התוכן הוא לפי חומרי הלימוד:
Guyton and Hall, Physiology, 14th edition, chap 6-8.
מומזנים לקרוא גם את התרגול בנושא, שכולל נושאים נוספים שלא הוזכרו בשיעור.
שריר שלד
תפקידים
שריר שלד משמש ל:
- ייצור תנועה - הזזת גפיים ועצמות
- שמירת טונוס - גם במנוחה שרירים שומרים על מתח בסיסי שמחזיק אותנו בתנוחה
- רגולציה תרמית - ייצור חום, בעיקר דרך רעידות שריר בקור
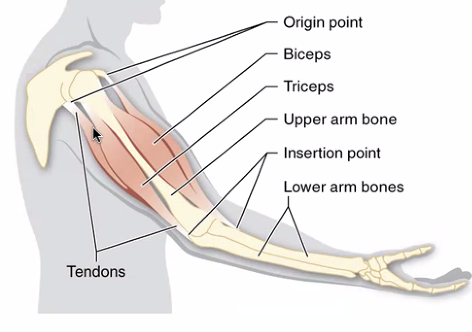
מכניקת תנועה
בדרך כלל, שריר שלד מחובר לשתי עצמות ועובר דרכן:
- Origin (מוצא) - נקודת חיבור לקצה עצם אחת
- Insertion (נקודת אחיזה/היאחזות) - נקודת חיבור ליד המפרק של העצם השנייה
כשהשריר מתכווץ, ה־Insertion נמשך לכיוון ה־Origin ונוצרת תנועת קיפול במפרק. לדוגמה: כיווץ ה־Biceps מרים את היד, בעוד שה־Triceps נמתח פסיבית. בתרגיל יד אחורית - ההפך.
סוגי שרירים ומבנים
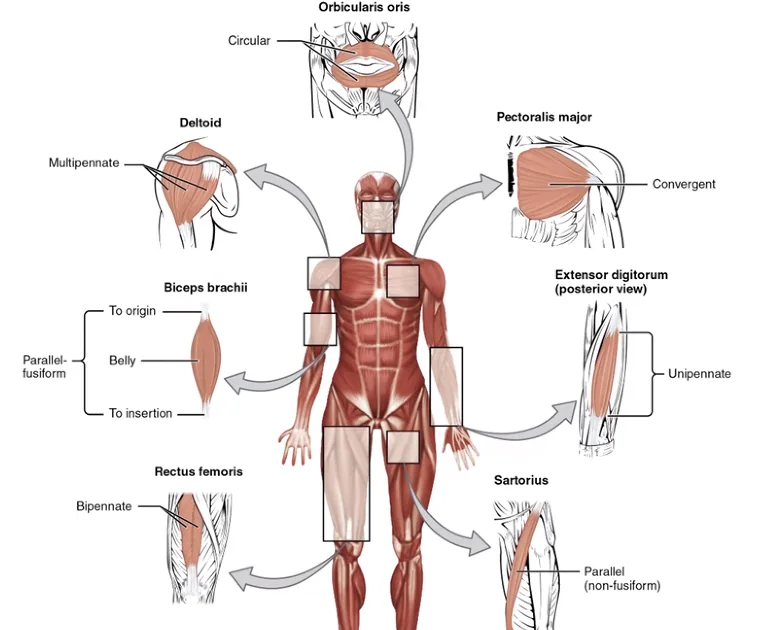
בהתאם לתפקיד השריר, ישנם מבנים חיצוניים שונים:
- Fusiform (ציר) - רחב באמצע ומחודד בקצוות, כמו ה־Biceps; מבנה סימטרי
- Multipennate - כמו ה־Deltoid בכתף; רחב, ארוך, ומכיל קבוצות שריר שונות שכל פעם קבוצה אחרת פעילה
- Circular (סרקולרי) - סביב פתחים כמו עין ושפתיים
- Parallel - אזור הירך הקדמי; עובי דומה ב־Insertion וב־Origin
למרות ההבדלים החיצוניים, המבנה הפנימי של כל השרירים זהה.
שריר לבן ושריר אדום (לידיעה כללית, מחוץ לסקופ הבחינה)
- שריר לבן (גליקוליטי/אנאירובי) - מהיר, מתפרץ, תלוי גליקוליזה; אופייני לשרירי חזה עוף
- שריר אדום (אירובי) - סיבולת, צריכת חמצן, עשיר במיוגלובין; אופייני לשרירי הרגליים
היחס בין סוגי הסיבים ניתן לשינוי על ידי אימון ומשפיע על ביצועי השריר.
הערה: למרות שבשיעור המידע הוצג בהקשר של הרחבה, בתרגול דווקא הופיעו שאלות על סוגי שרירים. לתשומת לבכם.
בתרגול גם הודגש ששריר אדום מתאפיין במיתוכונדריות, וביותר זרימת דם וחמצן.
המבנה ההיררכי של שריר שלד
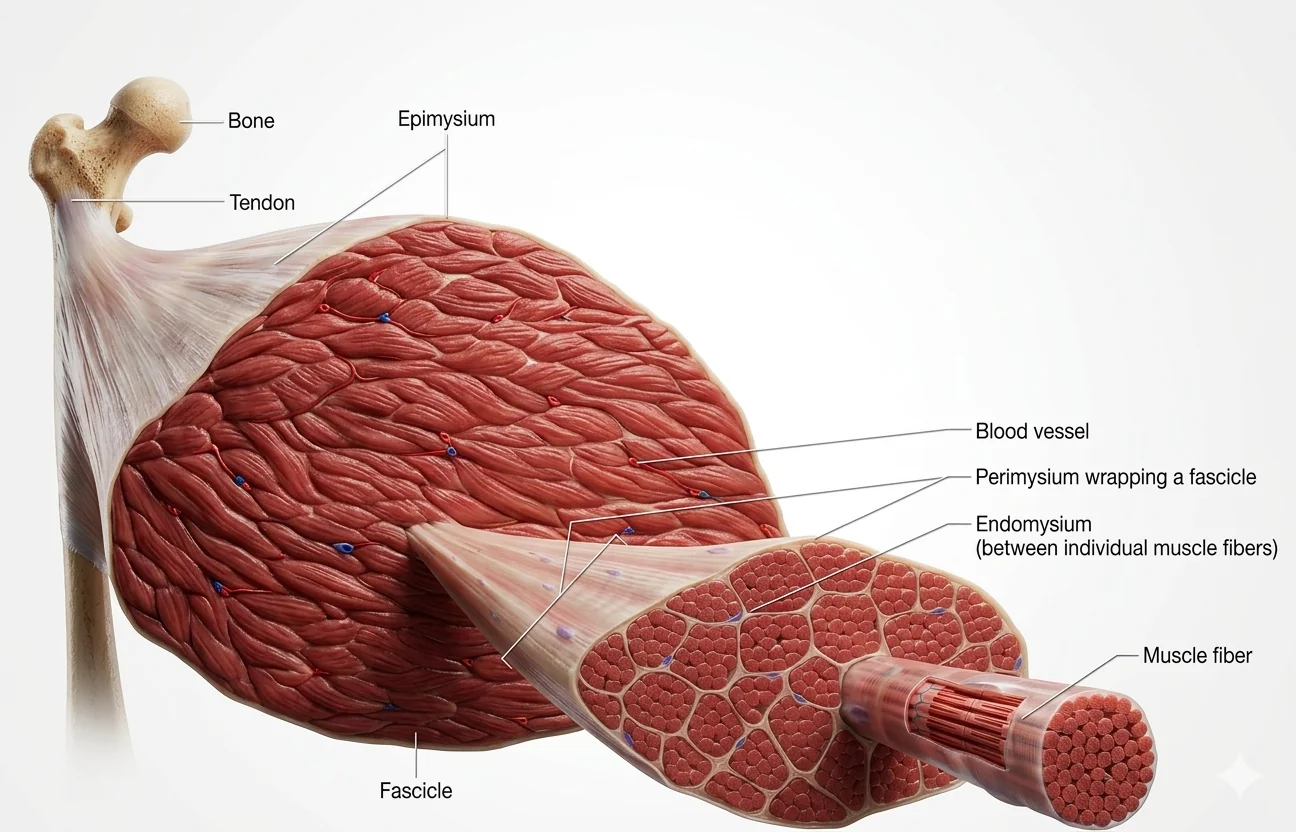
- שריר שלם
- Fascicles (חבילות מוקפות בפאסיה)
- Muscle Fibers/Myocytes (תאי שריר)
- Myofibrils (סיבונים תוך־תאיים)
Muscle Fiber - תא שריר בודד
- Multinucleated - מכיל מספר גרעינים בפריפריה של התא (נוצר מ־fusion של מיובלסטים בהתפתחות)
- קוטר: 80-10 מיקרון
- מכיל Myofibrils רבים הממלאים את נפחו
- מוקף בSarcolemma - הממברנה הייחודית של תא שריר
Myofibril - יחידה תת־תאית
- חוט דק וארוך המורכב מ
אקטיןוממיוזין - ניתן לבודד אותו (בצורה כימית) ולצפות בו כחוט בודד
- כולם שוכנים בתוך Sarcoplasm (הציטוזול של תא שריר) משותף, עטופים בממברנה אחת
בהתפתחות, מיובלסטים בודדים עוברים Fusion ויוצרים את תא השריר הרב־גרעיני. תאי לוויין (Satellite cells) בתוך נישות של כל שריר משמשים כמאגר לצמיחה ולהתחדשות.
