תרגום: שלושת השחקנים המרכזיים
תרגום הוא השלב שבו המידע שנמצא ב־mRNA הופך לרצף חומצות אמינו. שלושת רכיבי ה־RNA שנלמדו בשיעורים הקודמים מתכנסים כאן לתהליך אחד:
mRNA - התבנית. הרצף שלו נקרא בכיוון $5’ \to 3’$ ומכיל את הקוד לחלבון.
rRNA - מרכיב מרכזי של הריבוזום. הריבוזום הוא המקום שבו הקודון ב־mRNA מותאם ל־anticodon של tRNA ונוצר הקשר הפפטידי.
tRNA - מולקולת מתאם. מצד אחד היא מזהה קודון ב־mRNA, ומצד שני היא נושאת חומצת אמינו.
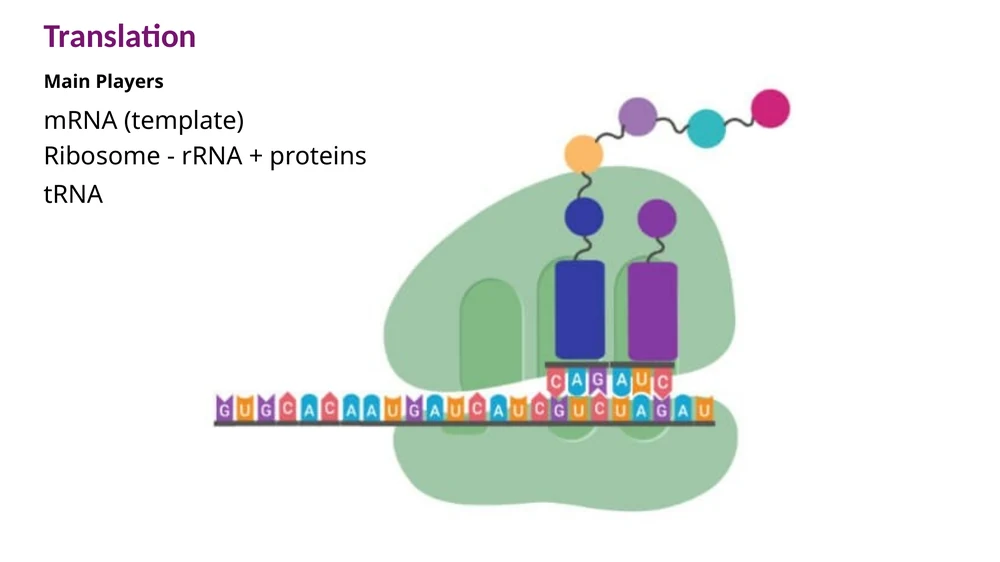
החלבון נבנה בכיוון N-terminus → C-terminus. לכן, כאשר קוראים mRNA מ־$5’$ ל־$3’$, רצף החומצות האמיניות שנוצר מתחיל בקצה האמיני ומתקדם אל הקצה הקרבוקסילי.
לזכור למבחן: ה־mRNA נקרא $5’ \to 3’$, והחלבון נבנה מ־N-terminus ל־C-terminus.
הקוד הגנטי
הריבוזום קורא את ה־mRNA בשלשות נוקלאוטידים. כל שלשה נקראת codon, וכל codon מתאים לחומצת אמינו מסוימת או לסיום תרגום.
