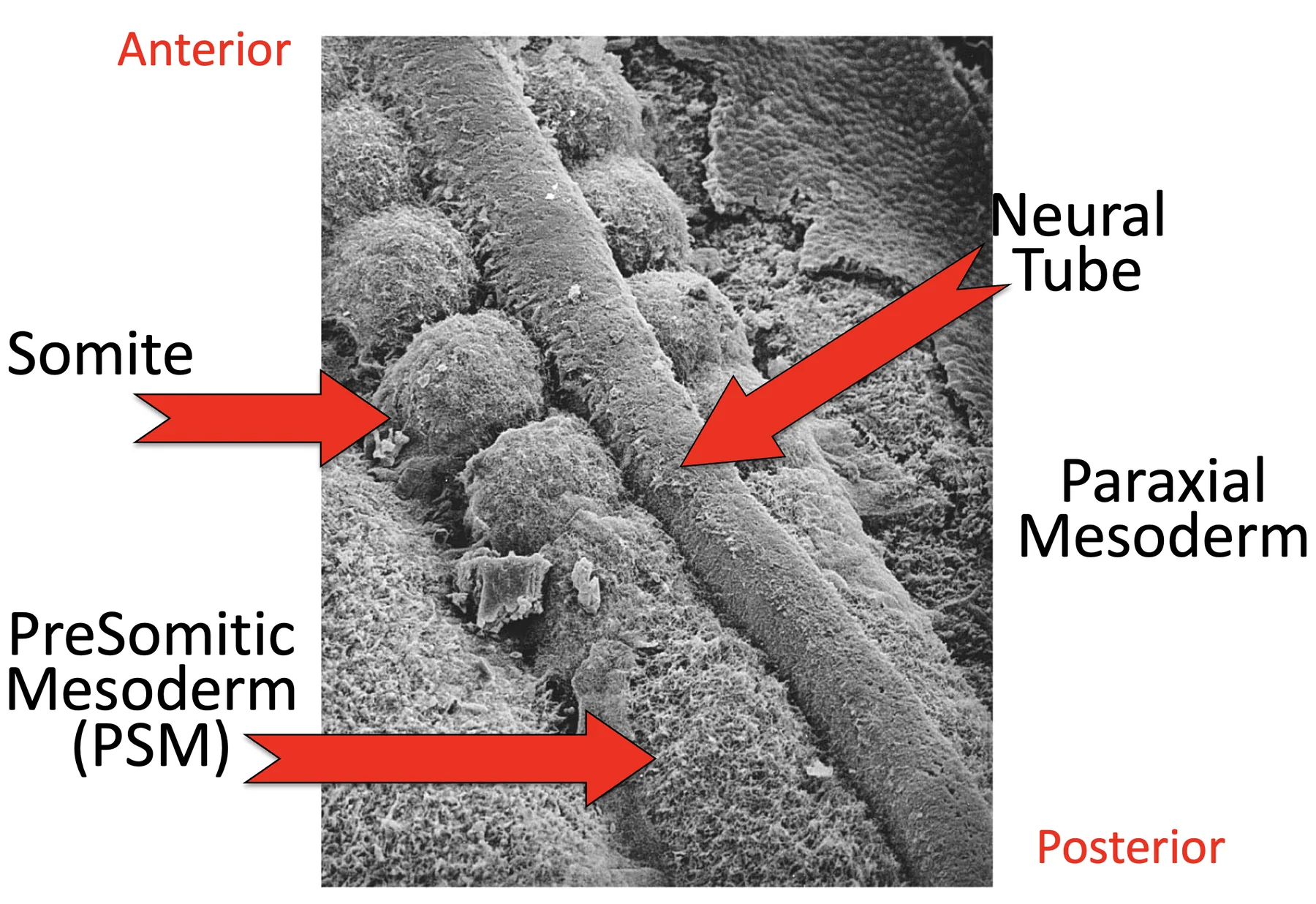 |  |
בתמונה שבה מסירים את ה־Surface ectoderm ורואים את העובר מהצד הדורסלי, אפשר לזהות:
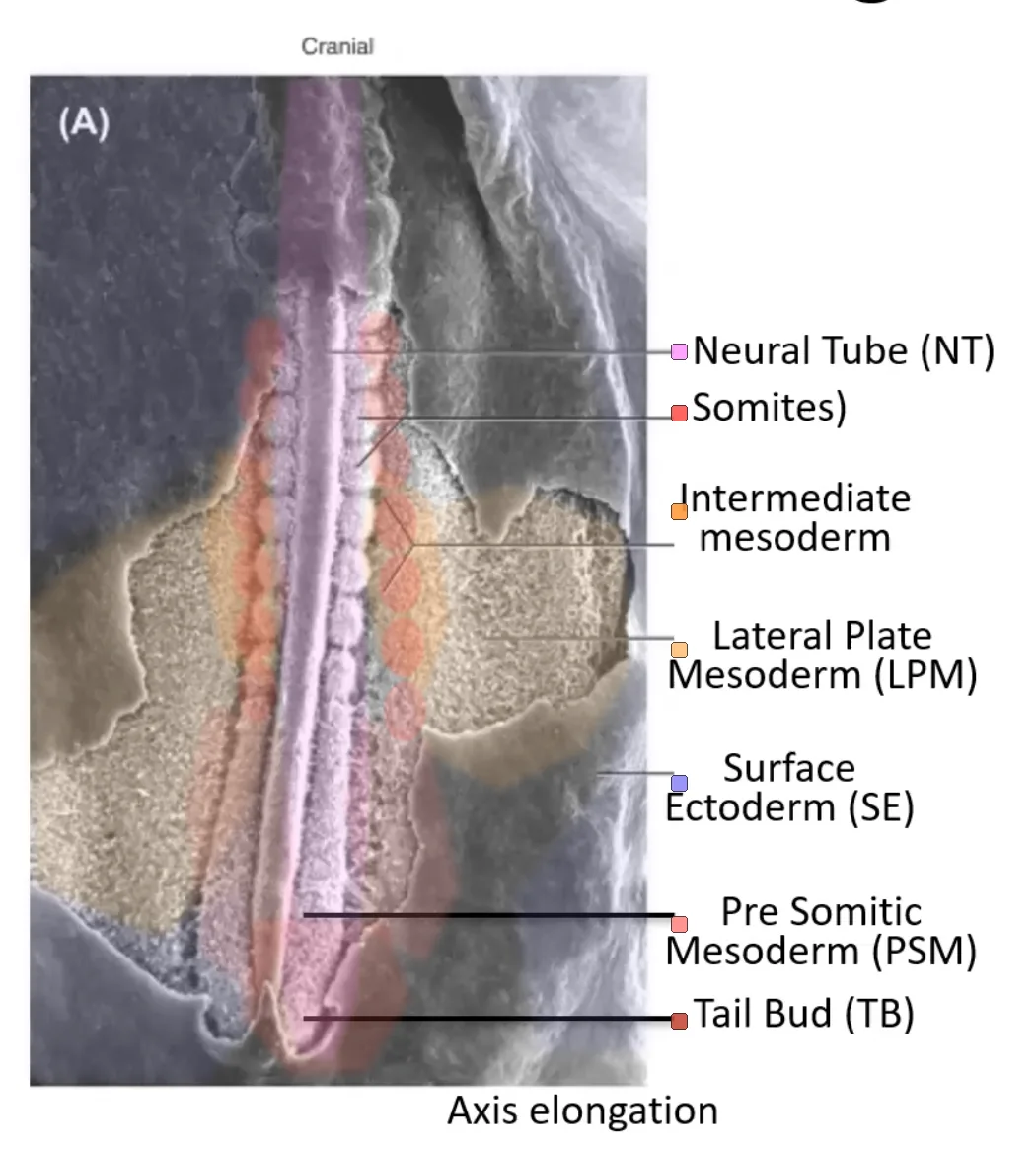 |  |
| מבנה | משמעות |
|---|---|
| צינור העצבים | הצינור העצבי שכבר נסגר באזורים מסוימים |
| הרכס העצבי | תאים שיוצאים מגג צינור העצבים ונודדים, הרכס נחשב חלק מהגג (roof) |
| אנדודרם | שכבת האנדודרם שמתחת למזודרם |
| אקטודרם | שכבת האקטודרם שמעל צינור העצבים (מקולפת בתמונה הימנית) |
| Paraxial mesoderm | משני צידי צינור העצבים |
| Somites (סומיטים) | יחידות סגמנטליות שכבר נוצרו |
| מזודרם פרה־סומיטי Presomitic Mesoderm (PSM) | האזור הפוסטריורי שטרם התפצל לסומיטים |
| Tailbud | אזור פוסטריורי שמאפשר Axis elongation |
סומיטוגנזה (Somitogenesis) היא תהליך שמתרחש ב־Paraxial mesoderm, ובו המזודרם הפרה־סומיטי מתפצל ליחידות חוזרות שנקראות סומיטים.