הסרקומר - היחידה הפונקציונלית
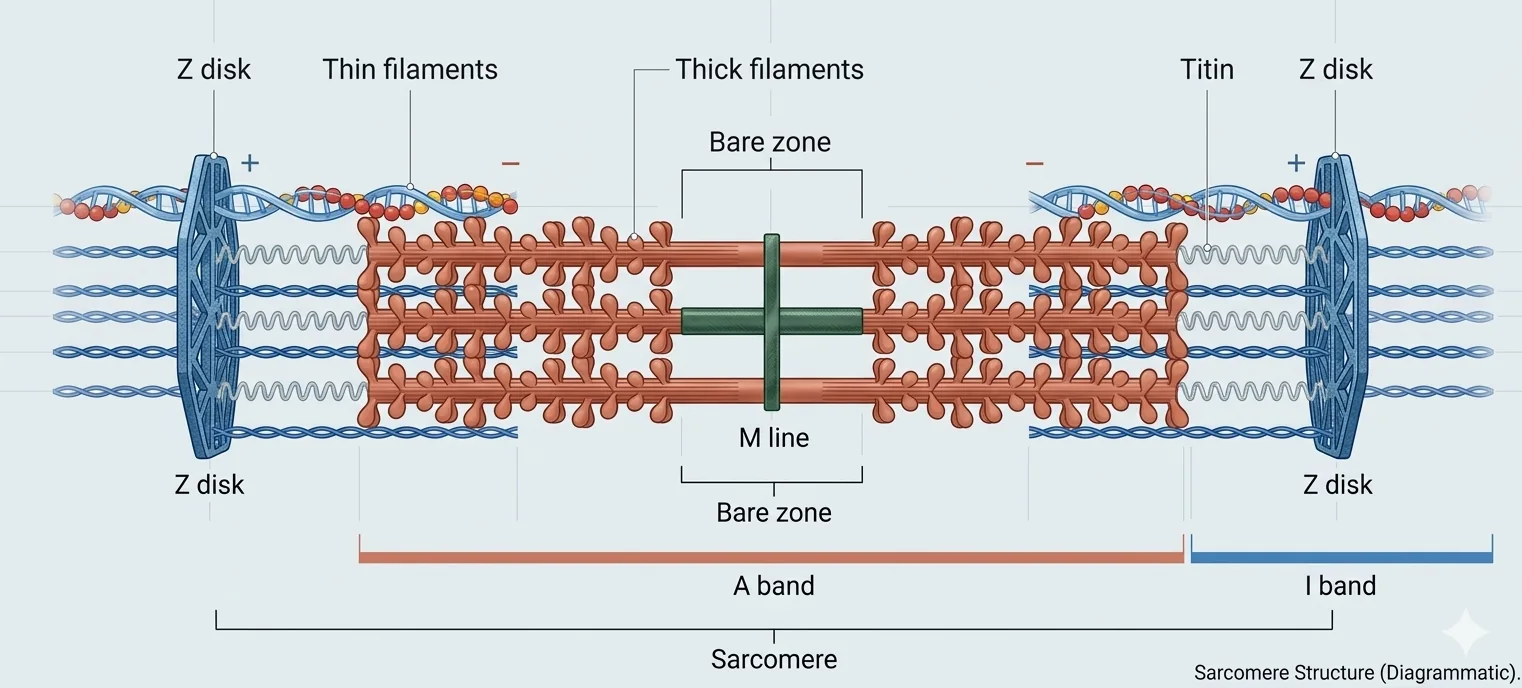 | 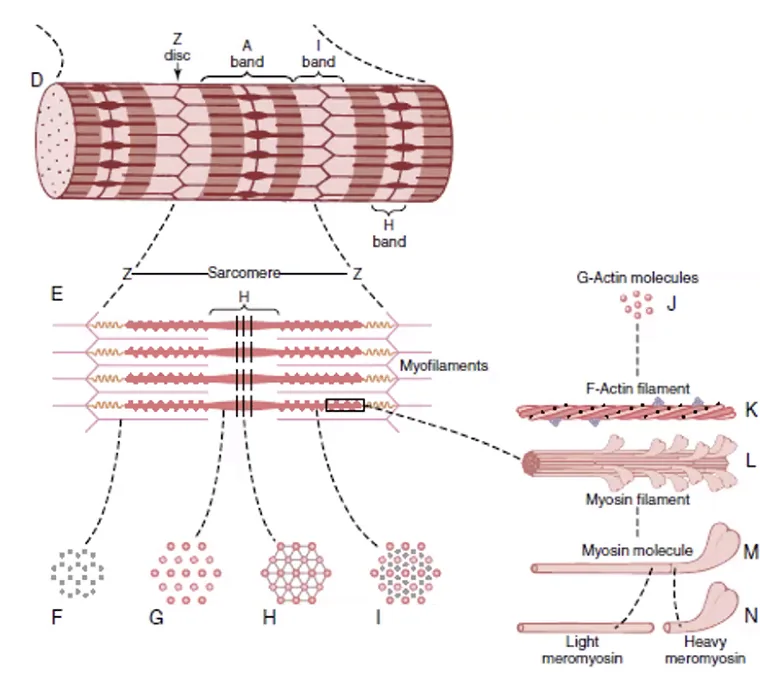 |
כל Myofibril מורכב מרצף ארוך של סרקומרים (Sarcomeres) שמסודרים בטור. כל סרקומר הוא יחידה פונקציונלית אחת - המרחק בין שני Z-Disks עוקבים.
מרכיבי הסרקומר
| מרכיב | תיאור |
|---|---|
Z-Disk | “קיר” שחוצץ בין סרקומרים, מעגן את חוטי האקטין. עשוי מחלבון α-Actinin |
A-Band | אזור כהה - לכל אורך פילמנט המיוזין (כולל אזור החפיפה עם אקטין) |
I-Band | אזור בהיר - אזור אקטין בלבד (ללא מיוזין), נפרס על פני שני סרקומרים שכנים; ה־Z-Disk עובר במרכזו |
M-Line | קו האמצע של פילמנט המיוזין; חלבונים מיוחדים מעגנים אותו ושומרים על סימטריה |
H-Zone (Bare Zone) | אזור באמצע ה־A-Band שיש בו מיוזין ללא ראשי מיוזין |
חשוב: בהתכווצות, ה־
I-Bandמתקצר (ה־Z-Diskמתקרבים), ה־A-Bandנשאר קבוע (אורך המיוזיןלא משתנה).