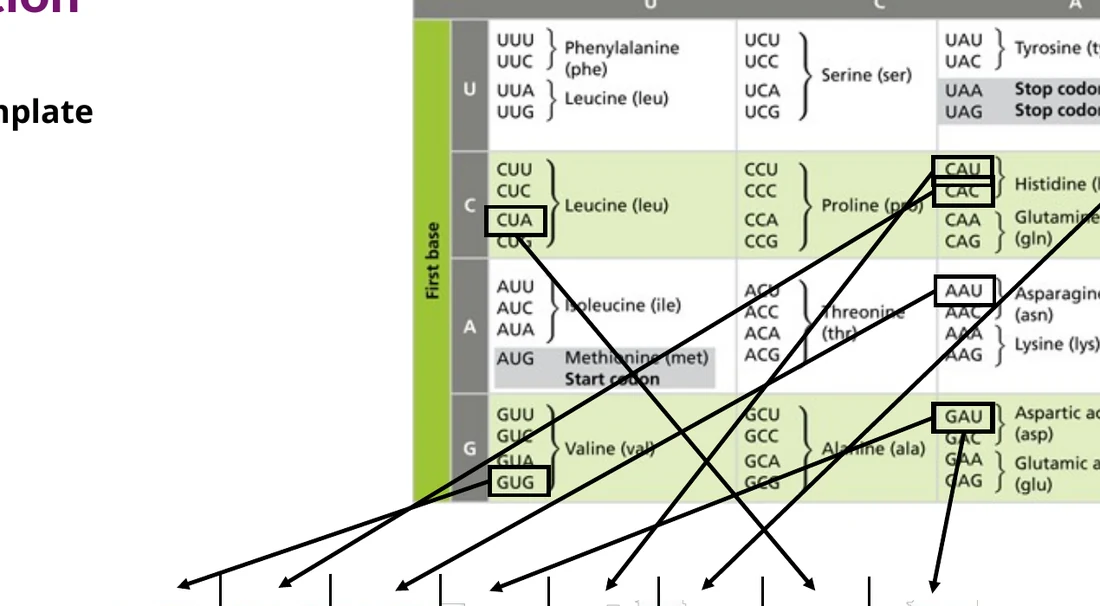
יש $4^3 = 64$ codons אפשריים. מתוכם:
| קבוצה | משמעות |
|---|---|
61 codons | מקודדים ל־20 חומצות אמינו |
AUG | Start codon, מקודד ל־Methionine |
UAA, UAG, UGA | Stop codons |
רוב חומצות האמינו מקודדות על ידי יותר מקודון אחד. לדוגמה, Alanine ו־Glycine יכולות להיות מקודדות על ידי כמה codons שונים. התכונה הזו נקראת degeneracy של הקוד הגנטי.
לזכור למבחן: 64 קודונים, מתוכם 61 מקודדים חומצות אמינו ו־3 הם stop codons. הקודון AUG מקודד ל־Methionine ומשמש בדרך כלל כ־start codon.
tRNA: מולקולת מתאם בין קודון לחומצת אמינו
ה־tRNA מתקפל למבנה המזכיר עלה תלתן. שני אזורים חשובים במיוחד:
- בקצה $3’$ יש רצף CCA. חומצת האמינו מתחברת ל־A האחרון.
- בלולאת ה־anticodon יש שלושה נוקלאוטידים שמזהים codon ב־mRNA.
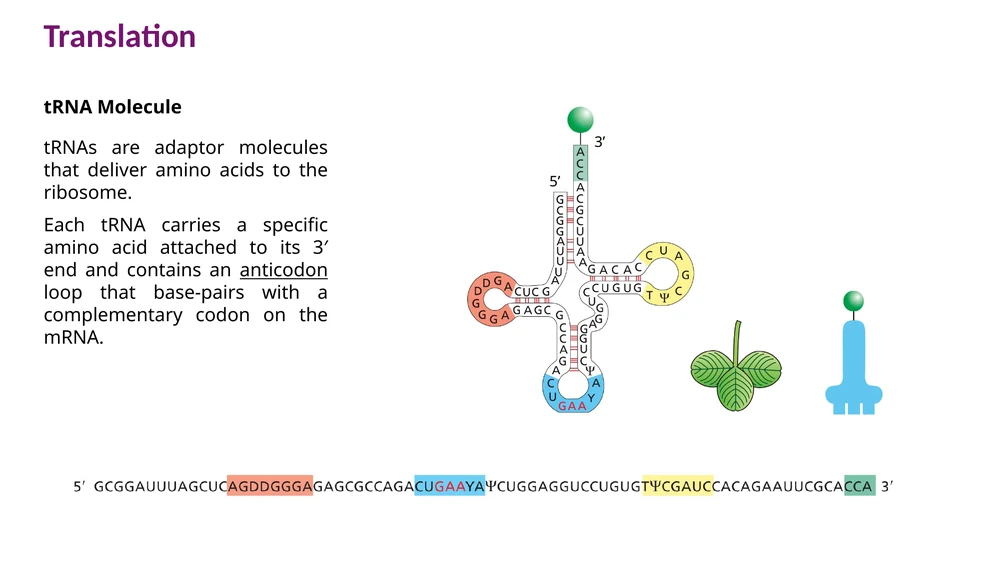
הקישור בין codon ל־anticodon הוא אנטי־מקבילי. לכן, כאשר מציירים tRNA בכיוון $5’ \to 3’$, צריך לשים לב שה־anticodon מזווג ל־codon בכיוון ההפוך.