בזמן שה־Tailbud מאריך את הציר לכיוון פוסטריורי, בצד האנטריורי של ה־PSM נוצרים זוגות חדשים של סומיטים, משני צידי צינור העצבים.
קצב יצירת זוגות סומיטים ושעון סגמנטציה
לכל מין יש קצב יחסית קבוע של יצירת זוג סומיטים אחד לאורך הציר הקדמי-אחורי.
| מין | קצב יצירת זוג סומיטים (בערך) |
|---|---|
| עכבר | כל שעתיים |
| תרנגולת | כל שעה וחצי |
| אדם | כל חמש שעות |
| דג זברה | כל שלושים דקות |
הדיוק הזה מלמד שיש מנגנון תזמון מולקולרי - שעון סגמנטציה - שמכתיב את מחזוריות יצירת הסומיטים.
S-1, S0 ו־S1
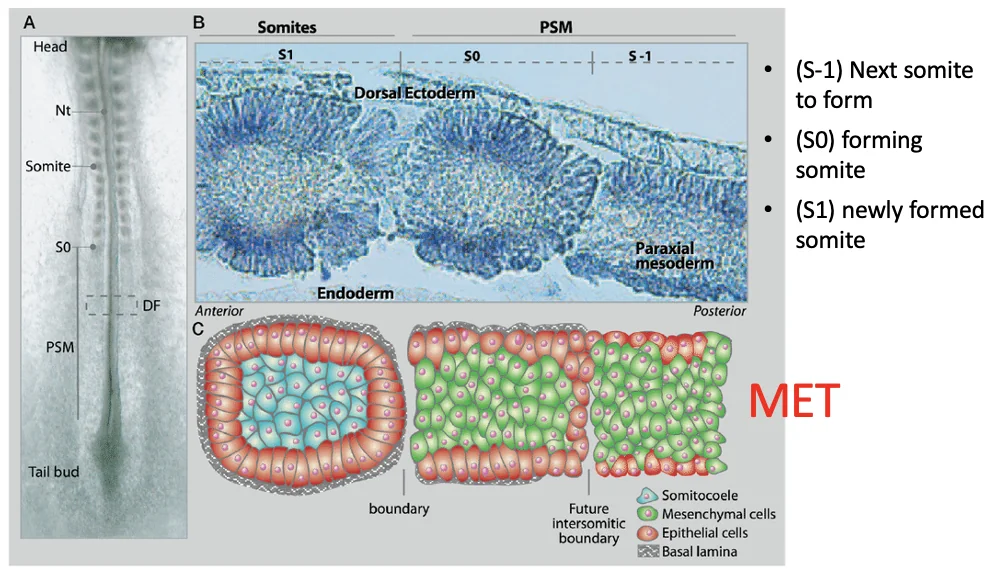
שלושה מצבים סביב יצירת סומיטים:
| סימון | משמעות |
|---|---|
| S-1 | אזור ב־PSM שכבר מיועד להפוך לסומיט, אבל עדיין לא נפרד |
| S0 - סומיט מספר אפס | סומיט שנמצא ממש בתהליך היווצרות/התפצלות |
| S1 | סומיט שכבר נוצר קודם לכן |
במהלך יצירת סומיטים מתרחש MET: תאים מזנכימליים ב־PSM מקבלים מאפיינים אפיתליאליים ומסתדרים במבנה כדורי (ויוצרים מישור). סביב הסומיט נוצרת ממברנה בזלית (ראו תמונה), ובתאים ההיקפיים מופיעים מאפיינים אפיתליאליים כמו N-cadherin ו־Adherens junctions. במרכז נשארים תאים פנימיים שנראים מזנכימליים יותר.
זאת דוגמה נוספת למוטיב חוזר בהתפתחות: תאים עוברים הלוך ושוב בין מצב אפיתליאלי למצב מזנכימלי באמצעות EMT ו־MET.
Mesp2 כגן בקרה ראשי

באזור S-1 מופיע ביטוי מוגבל בזמן של פקטור השעתוק Mesp2; הוא מופיע בחלון קצר, מפעיל תוכנית סגמנטציה, ואז נעלם.
נתקלנו ב־Mesp2 בשיעור הקודם, בהקשר של Cardiac mesoderm ו־Eomes.
Mesp2 מתפקד כגן בקרה ראשי של יצירת סומיטים (Master Regulation Gene): הוא מפעיל את תוכנית הסגמנטציה באזור שכבר הגיע לשלב המתאים.
ניתן לראות בתמונה למעלה שלאחר ש־Mesp2 התבטבא בתאים באזור המתאים, והפעיל את תוכנית הסגמנטציה, הביטוי שלו יורד (ראו את ההבדל בין תמונה I לתמונה J).
הערות:
- סומיטים לא נוצרים באזור האנטריורי. בפרט, סומיטים אינם נוצרים באזור הראש - מקור עצמות הגולגולת אחר לגמרי.
- עמוד השדרה, הצלעות וחלק גדול משרירי השלד, דווקא כן קשורים לסומיטים.
- נוירולציה וסומיטוגנזה מתרחשות במקביל (בעת סגירת צינור העצבים כבר התפצלו כחמישה סומיטים).
התמיינות סומיטים
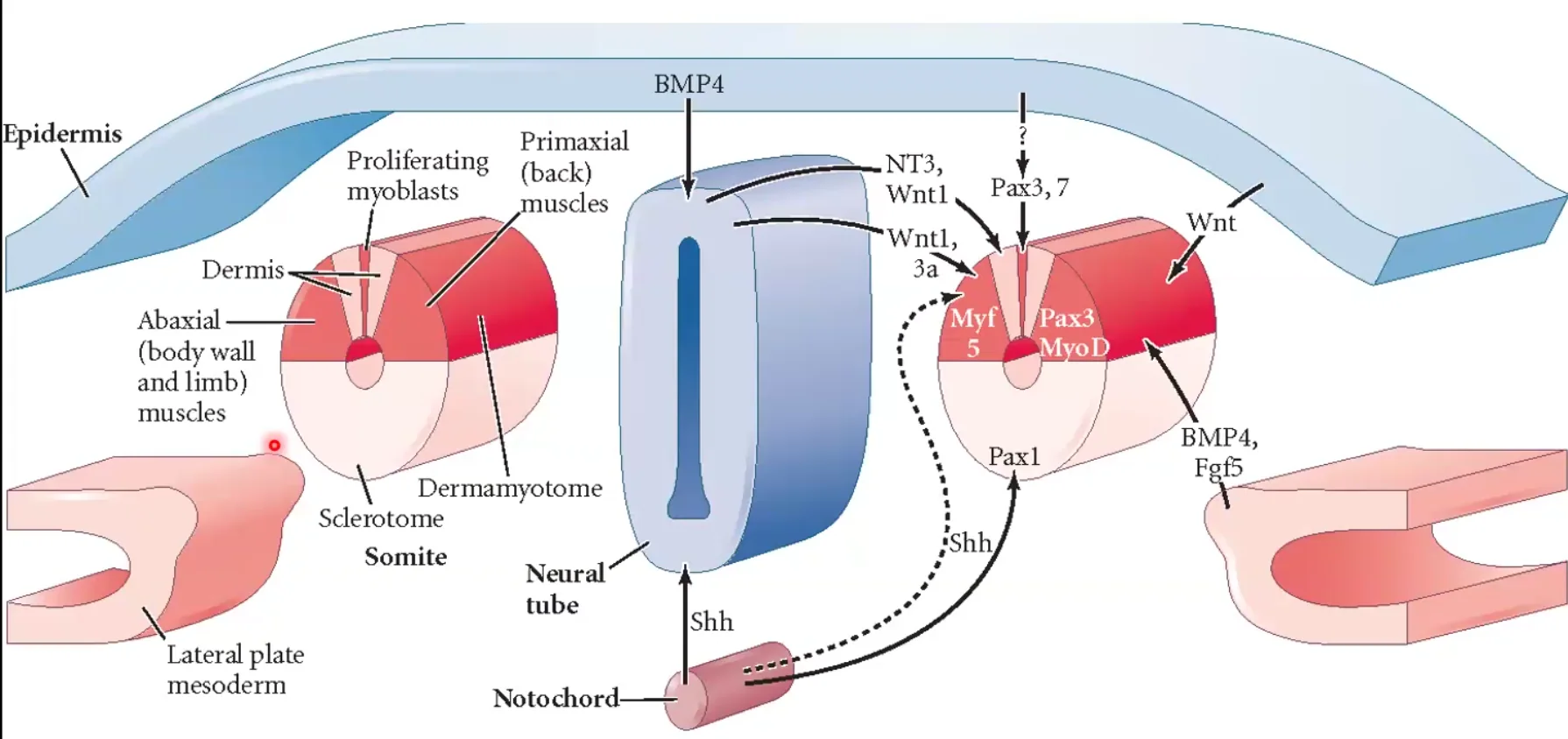
לאחר שסומיט נוצר, הוא עובר התמיינות פנימית בעצמו. כל אזור בו נחשף לסיגנלים ייחודיים מהרקמות שסביבו, ולכן מקבל גורל אחר.