ה־Z-Disk ומחלות שריר
מוטציות בחלבונים של ה־Z-Disk פוגעות בעיגון המכני של האקטין לקירות ובצימוד בין סרקומרים ← Myopathies (מחלות שריר הגורמות לחולשה ולפגיעה ביכולת בניית שריר).
טיטין (Titin)
חלבון ענק (אחד הגדולים בגוף, כ־4,000-3,000 ק”ד) המחבר את פילמנט המיוזין ל־Z-Disk. מצוייר כקפיץ - ולא בכדי:
- תפקיד מכני פסיבי: מקנה לשריר את האלסטיות הפסיבית שלו - קובע את הקשיחות הפסיבית הבסיסית
- הגנה על
מיוזין: מחזיק את פילמנטי המיוזיןבמרכז הסרקומר ומונע מהם להיסחף - תפקידים רגולטוריים: אתרים על טיטין מאפשרים קישור חלבונים נוספים המווסתים תכונות שריר
טיטין הוא פסיבי - לא משתתף אקטיבית בהתכווצות, אך קובע את תכונות השריר בהיעדר אקטיבציה.
T-Tubules והרשת הסרקופלסמית (Sarcoplasmic Reticulum - SR)
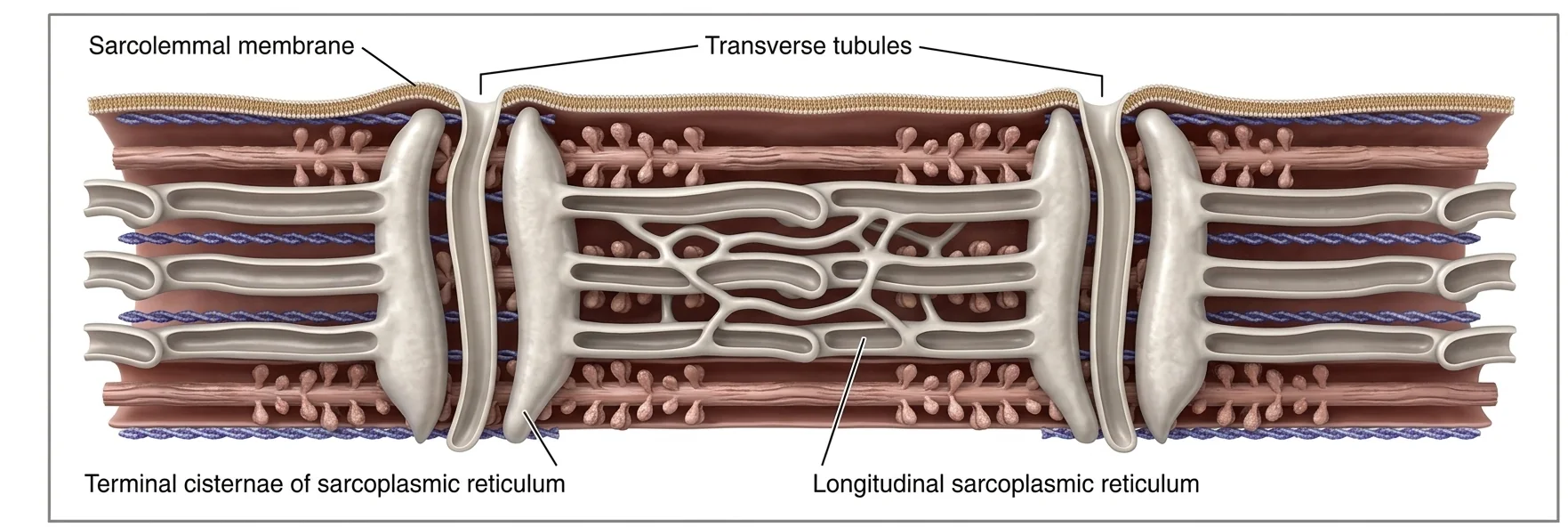
Sarcolemma
הממברנה הייחודית של תא שריר. יש עליה:
- פוטנציאל ממברנה ותעלות יונים
- T-Tubules - שקעים שנוצרים מהממברנה ונכנסים לעומק התא
T-Tubules - מדוע הם נחוצים?
תא שריר יכול להגיע לקוטר של כ־80 מיקרון ומכיל מיופיברילים רבים. אם העירור החשמלי היה מתפשט רק על פני הממברנה החיצונית, היה לוקח לו זמן להגיע לעומק התא. ה־T-Tubules מוליכים את גל הדה־פולריזציה לעומק התא, וכך מאפשרים הפעלה מהירה, אחידה ומתואמת של כל המיופיברילים.
Sarcoplasmic Reticulum (SR)
רשת של אורגנל תוך־תאי עם ממברנה משלו - מאגר סידן ($\ce{Ca^2+}$) תוך־תאי.
מאפיינים:
- מכיל בתוכו ריכוז גבוה מאוד של $\ce{Ca^2+}$
- משאבות SERCA (ATP-dependent) על הממברנה - מחזירות $\ce{Ca^2+}$ פנימה אחרי האקטיבציה
- נפרס לאורך כל הסרקומרים בסמוך ל־T-Tubules
ה־T-Tubule ממוקם בין שני ה־SR הסמוכים, ומשפעל אותם בצורה מקומית ← $\ce{Ca^2+}$ משתחרר לציטוזול ומפעיל את הפילמנטים.
חיבור מכני לגיד
Glycocalyx (ג’ל עשיר ב־ECM וקולגן) על פני הממברנה יוצר סיבי חיבור שמעגנים את סיבי השריר ומחברים אותם לגיד ← הגיד מתחבר לעצם. קולגן הוא קריטי לבריאות הגידים.
פילמנט המיוזין (Thick Filament)
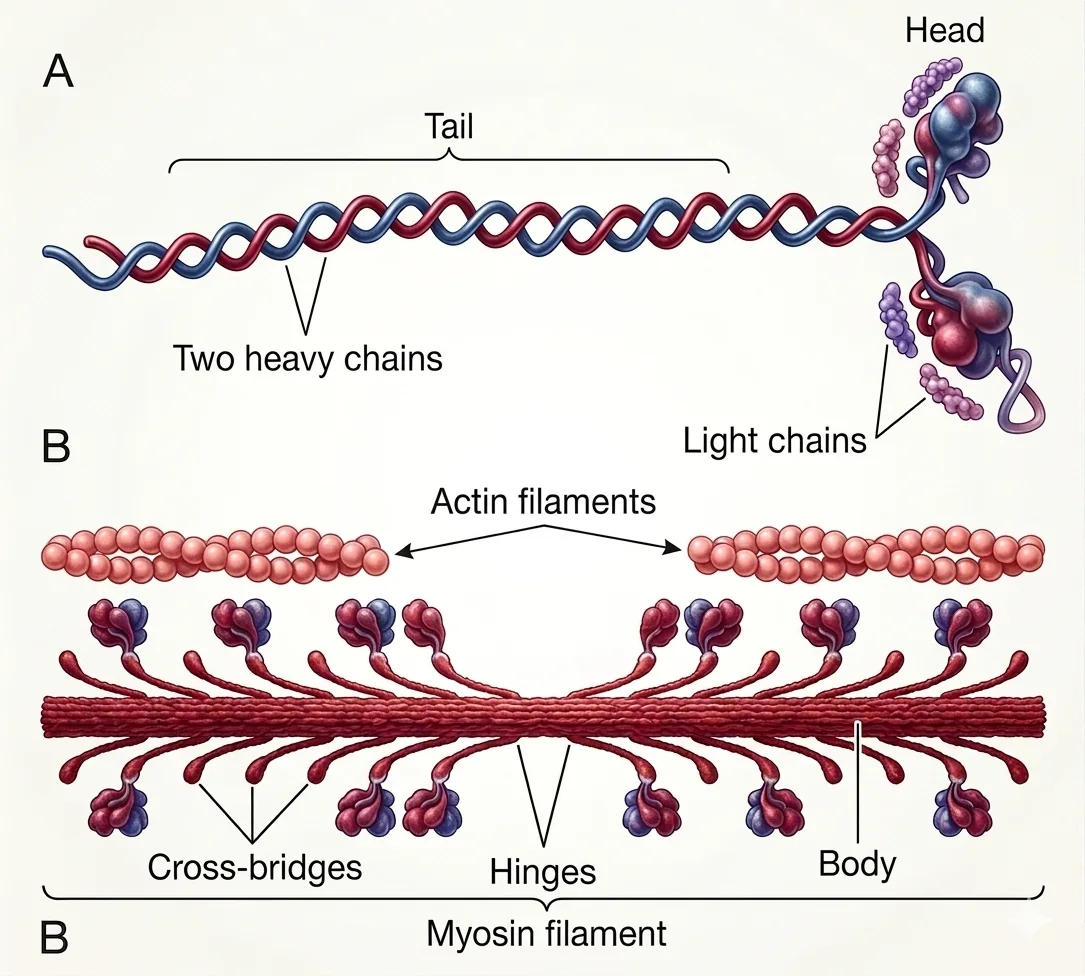
כל פילמנט מיוזין מורכב מכ־200 מולקולות מיוזין.