לדוגמה, codon ב־mRNA:
5'-UUC-3'
יוכר על ידי anticodon משלים בכיוון אנטי־מקבילי:
3'-AAG-5'
בכתיבה רגילה של רצף RNA, אם רוצים לכתוב את ה־anticodon בכיוון $5’ \to 3’$, הוא ייכתב הפוך.
Wobble: גמישות בעמדה השלישית של הקודון
בתא יש פחות סוגי tRNA ממספר ה־codons שמקודדים לחומצות אמינו. באדם יש בערך 48 סוגי tRNA שמספיקים לקריאת 61 codons. זה מתאפשר בזכות Wobble base-pairing.
הזיווג המדויק ביותר נדרש בשתי העמדות הראשונות של ה־codon. בעמדה השלישית יש יותר גמישות. כך tRNA אחד יכול לזהות יותר מקודון אחד, בדרך כלל codons שמקודדים לאותה חומצת אמינו.
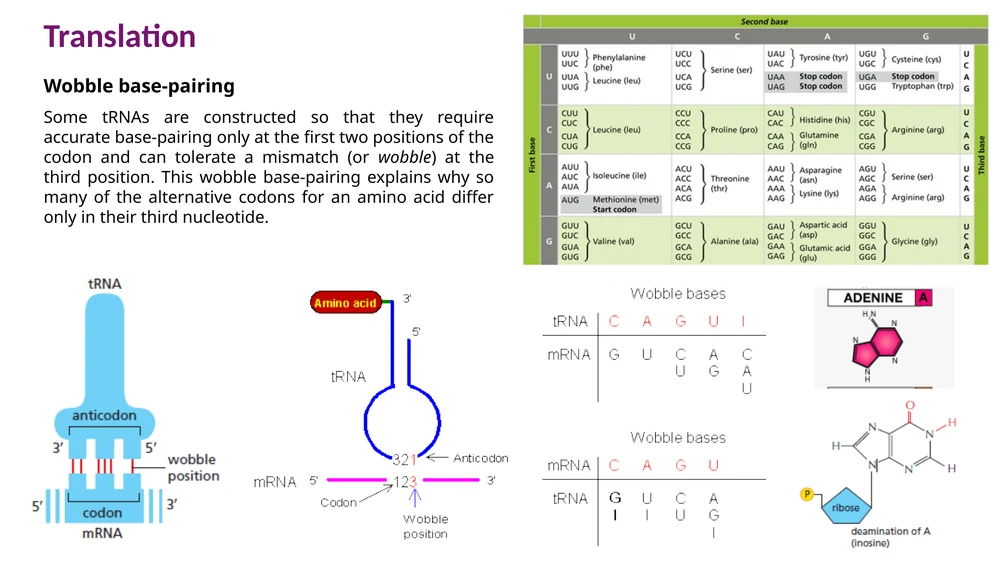
דוגמה חשובה היא inosine. כאשר inosine נמצא בעמדת ה־wobble של anticodon, הוא יכול להזדווג עם כמה בסיסים שונים ב־mRNA, למשל U, C או A. כך התא מצמצם את מספר מולקולות ה־tRNA הדרושות לקריאת הקוד.
הגמישות של wobble מתרחשת בעיקר בעמדה השלישית של ה־codon. שתי העמדות הראשונות שומרות על דיוק הקריאה של חומצת האמינו.
טעינת tRNA בחומצת אמינו
לפני ש־tRNA נכנס לריבוזום, הוא צריך להיות טעון בחומצת האמינו המתאימה. התהליך נעשה על ידי aminoacyl-tRNA synthetase.
לכל חומצת אמינו יש בדרך כלל synthetase מתאים. לכן יש בערך 20 אנזימים כאלה, אחד לכל חומצת אמינו סטנדרטית.