 |  |
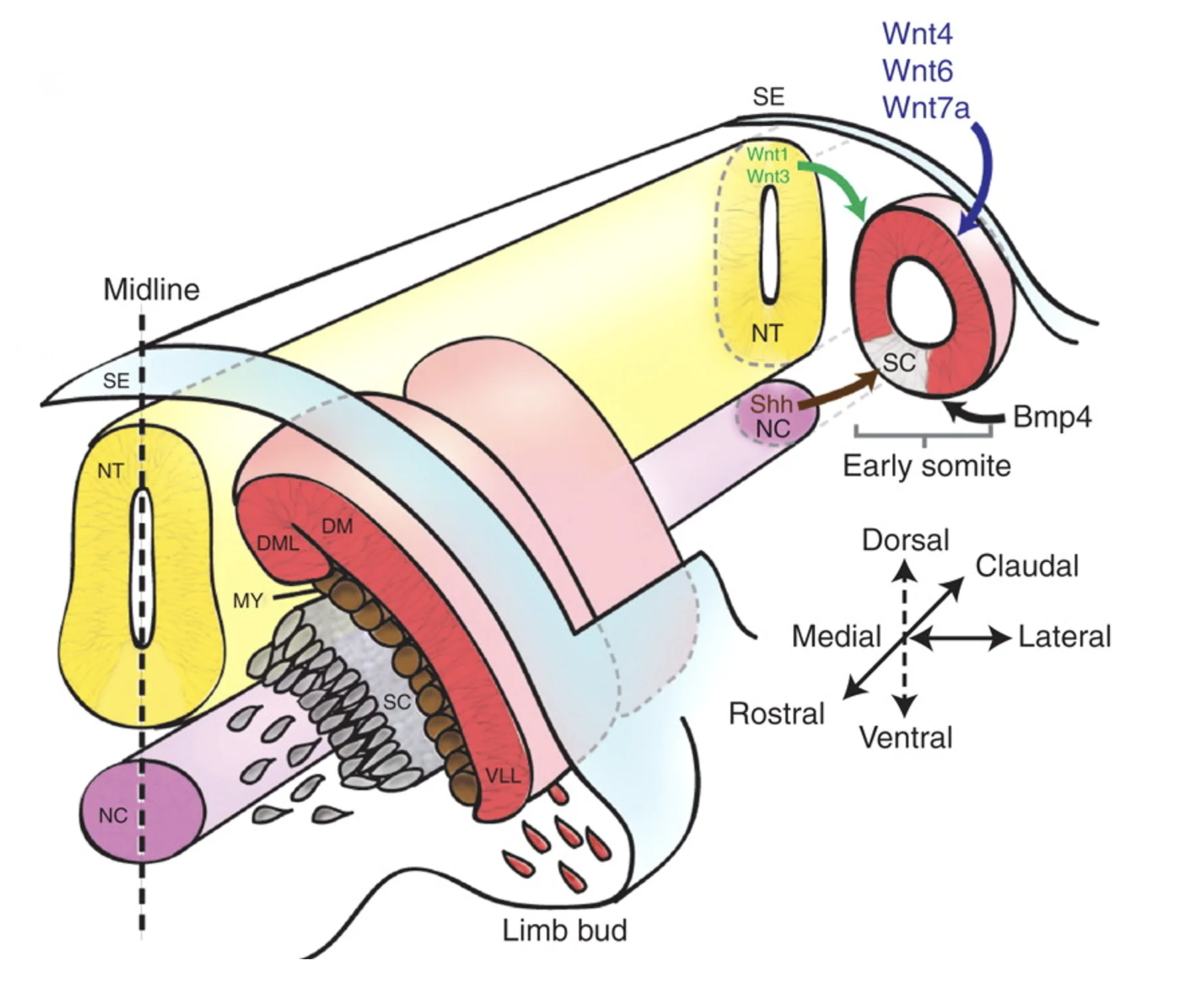
מקורות הסיגנלים סביב הסומיט:
| מקור | סיגנלים | השפעה כללית |
|---|---|---|
| Dorsal Neural Tube / Surface Ectoderm | Wnt | תורם לזהות דורסלית ולכיוון שרירי |
| Lateral Plate Mesoderm | BMP | משפיע על הצד הלטרלי ועל מצב ההתמיינות של הסומיט |
| Notochord / Floor Plate | Shh | מקדם זהות ונטרלית, בעיקר סקלרוטום |
 | 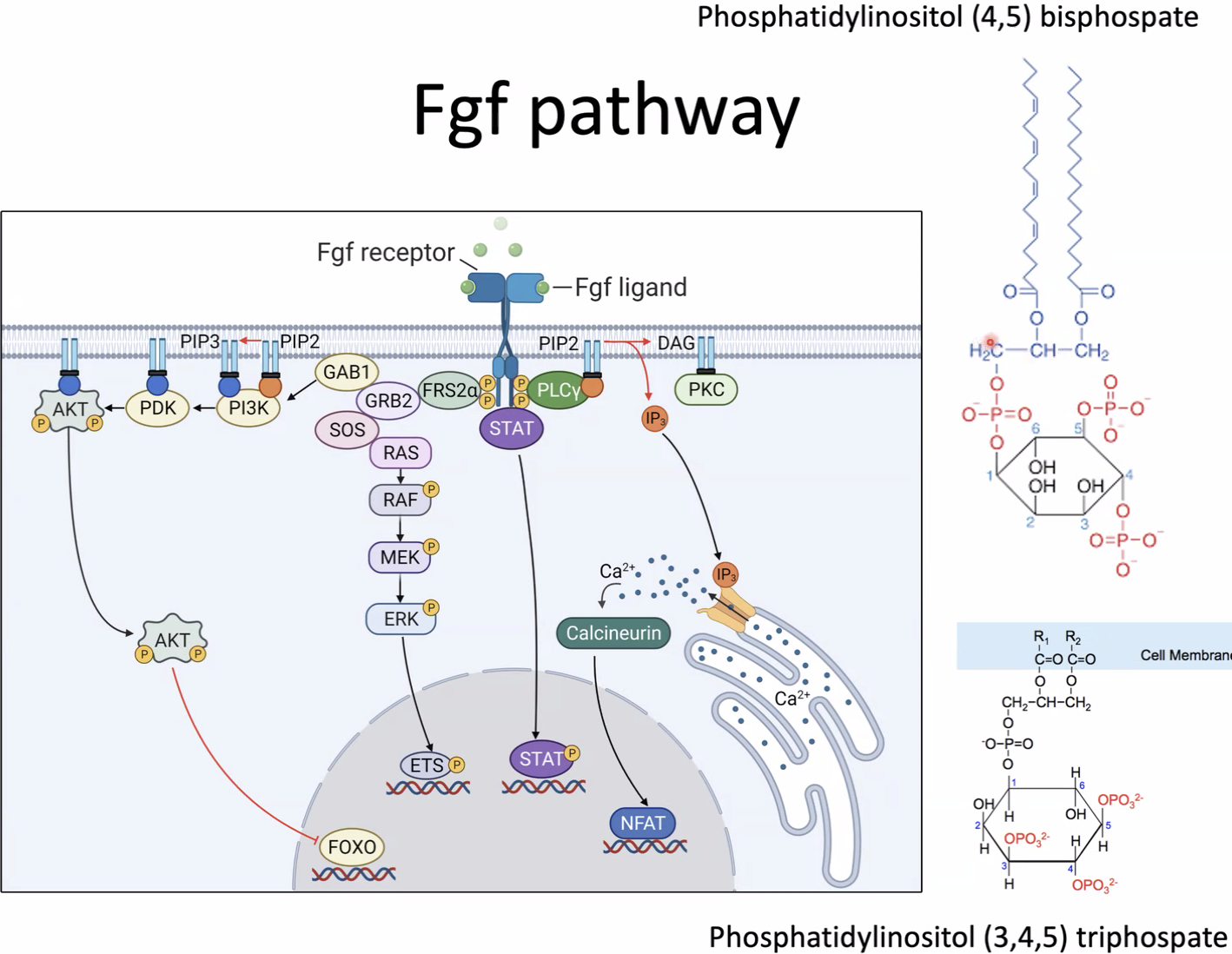 |
החלוקה הראשונית (בתמונה, החצי הדורסלי עליון, והחצי הוונטרלי תחתון):
| אזור בסומיט | מיקום | חשיפה | גורל מרכזי |
|---|---|---|---|
| דרמו־מיוטום (Dermomyotome) | חצי דורסלי | Bmp/Wnt | מקור לדרמטום ולמיוטום |
| סקלרוטום | חצי ונטרלי | Shh | חוליות, רכיבי עמוד השדרה, צלעות, ובהמשך גם מקור ל־Syndetome |
חלוקות נוספות שנראה בהמשך:
| אזור בסומיט | מיקום | גורל מרכזי |
| דרמטום | מרכיב דורסלי | דרמיס של העור |
| מיוטום (Myotome) | שכבת תאי אב שריריים | שרירי שלד |
| Syndetome | תאים שמקורם בסקלרוטום | גידים המחברים שריר לעצם |
סקלרוטום
החצי התחתון (הוונטרלי) של הסומיט הופך לסקלרוטום (Sclerotome). תאי הסקלרוטום עוברים EMT (הופכים לתאים מזנכימליים), נודדים ומקיפים את הנוטוכורד ואת צינור העצבים.