מבנה מולקולת מיוזין - שישה פפטידים
2 Heavy Chains- יוצרות את ה”זנב” הארוך (מלופפות בהליקס); הזנב מרכיב את גוף הפילמנט4 Light Chains- שתיים לכל ראש; תפקיד רגולטורי בבקרת מחזור ה־Cross-Bridge(להלן)
Cross-Bridge
הכינוי לצמד צוואר + ראש גלובולרי שיוצא מהפילמנט לכיוון האקטין. מאפיינים:
- ראש גלובולרי - אתר קטליטי להידרוליזה של ATP
- תכונות קפיץ - ה־Cross-Bridge יכול להיטען באנרגיה כקפיץ נמתח ולשחרר אותה ב־Power Stroke
- שני צירי תנועה (Hinges): אחד בחיבור הזרוע לפילמנט, ושני בחיבור הצוואר לראש
אורך פילמנט המיוזין: 1.6 מיקרון (קבוע!)
מגביל את אורך הסרקומר המינימאלי: כשה־Z-Disk מגיעים ל־1.6 מיקרון, הסרקומר מתקפל לחלוטין (אין קיצור פיזיולוגי נוסף מתחת לאורך הזה).
פילמנט האקטין (Thin Filament)
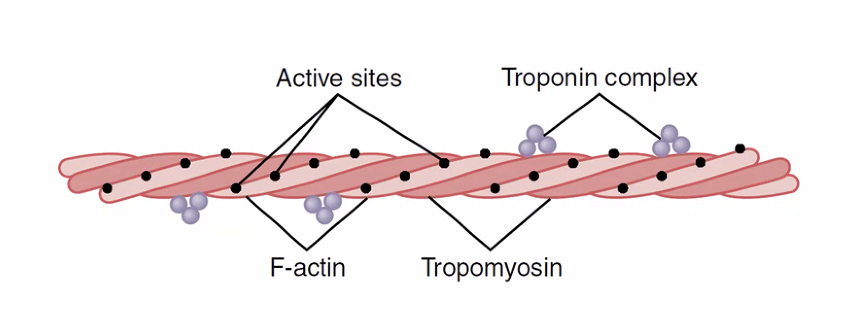
מבנה
- שתי שרשראות
אקטיןמלופפות בהליקס כפול - בשקע שבין השרשראות - Active Sites (אתרי קישור ל
מיוזין), מסודרים במרחקים שווים - אורך: כ־1 מיקרון מכל צד של ה־
Z-Disk - מחובר ל־
Z-Diskבצורה קשיחה (בניגוד למיוזיןשמחובר דרך הטיטין הגמיש)
Tropomyosin - מולקולת חסימה
מולקולה ארוכה שנכרכת לאורך האקטין. תפקיד במנוחה: חוסמת את ה־Active Sites - מונעת מהמיוזין להיקשר לאקטין גם אם ירצה.
קומפלקס הטרופונין - מנגנון הבקרה
הטרופונין יושב על ה־Tropomyosin במרחקים שווים לאורך הפילמנט. מורכב משלוש יחידות משנה:
| יחידה | תפקיד |
|---|---|
| Troponin T (TnT) | קשור ל־Tropomyosin |
| Troponin I (TnI) | קשור ישירות לאקטין; מחבר את הקומפלקס לפילמנט |
Troponin C (TnC) | קושר $\ce{Ca^2+}$ - קריטי להפעלת ההתכווצות |
מנגנון הפעולה:
- עליית $\ce{Ca^2+}$ ציטוזולי ← נקשר ל־TnC (אפיניות גבוהה)
- שינוי קונפורמציה בקומפלקס ← Tropomyosin נמשך ומסתובב
- ה־Active Sites נחשפים ←
מיוזיןיכול להיקשר לאקטין
משוב חיובי: ברגע שסידן מתחיל להיקשר ל־TnC, האפיניות של TnC לסידן עולה עוד יותר - גורם לפרופגציה מהירה מאוד של הקישור לכל אורך הפילמנט. המנגנון הזה, יחד עם ה־SR שנפרס לאורך הסרקומרים, מאפשר הפעלה אחידה ומהירה של כל תא השריר.
מנגנון ההתכווצות - Sliding Filament Theory
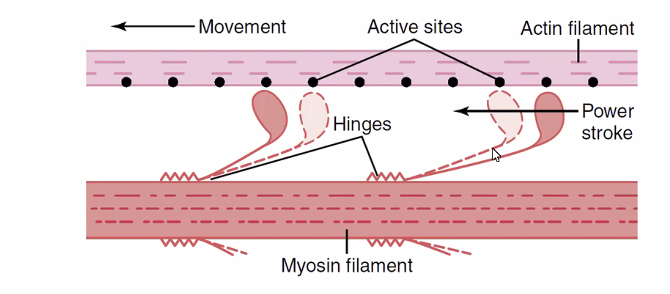
בהתכווצות, פילמנטי האקטין “מחליקים” על פני פילמנטי המיוזין, ה־Z-Disks מתקרבים והסרקומר מתקצר.