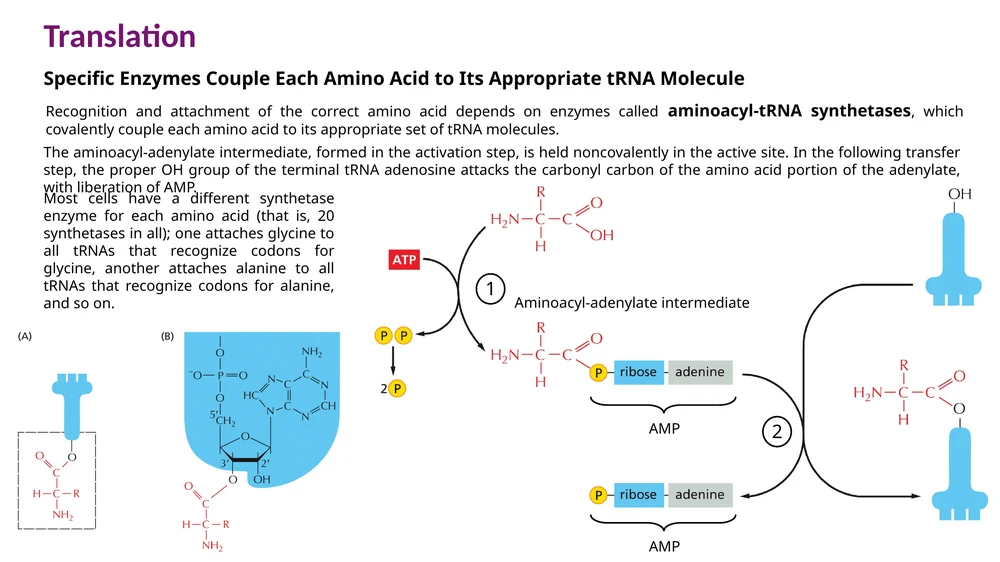
התהליך מתרחש בשני שלבים:
- האנזים קושר חומצת אמינו ו־ATP, ויוצר תוצר ביניים שנקרא aminoacyl-adenylate.
- חומצת האמינו מועברת לקצה $3’$ של ה־tRNA, אל קבוצת ה־OH של ה־A האחרון ברצף CCA.
הקשר בין חומצת האמינו ל־tRNA הוא קשר עתיר אנרגיה. האנרגיה הזו תנוצל בהמשך ליצירת הקשר הפפטידי בתוך הריבוזום.
דיוק הטעינה
דיוק התרגום תלוי בשתי בדיקות:
- התאמת חומצת האמינו ל־tRNA על ידי aminoacyl-tRNA synthetase.
- התאמת ה־anticodon של ה־tRNA ל־codon ב־mRNA בתוך הריבוזום.

בחלק מהאנזימים יש גם editing site. האתר הזה מזהה טעינה שגויה, למשל כאשר חומצת אמינו דומה מאוד נכנסה בטעות, ומפרק את הקשר לפני שה־tRNA יגיע לריבוזום.
לזכור למבחן: tRNA טעון נקרא aminoacyl-tRNA. האנזים שטוען אותו הוא aminoacyl-tRNA synthetase, והדיוק שלו קריטי כי הריבוזום מזהה בעיקר את ה־anticodon, לא את זהות חומצת האמינו המחוברת.
יצירת הקשר הפפטידי
החלבון מתארך על ידי יצירת קשר פפטידי בין:
- הקבוצה הקרבוקסילית בקצה השרשרת הגדלה.
- הקבוצה האמינית של חומצת האמינו החדשה.
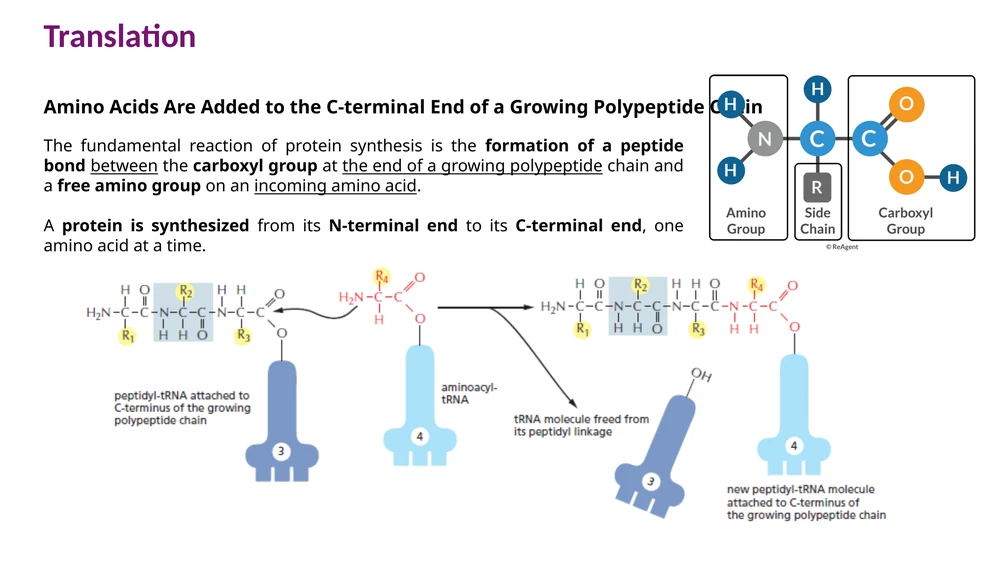
השרשרת הגדלה עוברת בכל מחזור אל ה־tRNA החדש שנמצא באתר A. לאחר מכן הריבוזום מתקדם codon אחד, והאתרים מסתדרים למחזור הבא.