תאי הסקלרוטום הם המקור המרכזי לחוליות עמוד השדרה, לצלעות ולחלקים של השלד האקסיאלי.
דרמו־מיוטום, מיוטום ודרמטום
החצי העליון (הדורסלי) של הסומיט יוצר את הדרמו־מיוטום. ממנו נוצרים שני מרכיבים עיקריים:
| מרכיב | גורל |
|---|---|
| דרמטום | דרמיס של העור |
| מיוטום | תאי אב של שרירי שלד |
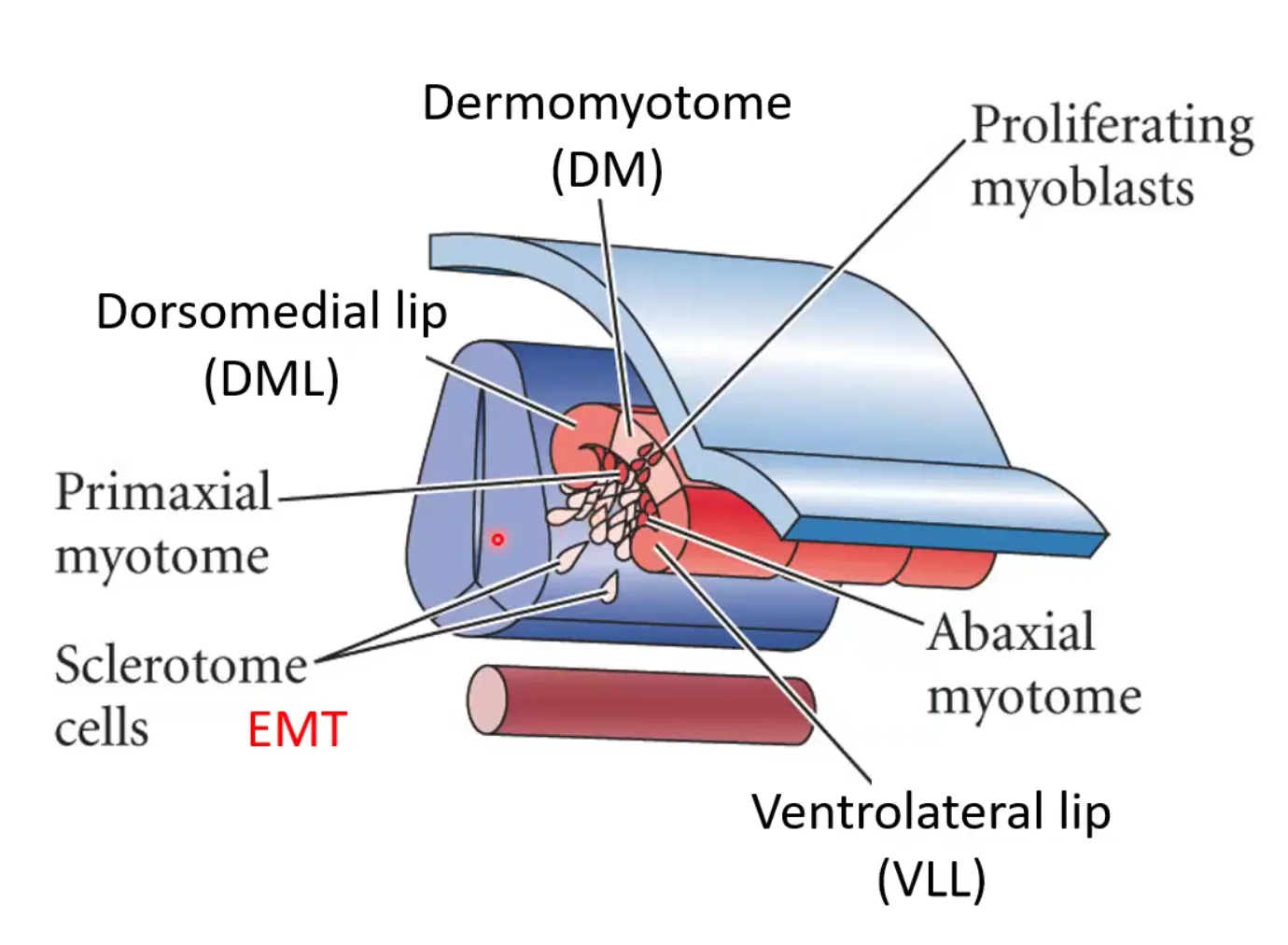
בדרמו־מיוטום מבחינים בשתי שפות, שנוצרות מכפל של תאים אפיתיליוםיים:
| אזור | המשך |
|---|---|
| Dorsomedial lip (DML) | תורם לשרירים אקסיאליים, בעיקר שרירי גב הקשורים לעמוד השדרה |
| Ventrolateral lip (VLL) | תורם לשרירים ונטרליים ולטרליים; באזור ניצני הגפיים (buds) תאים נודדים לגפה ויתרמו לשרירי הגפיים |
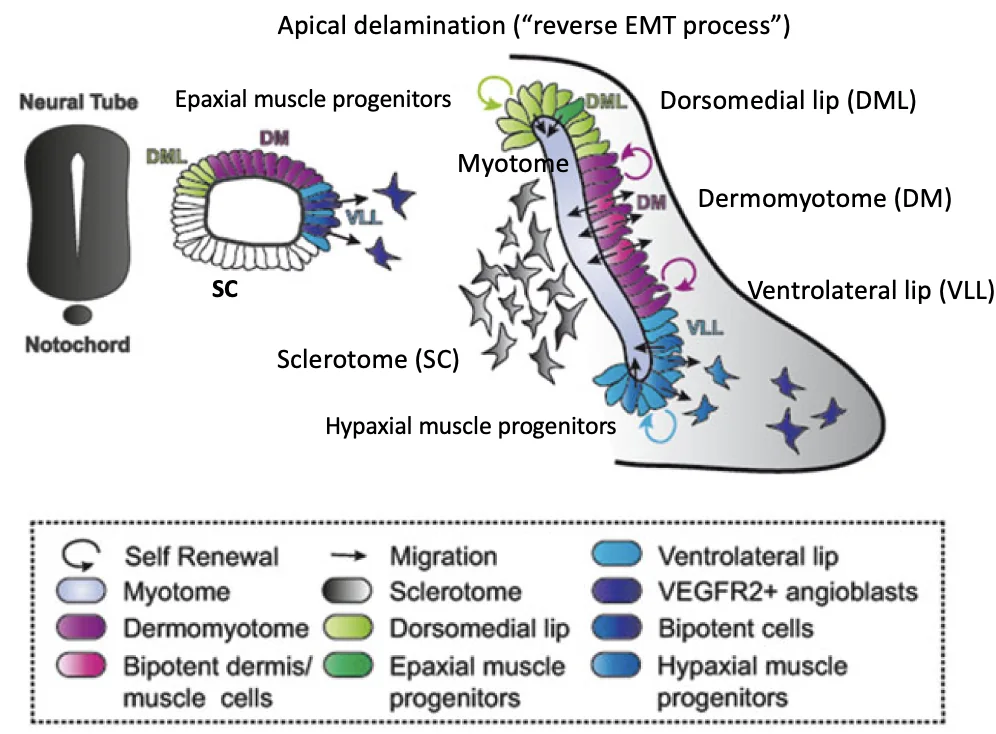
התאים בשפות (הכחול והירוק בתמונה) עוברים Apical Delamination (או "reverse EMT process") ונכנסים בין הסקלרוטום לבין הדרמו־מיוטום. כך נוצרת שכבת המיוטום, שממנה יתפתחו תאי השריר.
בשפות ה־Dermomyotome התאים עוברים תהליך המכונה apical delamination, או reverse EMT, במהלכו מתרחשת התכווצות בצד הבזלי של התאים, הגורמת להם להיפרד מהשכבה האפיתליאלית, לנוע פנימה מתחת ל־Dermomyotome וליצור את שכבת ה־Myotome, שממנה יתפתחו השרירים.
בנוסף לקצוות, גם תאים בשכבת האמצע של הדרמו־מיוטום (לא בקצוות) עוברים EMT דומה ל־Primitive streak, ונודדים פנימה ליצירת שכבת המיוטום. כך נוצרת שכבת המיוטום מכלל הדרמו־מיוטום.
התאים שנשארו הם הדרמטום, שייתרמו לדרמיס העור.