- הפילמנטים עצמם לא מתקצרים - רק החפיפה ביניהם גדלה
- ה־
A-Band(אורךמיוזין) - קבוע - ה־
I-Bandוה־H-Zone- מתקצרים
מחזור ה־Cross-Bridge (Cross-Bridge Cycle)
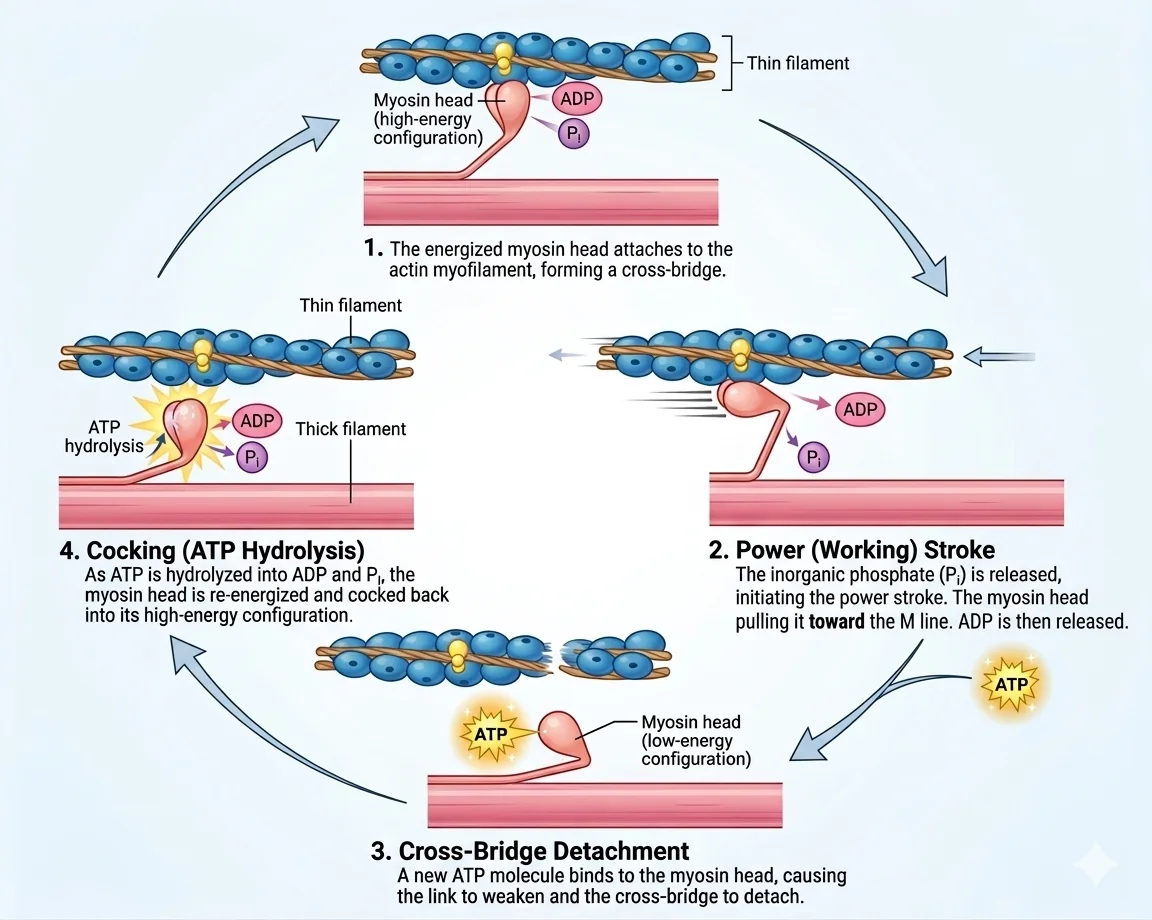
תנאי הכרחי להתחלה: סידן $\ce{Ca^2+}$ קשור ל־TnC (ה־Active Sites חשופים)
שלבי המחזור (לא בסדר שהם מוצגים בתרשים):
- קישור ATP לראש ה
מיוזין: שינוי קונפורמציה, ניתוק הראש מהאקטין - הידרוליזה של
ATPאלADP + Pi(עדיין מצומדים לראש): “טעינת הקפיץ” - הראש נמשך למצב “מוכן” (Cocked position) - קישור ראש ה
מיוזיןל־Active Site עלאקטין(מקישור חלש לחזק) - שחרור
Pi: שחרור פופסט לא אורגני מוביל ל־Power Stroke - הקפיץ משתחרר, הראש מבצע תנועת “חתירה” ומזיז את האקטין; שחרור ADP - קישור ATP חדש: ניתוק הראש וחזרה לשלב 1
הפעולה חוזרת כל עוד יש $\ce{Ca^2+}$ על הפילמנטים ו־ATP זמין. כאשר $\ce{Ca^2+}$ מוחזר ל־SR,
Tropomyosinחוזר לחסום ומכאן ניתוק ורלקסציה.
שתי מערכות תהליכים דורשות ATP בשריר:
- מחזור ה־Cross-Bridge (כולל התנועה/ה־Power Stroke של ראש ה
מיוזין) - החזרת $\ce{Ca^2+}$ ל־SR באמצעות משאבות SERCA
בהיעדר ATP (מוות) - ראשי ה
מיוזין“נתקעים” על האקטין← Rigor Mortis (קשיון המוות).
Force-Length Relationship
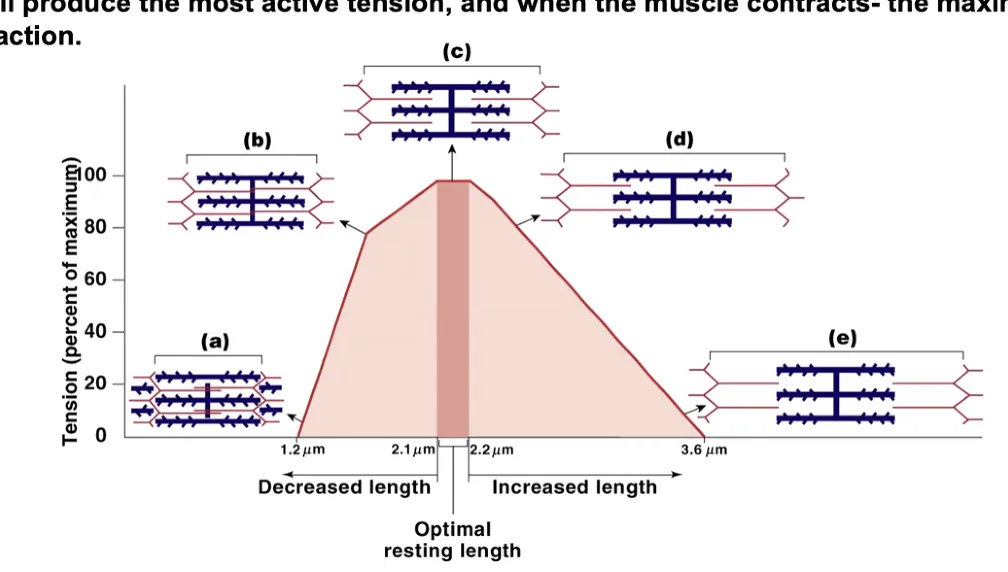
הכוח שסרקומר מסוגל לייצר תלוי באורך ההתחלתי שלו (מידת החפיפה בין אקטין למיוזין):
| אורך סרקומר | מצב | כוח |
|---|---|---|
| $<1.6 \, \mathrm{μm}$ | Buckling - הפילמנטים קורסים; אקטין מפריע לעצמו | אפס |
| $\sim 2.0-2.2 \, \mathrm{μm}$ | חפיפה מקסימלית אפקטיבית בין אקטין למיוזין | מקסימלי |
| $\sim 2.0-2.4 \, \mathrm{μm}$ | “פלטו” - חלק ה־H-Zone אינו תורם, אך אין הפרעה | גבוה, יציב |
| $>2.4-3.6 \, \mathrm{μm}$ | החפיפה פוחתת - פחות קרוס-בריג’ים | יורד |
| $>3.6 \, \mathrm{μm}$ | אין חפיפה | אפס |
אורך הסרקומר במנוחה (פיזיולוגי): $\sim 1.8-1.9 \, \mathrm{μm}$ - תחום עבודה נמצא לאורך הפלטו, קרוב לאופטימום.
מצבי הקצה (קצר מאוד / ארוך מאוד) אינם פיזיולוגיים - הם מצבים ניסיוניים בלבד.