מבנה הריבוזום ואתרי A, P, E
הריבוזום מורכב מתת־יחידה קטנה ומתת־יחידה גדולה. באאוקריוטים התת־יחידה הקטנה היא 40S והתת־יחידה הגדולה היא 60S, וביחד מתקבל ריבוזום 80S.
לתת־היחידה הקטנה יש אתר קישור ל־mRNA. כאשר שתי התת־יחידות מתחברות, נוצרים שלושה אתרי קישור ל־tRNA:
| אתר | שם | תפקיד |
|---|---|---|
| A site | Aminoacyl site | כניסת aminoacyl-tRNA חדש |
| P site | Peptidyl site | החזקת ה־tRNA שעליו נמצאת השרשרת הגדלה |
| E site | Exit site | יציאת tRNA שכבר מסר את חומצת האמינו |
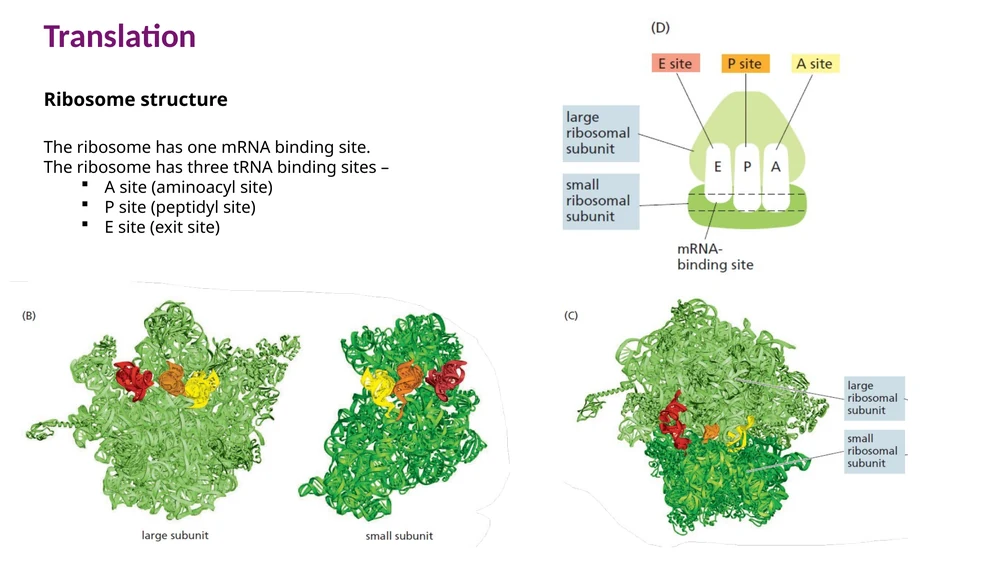
ה־initiator tRNA עם Methionine נכנס ישירות ל־P site בתחילת התרגום. שאר ה־tRNAs נכנסים במהלך elongation דרך A site.
שלבי התרגום
תהליך התרגום מחולק לשלושה שלבים:
- Initiation - מציאת start codon והרכבת הריבוזום.
- Elongation - כניסת tRNAs, יצירת קשרים פפטידיים ותזוזת הריבוזום.
- Termination - זיהוי stop codon, שחרור החלבון ופירוק הקומפלקס.
פקטורים מרכזיים בתרגום
| שלב | פקטורים | תפקיד |
|---|---|---|
| Initiation | eIF4E | קושר את ה־5′ cap |
| Initiation | eIF4G | קושר את eIF4E ואת PABP על poly-A tail ויוצר לולאה של mRNA |
| Initiation | eIF2-GTP | מביא את initiator Met-tRNAi לתת־היחידה הקטנה |
| Elongation | eEF1A-GTP | מביא aminoacyl-tRNA חדש ל־A site |
| Elongation | eEF2-GTP | מקדם translocation של הריבוזום |
| Termination | eRF1 | מזהה stop codons |
| Termination | eRF3-GTP | מסייע בשחרור הפוליפפטיד ובסיום התהליך |
Initiation: התחלת התרגום
שלב ה־initiation מתחיל ביצירת קומפלקס על תת־היחידה הקטנה של הריבוזום. הקומפלקס כולל:
- תת־יחידה קטנה של הריבוזום
- initiator Met-tRNAi
- eIF2-GTP
- פקטורי initiation נוספים
במקביל, ה־mRNA עובר activation:
- eIF4E נקשר ל־5′ cap.