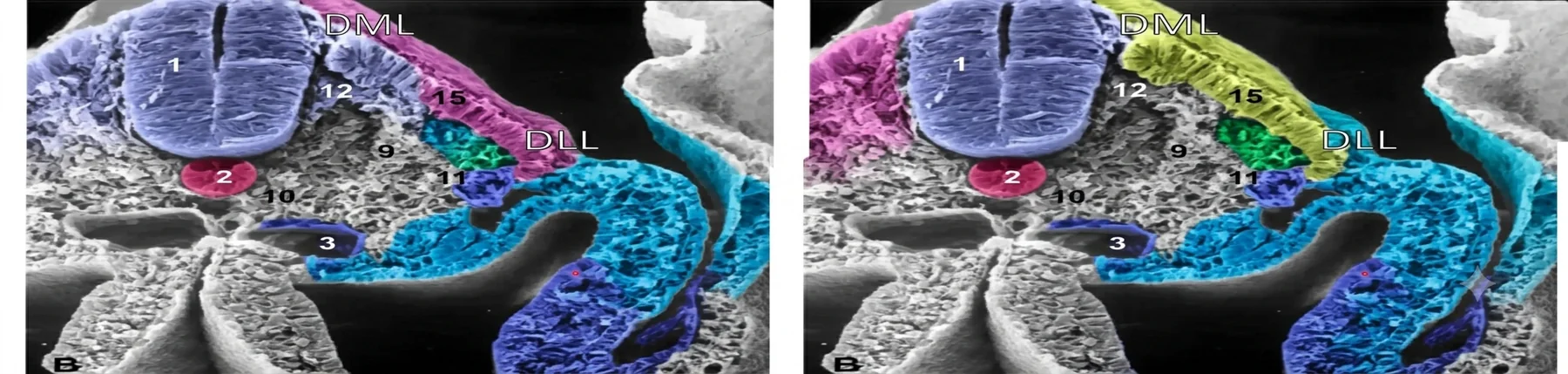
לאחר התמיינות הדרמו־מיוטום, הסומיט מחולק לשלושה אזורים
| אזור | מיקום | גורל מרכזי |
|---|---|---|
| דרמטום | דורסלי | דרמיס של העור |
| מיוטום | אמצעי | תאי אב של שרירי שלד |
| סקלרוטום | ונטרלי | חוליות, צלעות, חלקים של השלד האקסיאלי, ובהמשך גם מקור ל־Syndetome |
Syndetome
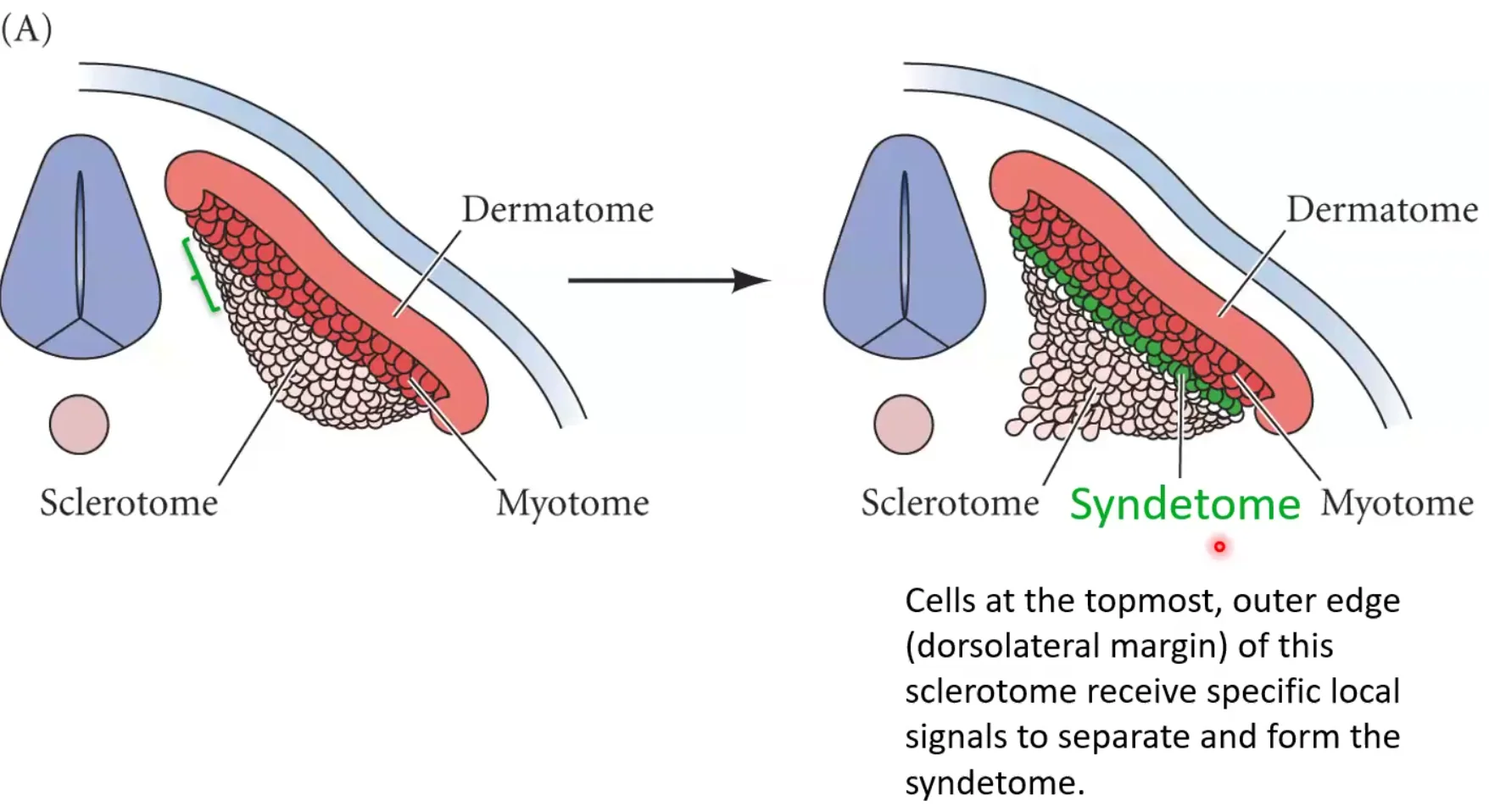
חלק מתאי הסקלרוטום מתמיינים ל־Syndetome. זה המקור לגידים (רקמת חיבור בין שריר לעצם). זה קורה בכל סומיט בנפרד. אז כל סומיט תורם לא רק לשלד ולשריר, אלא גם לחיבור המכני ביניהם.
בתמונה, ניתן לראות את העבור ״עם עומק״. בחלק הפוסטריורי (החלק ״העמוק״, ימני-עליון), נמצא סומיט צעיר שזה עתה נוצר, ובחלק האנטריורי (״שיוצא אלינו״) סומיט ותיק שכבר מתחלק.
הצד האנטריורי בתמונה הוא הצד השמאלי-תחתון, ניתן לזהות זאת לפי מצבו של הסומיט שם - הוא בוגר וניתן לזהות בו סקלרוטום, מיוטום ודרמטום.
חוליות, שרירים וגידים
יצירת עמוד השדרה
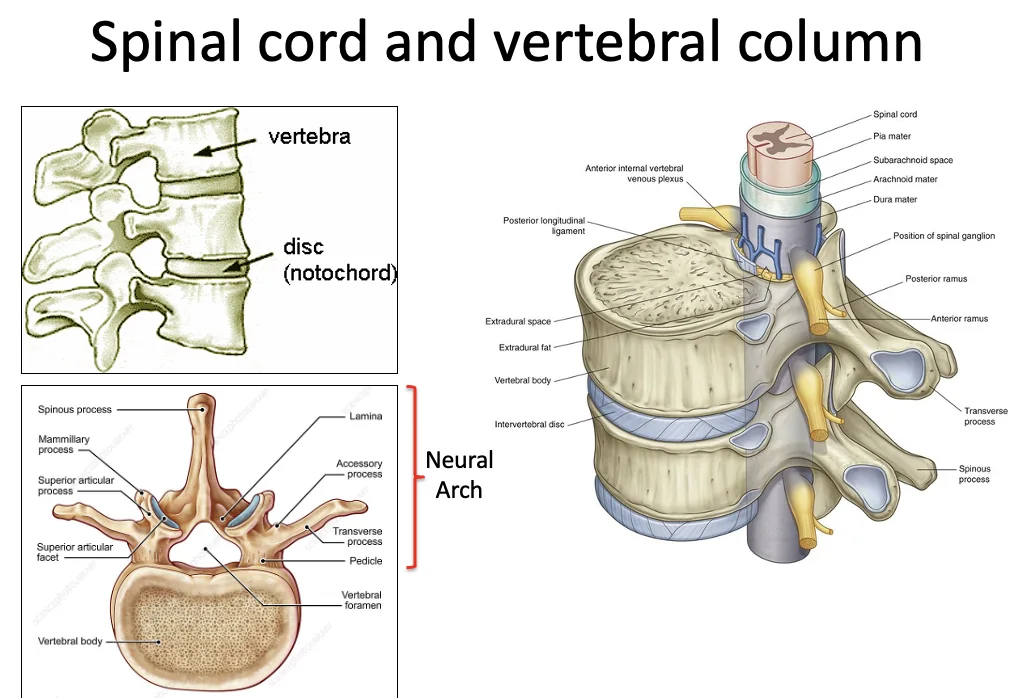
תאי הסקלרוטום נודדים סביב הנוטוכורד וצינור העצבים. בהמשך הם מתארגנים סביבם ויוצרים את מבנה החוליות.