Force-Length Relationship של שריר שלם
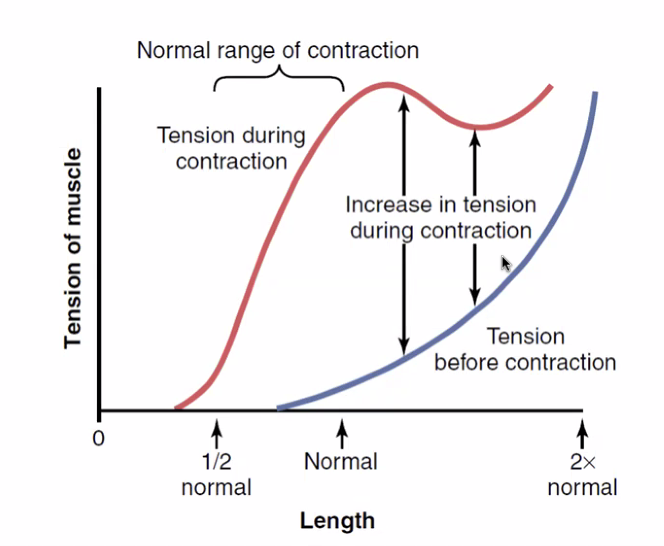
ברמת השריר השלם נוספת עקומת הפסיביות:
- עקומה כחולה (פסיבית) - מתח שריר ללא אקטיבציה חשמלית (תכונת רקמת החיבור, הגידים, הממברנה); עולה עם המתיחה
- עקומה אדומה (אקטיבית) - סכום התכונות האקטיביות + הפסיביות
- גם נצפה בשריר הלב - ככל שנפח מילוי גדל, כוח הפליטה עולה (חוק פרנק-סטארלינג)
דוגמה: קפיצה במקום - “טעינה אחורית” של השריר לפני הניתור מגדילה את האורך ההתחלתי ומאפשרת כוח מתפרץ גדול יותר.
Force-Velocity Relationship
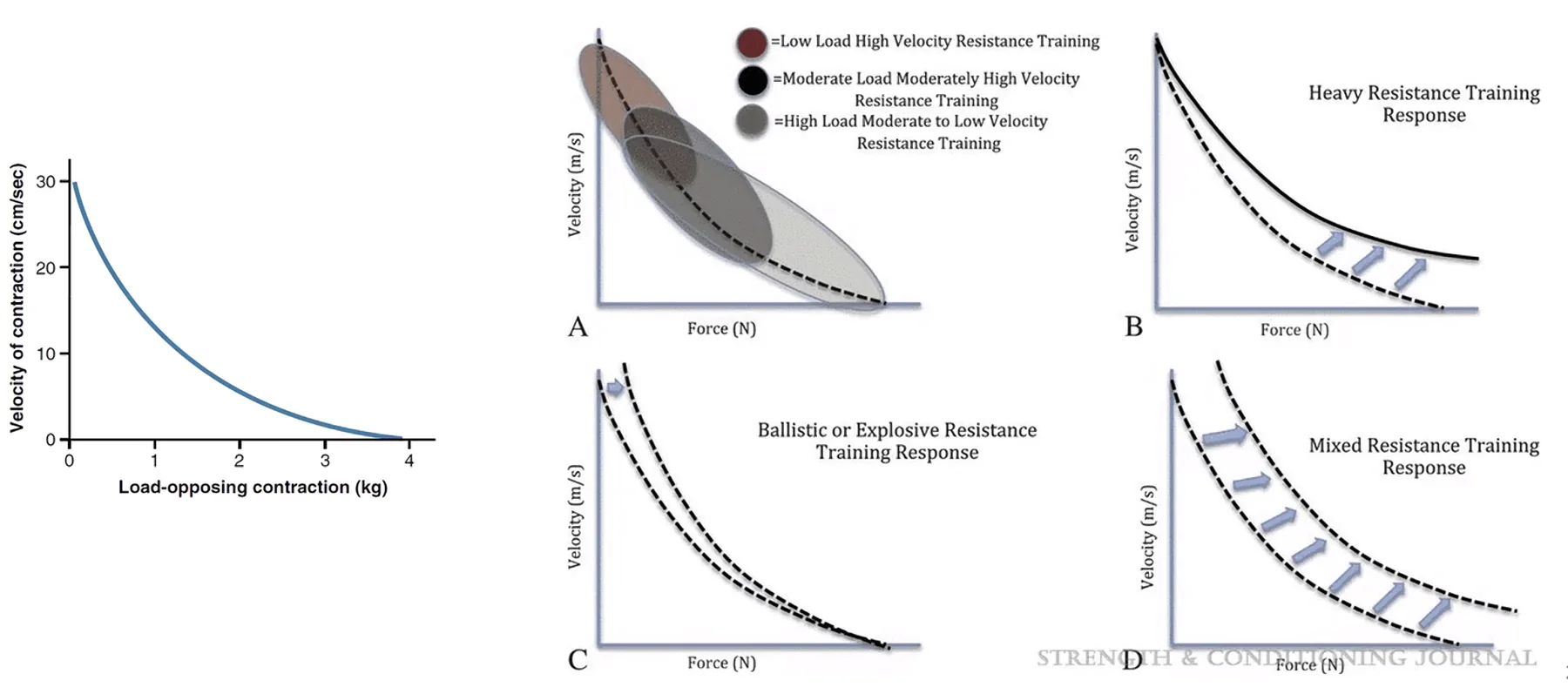
מהירות ההתכווצות של שריר קשורה לעומס שהוא נושא:
- עומס נמוך / אפסי (Unloaded) ← מהירות מקסימלית
- עומס גבוה ← מהירות יורדת
- עומס מקסימלי ← מהירות = אפס (Isometric)
אימון משנה את העקומה:
- אימון כוח (Resistance) - מושך את העקומה ימינה ← שריר מסוגל לשאת משקל גבוה יותר
- אימון מהירות/כוח מתפרץ - מעלה את ה־Unloaded Velocity (חיתוך עם ציר Y)
- אימון משולב - העקומה כולה עולה
השינויים האלה מתאפשרים בשל שינוי ביחסי האיזופורמים של
מיוזין- שינויים קלים ב־Heavy/Light Chains משנים את קינטיקת הידרוליזת ATP ואת תכונות השריר.
סוגי התכווצות
| סוג | הגדרה | דוגמה |
|---|---|---|
| Isometric | כיווץ בלי שינוי אורך - יצירת כוח ללא תנועה | להחזיק צלחת ללא הזזה |
| Isotonic Concentric | התקצרות פעילה כנגד עומס | הרמת משקולת למעלה |
| Isotonic Eccentric | התארכות פעילה - השריר מפעיל כוח אך נמתח | ירידה במדרגות (Quadriceps); הורדת משקולת לאט |
פעילות Eccentric נחשבת מאתגרת יותר מבחינה שרירית ואחראית ל־DOMS (כאב שרירים מושהה).
אקטיבציה של שריר - Excitation-Contraction Coupling
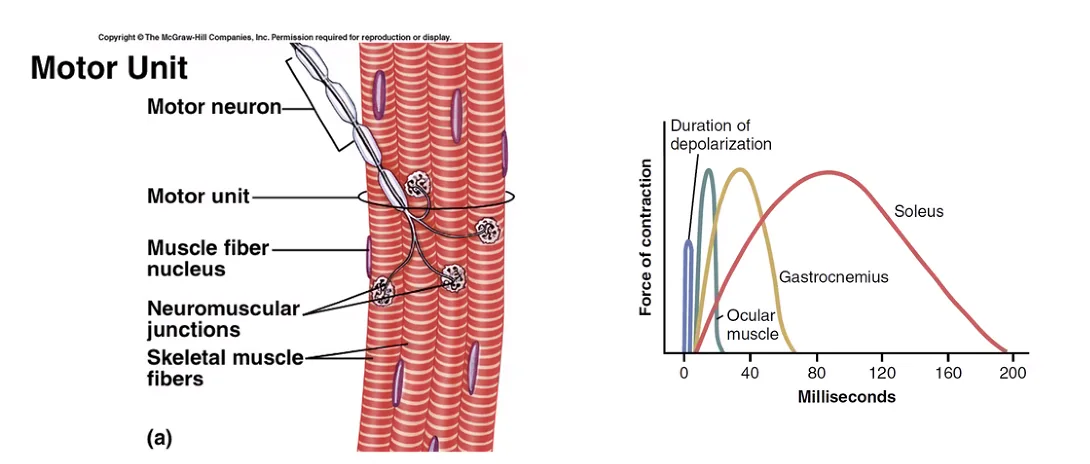
יחידה מוטורית (Motor Unit)
יחידה פונקציונלית המורכבת מ:
- נוירון מוטורי אחד (גוף תא + אקסון)
- קבוצת תאי שריר שהוא מעצבב
כל תאי השריר ביחידה מוטורית אחת הם מאותו סוג סיב (לבן / אדום).