הארגון הזה מסביר את מיקום חוט השדרה: חוט השדרה עובר בתוך תעלה שנוצרת על ידי החוליות (בחלק הדורסלי שלהן), והחוליות מתארגנות סביבו.
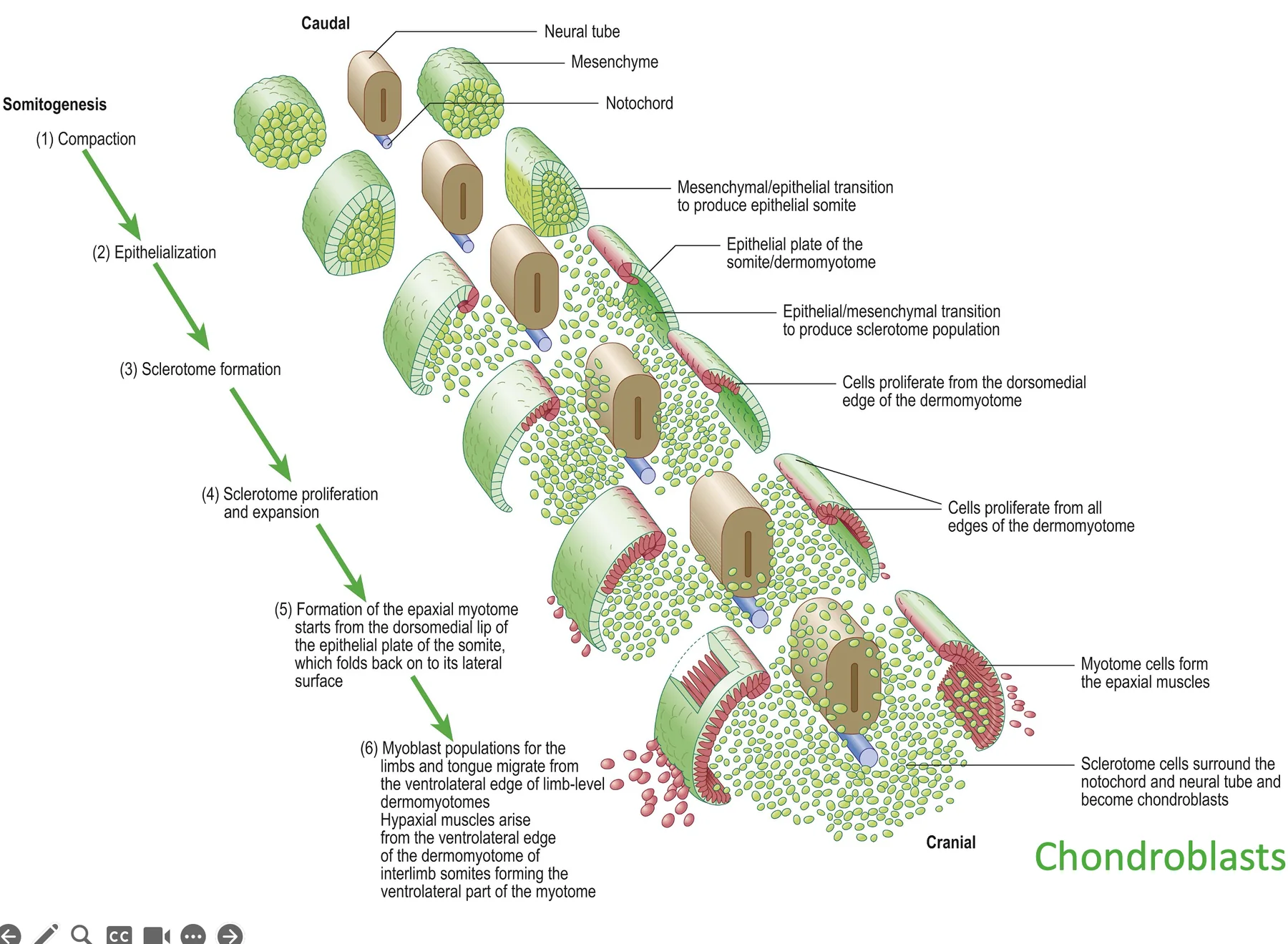
הנוטוכורד כמעט נעלם בהמשך, אבל תורם ליצירת הדיסק הבין־חולייתי. התאים שמסתדרים סביב הנוטוכורד וצינור העצבים הופכים לכונדרובלסטים (Chondroblasts), שמייצרים את סחוס החוליות.
מיוגנזה
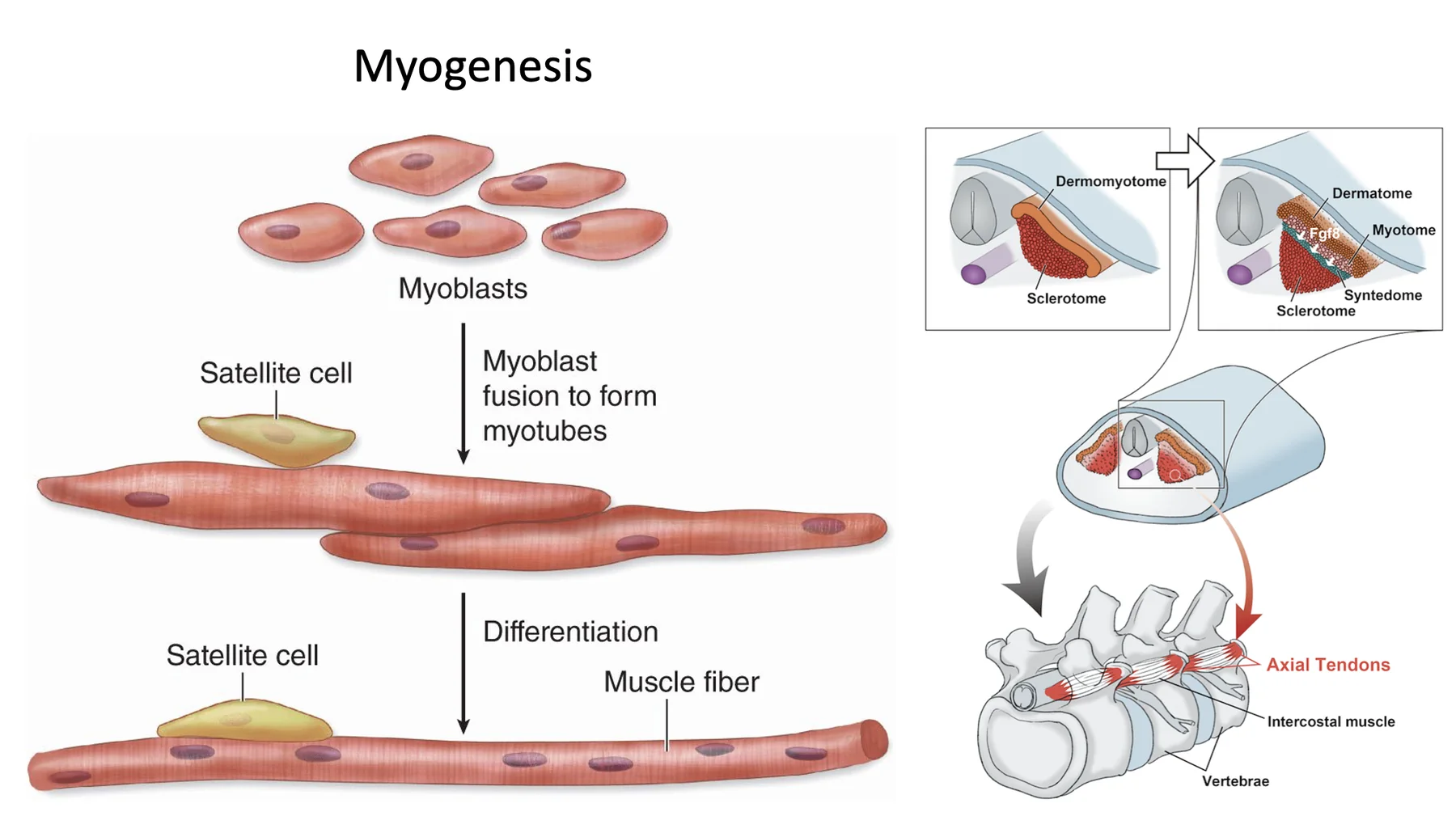
מיוגנזה (Myogenesis) היא תהליך יצירת שריר. המיובלסטים מתמיינים ומתאחים זה עם זה ליצירת סיבי שריר רב־גרעיניים (סיבי שריר בוגרים מכילים יותר מגרעין אחד).
במהלך ההתמיינות נוצרים גם תאי לוויין (Satellite cells): תאי גזע בוגרים שהתמיינו מהמיובלסטים, ונשארים בשריר הבוגר במצב רדום יחסית. במקרה של פגיעה בשריר הם יכולים להתעורר, להתחלק ולתרום להתחדשות סיבי שריר.
רה־סגמנטציה: למה סומיט אחד אינו חוליה אחת
סומיט אחד לא הופך ישירות לחוליה אחת. כל סומיט מתחלק לאזור רוסטרלי ולאזור קאודלי. בתהליך רה־סגמנטציה (Resegmentation), כל חוליה מורכבת מחלקים של שני סומיטים סמוכים:
- החלק הקאודלי (אחורי) של סומיט אחד (Caudal פירושו ״זנב״). זהו רוב החוליה.
- החלק הרוסטרלי (קדמי) של ה־סומיט הבא (Rostral פירושו ״ראש״)
זה מאפשר סידור שבו שרירים וגידים יכולים לחבר בין חוליות ולא רק להיצמד לחוליה אחת. התוצאה היא מערכת שיכולה לאפשר תנועה בין חוליות.
חשוב לדייק לגבי הגפיים: שרירי הגפיים מגיעים מתאים שמקורם בסומיטים ונודדים לניצני הגפיים. אך עצמות הגפיים אינן נוצרות מהסומיטים באותו אופן שבו החוליות נוצרות מהם.
שעון סגמנטציה ומודל Clock-and-Gradient
הסומיטים נוצרים בקצב מחזורי מדויק. כדי להסביר זאת הוצג מודל Clock-and-Wavefront, ובהמשך נוסח בשיעור כמודל מדויק יותר של Clock-and-Gradient.
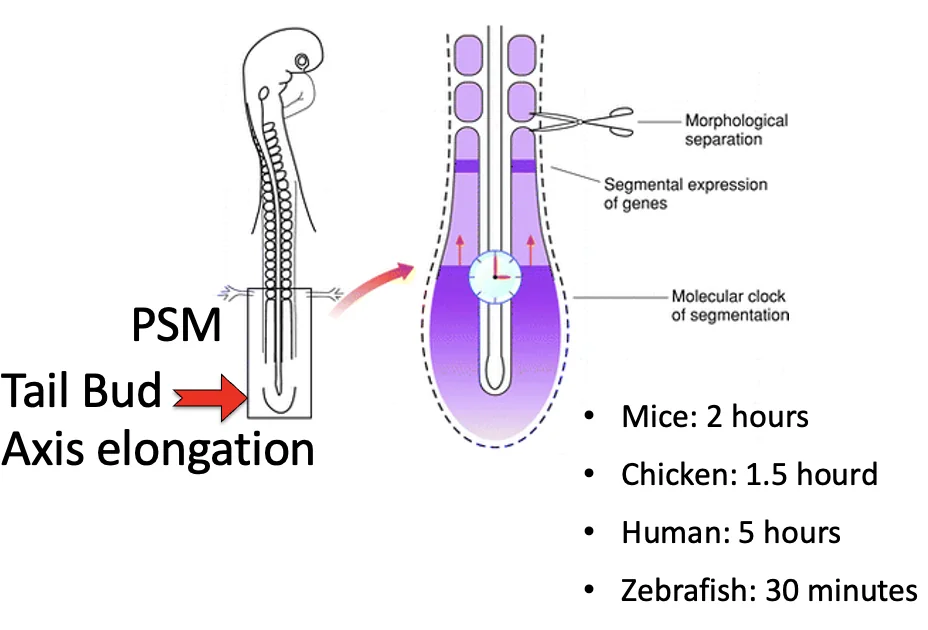
המודל כולל שני רכיבים:
| רכיב | משמעות |
|---|---|
| שעון | גן או קבוצת גנים שמתבטאים באופן מחזורי (לא כל הזמן) ב־PSM |
| Wavefront/Gradient | גבול הבשלה שנע אחורה (פוסטריורית) בזמן שהציר מתארך |
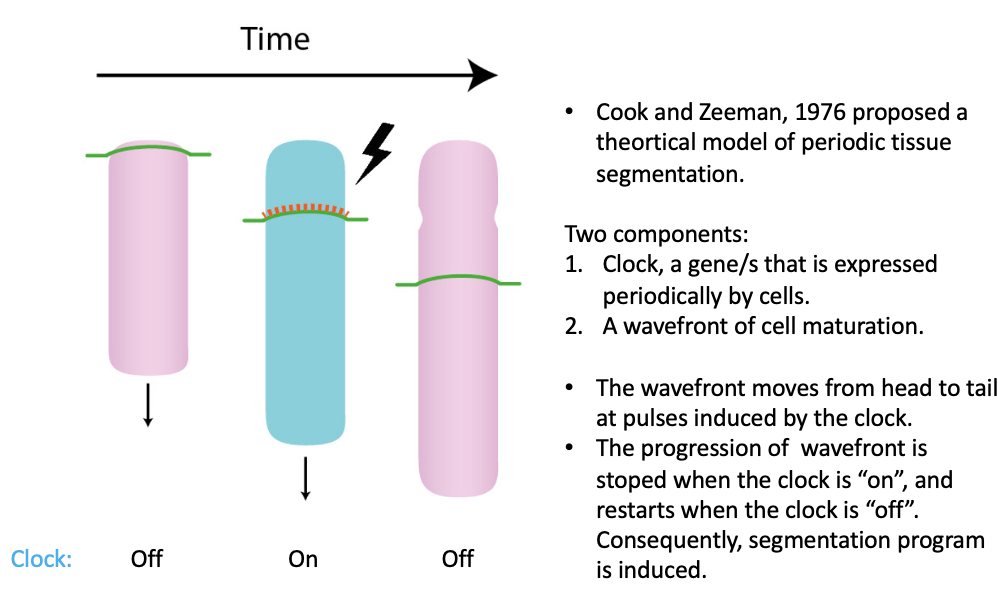
Hes7 - השעון
בעובר תרנגולת הוצג Hairy1, ובעכבר גן הומולוגי כמו Hes7. ב־in situ hybridization רואים דפוסי ביטוי שונים גם בעוברים עם אותו מספר סומיטים. לכן לא מדובר רק בעוברים שנמצאים בשלבי התפתחות שונים, אלא במחזור ביטוי בתוך אותו חלון זמן.
מספר הסומיטים משמש כמדד לשלב ההתפתחותי, כי קשה לתפוס בדיוק את נקודת הזמן בתוך מחזור קצר של שעה וחצי או שעתיים. אם עוברים עם אותו מספר סומיטים מציגים דפוסי ביטוי שונים, זה מתאים לרעיון של ביטוי מחזורי.
בצילום בזמן אמת, כאשר GFP מוכנס ללוקוס של גן מחזורי, אפשר לראות שהביטוי עולה ויורד בתאים, ושהדפוס מתקדם באופן דינמי לאורך ה־PSM (קרי, הביטוי מופיע באוזרים שונים, בשונה ממצב של ״מופיע״ או ״לא מופיע״). לכן השעון הוא למעשה אוסצילטור, הוא כולל רצף של מצבים והוא אינו בינארי.