שרירים שדורשים בקרה עדינה (כמו שרירי העין) - יחידות מוטוריות קטנות (1:1 עד 1:3). שרירים גסים (כמו Gastrocnemius) - יחידות מוטוריות גדולות.
Neuromuscular Junction (NMJ) - לוחית מוטורית
מקום הפגישה בין קצה הנוירון לממברנת השריר:
- Synaptic Cleft - מרווח של 30-20 ננומטר בין הטרמינל לממברנה; קריטי להעברת הסיגנל בצורה מהירה ומדויקת
- הטרמינל אינו מכוסה במיאלין (המיאלין נגמר לפני ה־NMJ)
- הטרמינל עשיר במיטוכונדריות, ובו גם מתבצעים סינתזה ואריזה של ACh בשלפוחיות סינפטיות.
- Synaptic Vesicles - שלפוחיות ממברנליות מכילות Acetylcholine (ACh) מוכן לשחרור
כיצד עובד ה־NMJ
- פוטנציאל פעולה מתקדם באקסון (מצופה במיאלין) עד לטרמינל
- בטרמינל - Voltage-Gated $\ce{Ca^2+}$ Channels נפתחות בתגובה לדה־פולריזציה
- כניסת $\ce{Ca^2+}$ לטרמינל ← הוסיקולות נודדות לממברנה ועושות Fusion ← שחרור ACh לחריץ הסינפטי
- ACh נקשר לNicotinic ACh Receptors על הממברנה של השריר - אלו הן תעלות יונים (Ligand-Gated) שנפתחות בקשירת ACh
- כניסת Na⁺ ← End-Plate Potential (EPP) - דה־פולריזציה לוקלית
- ה־EPP מפעיל Voltage-Gated Na⁺ Channels ← פוטנציאל פעולה המתפשט בכל הסרקולמה כולל T-Tubules
- Acetylcholinesterase בחריץ הסינפטי מפרק את ה־ACh מיד - כדי לצמד “ירי אחד - טוויץ’ אחד” ולמנוע אקטיבציה נוספת ללא פקודה
All-or-Nothing: קיים סף לאקטיבציה - מספיק ACh / דה־פולריזציה מספקת חייבת להצטבר כדי להפעיל פוטנציאל פעולה.
E-C Coupling - מה קורה בתוך תא השריר
תהליך מקיף המחבר בין הגירוי החשמלי להתכווצות המכנית:
- פוטנציאל פעולה ← מתקדם על הסרקולמה ← נכנס לT-Tubules
- שינוי מתח לוקלי ב־T-Tubule ← מפעיל DHP Receptors (Dihydropyridine Receptors) - רצפטורים תלויי-מתח
- DHP מתקשר פיזית לRyanodine Receptors (RyR) על ממברנת ה־SR ← נפתחים
- שחרור מהיר של $\ce{Ca^2+}$ מה־SR (מרחק ריכוזים עצום: ה־SR מרוכז בערך פי 10,000 מהציטוזול)
- $\ce{Ca^2+}$ נקשר לTroponin C ← הפעלת הפילמנטים ← Cross-Bridge Cycling ← התכווצות
- לאחר מילי-שניות בודדות - SERCA pumps (ATP-dependent) מחזירות $\ce{Ca^2+}$ ל־SR
- $\ce{Ca^2+}$ יורד ← TnC מאבד סידן ← Tropomyosin חוסם שוב ← רלקסציה
ריכוז $\ce{Ca^2+}$ ציטוזולי:
- מנוחה: ~10⁻⁷ M
- אקטיבציה: ~10⁻⁴ M (עלייה של 3 סדרי גודל בתוך מילי-שניות)
ב־SR קיים Calsequestrin - חלבון שקושר בערך 40 יוני $\ce{Ca^2+}$ למולקולה ומאפשר את ריכוז הסידן הגבוה ב־SR.
Twitch, Summation וטטנוס
Twitch - התכווצות אחת של שריר; נמשכת כ־100-80 מילי-שניות (תלוי בסוג השריר).
כיצד שריר מעלה כוח?
מסלול 1 - Summation (סכימה): אם גירוי חדש מגיע לפני שהטוויץ’ הקודם נגמר ← הכוחות מסתכמים. בתדר גבוה מספיק ← Tetanus (טטנוס/פעילות טטנית) - כוח מקסימלי, מתמשך ויציב.
שריר שלד אין לו תקופה רפלקטורית מכנית כמו שריר הלב - לכן ניתן לסכום טוויץ’ים ולהגיע לטטנוס. כמעט כל פעילות רצופה שלנו היא פעילות טטנית.
מסלול 2 - גיוס יחידות מוטוריות: הפעלה של יותר יחידות מוטוריות בו־זמנית ← יותר תאי שריר פעילים ← כוח כולל גדול יותר.
- פעולות עדינות (Fine Motor): יחידות מוטוריות בודדות בטטנוס
- פעולות גסות/חזקות: גיוס מסיבי של יחידות מוטוריות
שריר חלק
לעומת שריר מסורטט/מפוספס (שריר שלד) - שריר חלק אינו מפוספס (אין סרקומרים מסודרים) ולכן נקרא “חלק”.
היכן נמצא שריר חלק?
- דרכי אוויר - ספאזם בתגובה לאסטמה ואלרגיה; הרחבה/כיווץ של דרכי הנשימה
- כלי דם - שמירה על טונוס ולחץ דם; בקרת זרימת דם לפריפריה
- מערכת העיכול - פריסטלטיקה (כיווץ גלי להעברת מזון)
- האישון - פתיחה וסגירה של האישון
- רחם, שלפוחית השתן, מערכת הנשימה
כל השרירים האלה הם לא־רצוניים - אינם בשליטה מודעת.
מבנה שריר חלק
- תאים בודדים (לא Multinucleated!) - כל תא עם גרעין אחד
אקטיןומיוזיןקיימים אך לא מסודרים בסרקומרים (לכן אין מראה מפוספס במיקרוסקופ)- בשריר חלק אין מערכת T-tubules מפותחת כמו בשריר שלד; במקום זאת קיימות caveolae
- ה־SR קיים אך הרשת פחות מסועפת (התאים קטנים יותר ואקטיבציה מקומית מספיקה)
- תאים קטנים יותר - אין צורך בנטוורק עמוק כמו בשריר שלד
סוגי שריר חלק
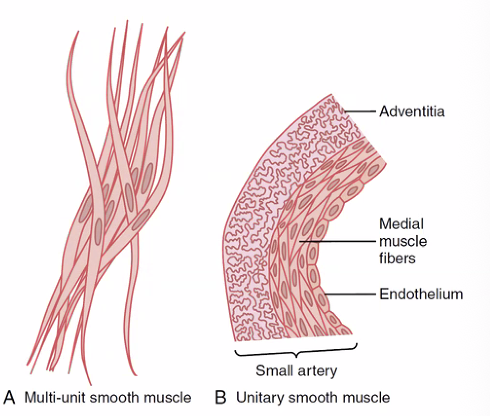
| סוג | מאפיינים | דוגמאות |
|---|---|---|
| Unitary (Single-Unit) | תאים מחוברים בGap Junctions; מופעלים כמקשה אחת; דה־פולריזציה מתפשטת בין תאים | מעי, רחם, שלפוחית השתן, כלי דם קטנים |
| Multi-Unit | כל תא מקבל עצבוב עצמאי; אפשרות לבקרה עדינה ומדויקת | שרירי עין (Ciliary body, Iris), שרירי שערות (Arrector pili) |
Gap Junctions בשריר חלק יוניטרי דומים למה שקיים בשריר הלב - תעלות המחברות ציטוזול של שני תאים ומאפשרות מעבר חשמלי ישיר.