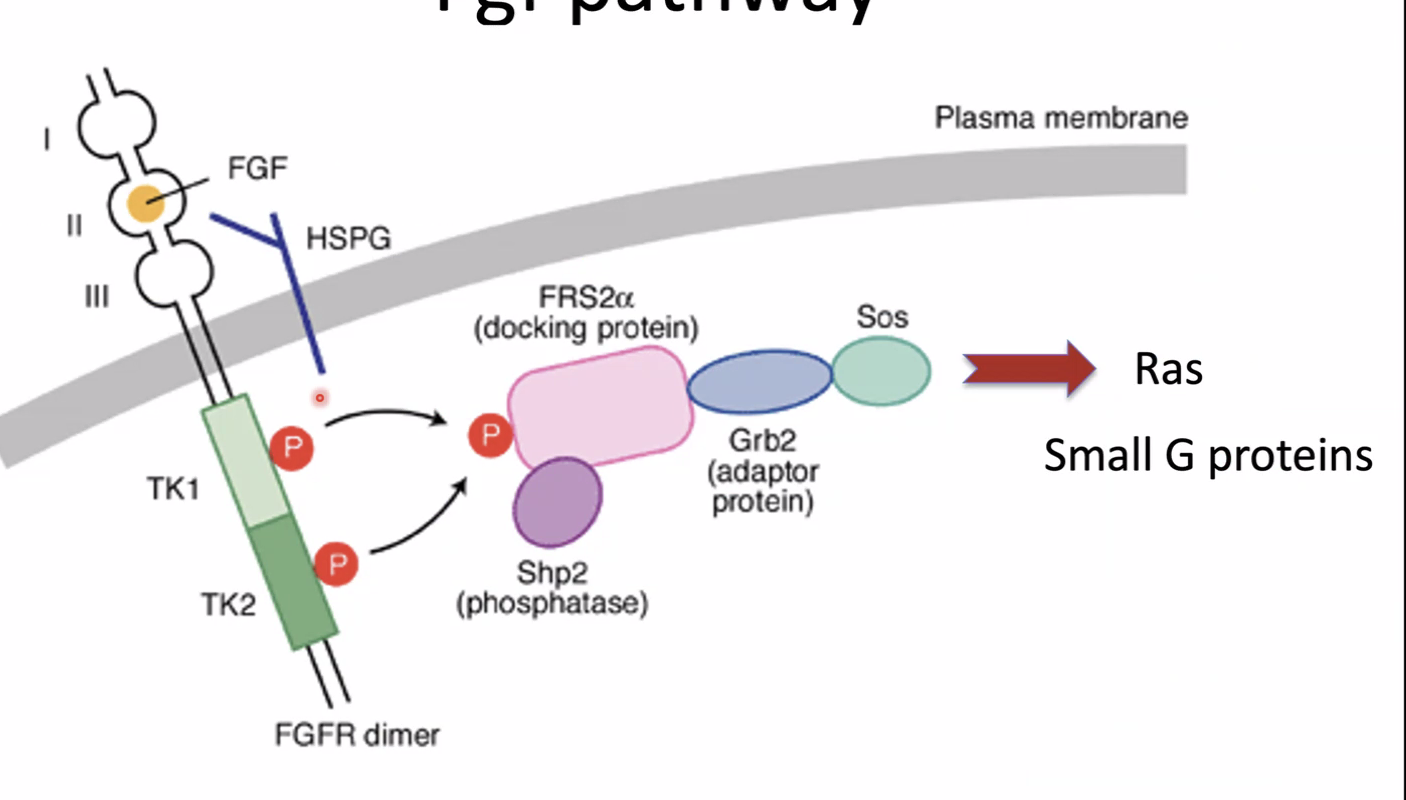
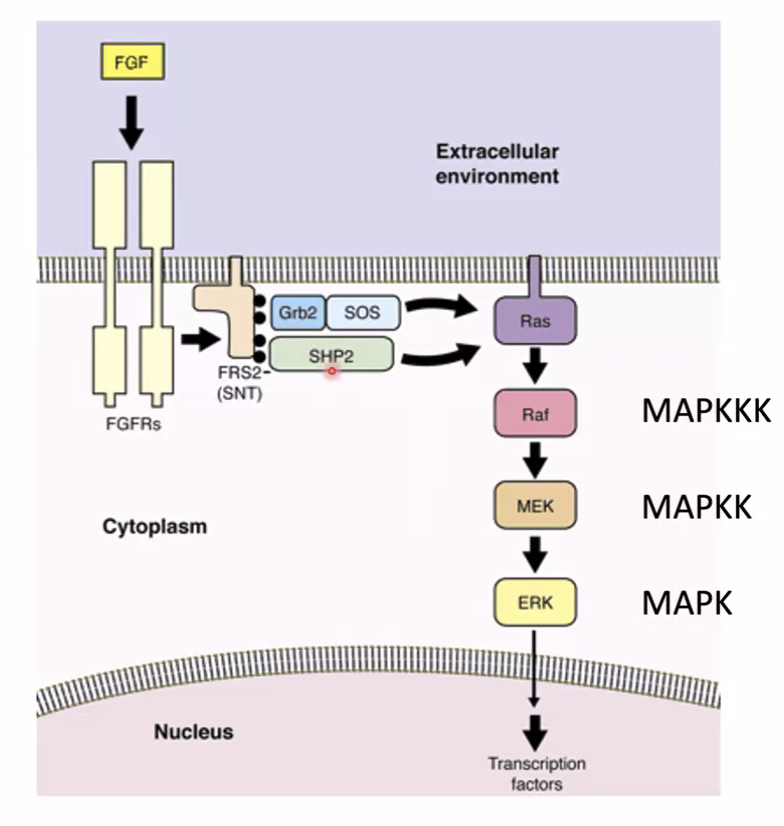
FGF8 - הגרדיאנט
המרכיב השני הוא ה־Wavefront. ה־FGF8 הוצג כאחד הגורמים החשובים ב־PSM.
העיקרון:
| מצב FGF signaling | מצב התאים |
|---|---|
| FGF גבוה | תאי PSM נשמרים במצב לא ממוין / immature |
| FGF נמוך יותר | התאים יכולים לעבור הבשלה (maturation) ולהיות מוכנים לסגמנטציה |
בצד הפוסטריורי, באזור ה־Tailbud, יש רמות גבוהות יותר של FGF signaling. בזמן שהציר מתארך, ה־front של הגרדיאנט נע לכיוון פוסטריורי, ותאים שנשארים מאחור כבר אינם חשופים לרמות FGF גבוהות. אז הם יכולים לעבור הבשלה ולהגיב לשעון הסגמנטלי.
איך נוצר גרדיאנט FGF8 ברמת RNA וחלבון
ב־in situ ל־FGF8 רואים גרדיאנט גם ברמת ה־RNA. זה מעניין, כי RNA בדרך כלל נוצר בתוך תאים מסוימים ולא ״מתפזר״ כמו חלבון מופרש.
כדי להבין זאת משתמשים בשני probes שונים:
| Probe | מה הוא מזהה | מה לומדים ממנו |
|---|---|---|
| Probe ל־mature mRNA | RNA בשל בציטופלזמה | איפה נמצא ה־mRNA שנוצר ונשמר בתאים |
| Probe לאינטרון | RNA בזמן שעתוק | איפה מתרחש transcription ברגע המדידה |
כאשר משתמשים ב־probe לאינטרון, רואים ששעתוק FGF8 מתרחש בעיקר ב־Tailbud. כאשר משתמשים ב־probe ל־mature mRNA, רואים mRNA גם באזור רחב יותר ב־PSM.
ההסבר הוא RNA decay mechanism:
- תאים באזור ה־Tailbud משעתקים FGF8.
- בגלל Axis elongation, ה־Tailbud ממשיך להתארך לכיוון פוסטריורי.
- התאים ששיעתקו FGF8 נשארים מאחור.
- באותם תאים, כבר אין שעתוק פעיל של FGF8 באותה רמה, אבל ה־mRNA שנוצר קודם לכן עדיין קיים.
- עם הזמן ה־RNA עובר degradation.
- כך נוצר גרדיאנט של FGF8 RNA, ובהמשך גם גרדיאנט חלבון.
הגרדיאנט הזה מספיק רחב כדי לשמש כ־Wavefront: הוא מגדיר אילו תאים עדיין צעירים ולא ממיינים, ואילו תאים כבר יכולים להיכנס לתוכנית סגמנטציה.
Clock-and-Gradient model
המודל הסופי מחבר בין שני רכיבים:
- FGF8 gradient שומר על תאי PSM במצב לא ממוין באזור הפוסטריורי, וכאשר הוא נסוג אחורה עם ה־axis elongation, תאים עוברים הבשלה.
- Oscillating genes כמו Hairy1/Hes7 יוצרים דפוס ביטוי מחזורי שמגיע לאזור שבו התאים כבר מוכנים להגיב.
כאשר הגל המחזורי מגיע לתאים שכבר יצאו מאזור FGF גבוה ועברו הבשלה, מופעלת תוכנית סגמנטציה, בין היתר דרך Mesp2, ונוצר סומיט חדש.
רצף האירועים המרכזי
- בסוף הגסטרולציה יש שלוש שכבות נבט, ובקו האמצע נוצר Axial mesoderm.
- ה־Axial mesoderm יוצר נוטוכורד.
- הנוטוכורד גורם לאקטודרם שמעליו להפוך ל־Neural Plate.
- ה־Neural Plate מתקפל דרך MHP ו־DLHP, עובר Fusion ויוצר Neural Tube.
- תאי Neural Crest יוצאים מהאזור הדורסלי של צינור העצבים, עוברים EMT ונודדים.
- צינור העצבים נסגר בנקודות Closure שונות, ובין נקודות הסגירה קיימים זמנית Neuropores.
- כשל בסגירה גורם ל־Neural Tube Defects, כמו Anencephaly או Spina bifida.
- ב־Spinal cord, גרדיאנטים של Shh, BMP ו־Wnt מייצרים דומיינים שונים של transcription factors.
- דומיינים אלה מובילים להתמיינות נוירונים שונים, למשל Motor neurons בצד הוונטרלי (שם יש Shh גבוה) ו־Sensory neurons בצד הדורסלי (שם יש BMP/Wnt גבוהים).
- במקביל, ה־Paraxial mesoderm עובר סומיטוגנזה ויוצר Somites בקצב קבוע.
- כל Somite מתמיין ל־סקלרוטום, Dermomyotome, מיוטום, דרמטום ו־Syndetome.
- הסקלרוטום יוצר חוליות וצלעות, ה־מיוטום יוצר שרירי שלד, הדרמטום יוצר דרמיס, וה־Syndetome יוצר גידים.
- ה־Segmentation clock וגרדיאנט FGF8 מתזמנים את יצירת ה־Somites החדשים לאורך הציר.