מנגנון ההתכווצות בשריר חלק - תפקיד הסידן
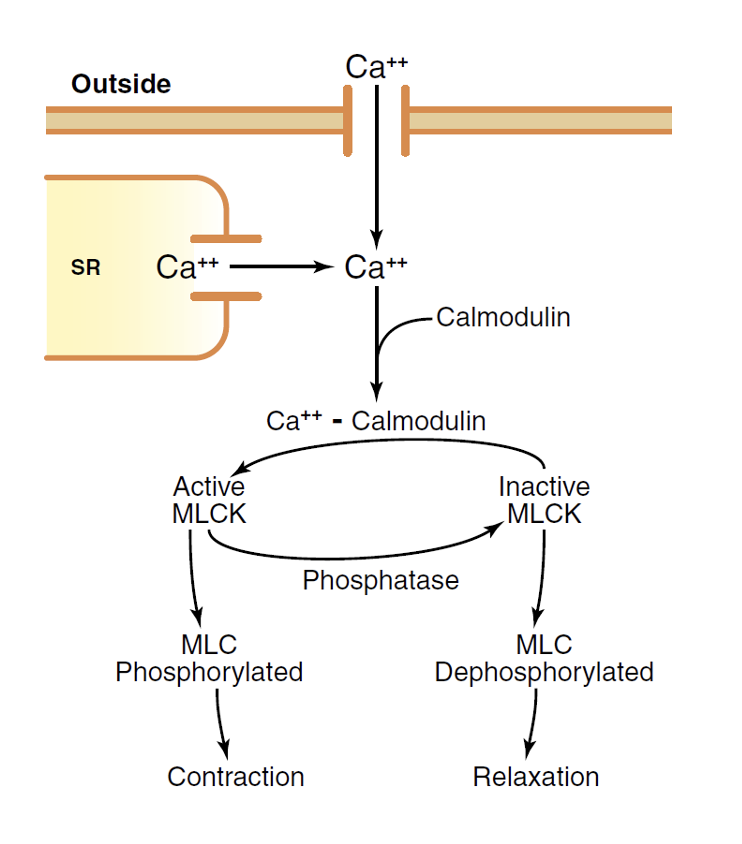
בשריר חלק אין טרופונין. הבקרה על ידי סידן מתבצעת דרך Calmodulin:
- עליית $\ce{Ca^2+}$ ציטוזולי (משחרור מה־SR + כניסה מחוץ לתא דרך Voltage-Gated $\ce{Ca^2+}$ Channels)
- $\ce{Ca^2+}$ נקשר לCalmodulin
- קומפלקס $\ce{Ca^2+}$-Calmodulin מפעיל Myosin Light Chain Kinase (MLCK)
- MLCK מבצע פוספורילציה של Light Chains על ראש ה
מיוזין - פוספורילציה ← הפעלת ההידרוליזה של ATP בראש ה
מיוזין← Power Stroke ו־Cross-Bridge Cycling
הבדל מרכזי:
- בשריר שלד - בקרת הסידן היא על
אקטין(דרך טרופונין).- בשריר חלק - בקרת הסידן היא על
מיוזין(דרך Calmodulin + MLCK).
רלקסציה בשריר חלק
- $\ce{Ca^2+}$ מוחזר ל־SR לאט יחסית
- ניתוק $\ce{Ca^2+}$ מ־Calmodulin - קינטיקה איטית יותר
- ← הרלקסציה ארוכה יותר מאשר בשריר שלד
משך ההתכווצות
| סוג שריר | משך טוויץ’/התכווצות |
|---|---|
| שריר שלד (מהיר) | כ־50-10 מילי-שניות |
| שריר שלד (איטי) | כ־100-80 מילי-שניות |
| שריר לב | כ־500-200 מילי-שניות |
| שריר חלק | שניות עד עשרות שניות |
היכולת של שריר חלק לשמור על התכווצות ארוכה ויציבה מאפשרת שמירה על טונוס כלי דם ופעילות פריסטלטית ממושכת.
סוגי התכווצות בשריר חלק
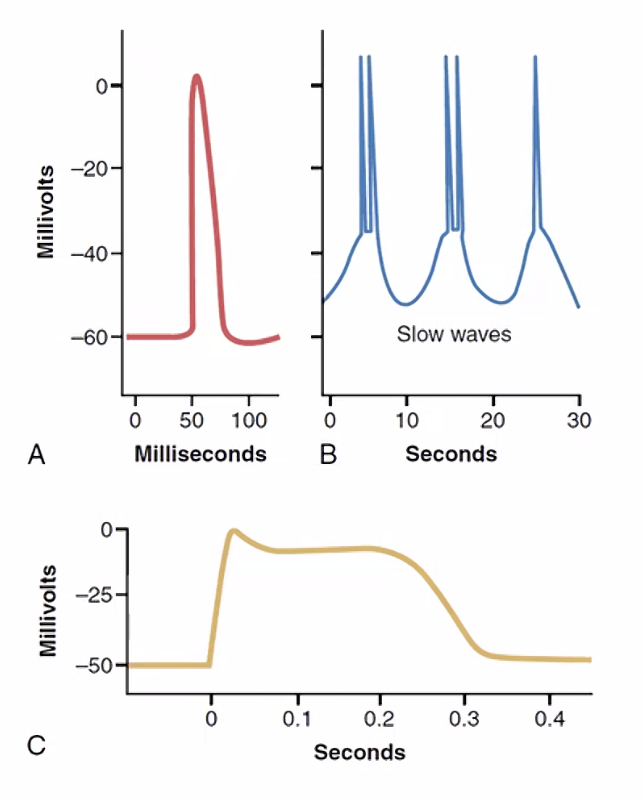
- Phasic Contraction - התכווצות קצרה יחסית; אופייני לשרירי העין, Arrector pili
- Tonic Contraction - שמירה על טונוס ממושך לאורך שניות ועשרות שניות; אופייני לכלי דם ולמעי
גורמי אקטיבציה של שריר חלק
- עצבוב אוטונומי - Acetylcholine ו־Norepinephrine (בכלי דם)
- גורמים הורמונליים
- מכני - Stretch Activation - מתיחה מפעילה תעלות מכאנו־רגישות; לדוגמה: מילוי הקיבה באוכל מפעיל פריסטלטיקה
סיכום השוואתי - שריר שלד לעומת שריר חלק
| מאפיין | שריר שלד | שריר חלק |
|---|---|---|
| מבנה | מפוספס, סרקומרים מסודרים | חלק, ללא סרקומרים |
| גרעינים לתא | מרובים (Multinucleated) | אחד לתא |
| בקרת $\ce{Ca^2+}$ | דרך Troponin C על האקטין | דרך Calmodulin + MLCK על המיוזין |
| T-Tubules | עמוקים ומפותחים | קוויולות קטנות בלבד |
| Gap Junctions | אין | יש (ב־Unitary) |
| שליטה | רצונית | לא־רצונית |
| משך התכווצות | עשרות מילי-שניות | שניות עד עשרות שניות |
| טטנוס | אפשרי | לא קיים |
| Excitation | נוירון מוטורי (ACh) | עצבוב + הורמונים + מתיחה |