- PABP נקשר ל־poly-A tail.
- eIF4G מקשר בין eIF4E לבין PABP, וכך מקרב את קצה $5’$ ואת קצה $3’$ של ה־mRNA.
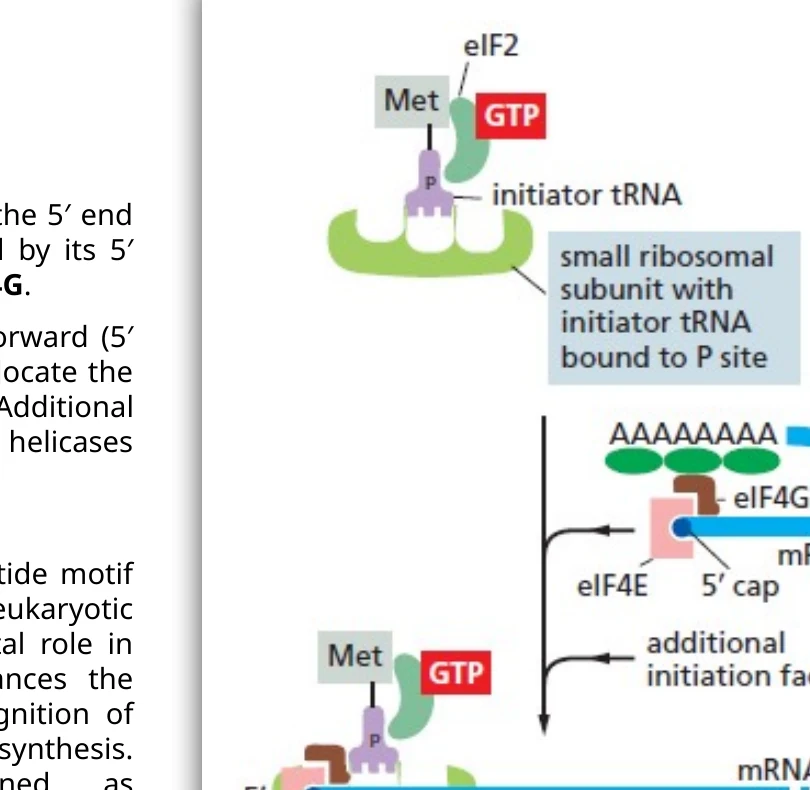
לאחר הקישור לקצה $5’$, תת־היחידה הקטנה סורקת את ה־5′ UTR בכיוון $5’ \to 3’$ עד שהיא מזהה AUG בהקשר מתאים.
Kozak sequence
ב־mRNA אאוקריוטי, זיהוי יעיל של start codon תלוי גם ברצף שמסביב ל־AUG. הרצף הזה נקרא Kozak sequence.
רצף הקונצנזוס:
5′-GCC(A/G)CCAUGG-3′
שתי עמדות חשובות במיוחד:
- A או G בעמדה $-3$ לפני ה־AUG.
- G בעמדה $+4$ אחרי ה־AUG.
כאשר ההקשר של Kozak חזק, ה־AUG מזוהה ביעילות. כאשר ההקשר חלש, תת־היחידה הקטנה יכולה להמשיך לסרוק ולבחור AUG מאוחר יותר. תופעה זו נקראת leaky scanning.
לזכור למבחן: באאוקריוטים תת־היחידה הקטנה נקשרת לקצה $5’$ של ה־mRNA וסורקת עד AUG. רצף Kozak טוב מעלה את יעילות הזיהוי של start codon.
לאחר זיהוי AUG, מתרחשת הידרוליזה של GTP על eIF2. פקטורי ה־initiation עוזבים, תת־היחידה הגדולה מצטרפת, וה־initiator Met-tRNAi נשאר ב־P site. מכאן מתחיל elongation.
Elongation: הארכת השרשרת
בכל מחזור elongation נכנסת חומצת אמינו אחת חדשה לשרשרת.
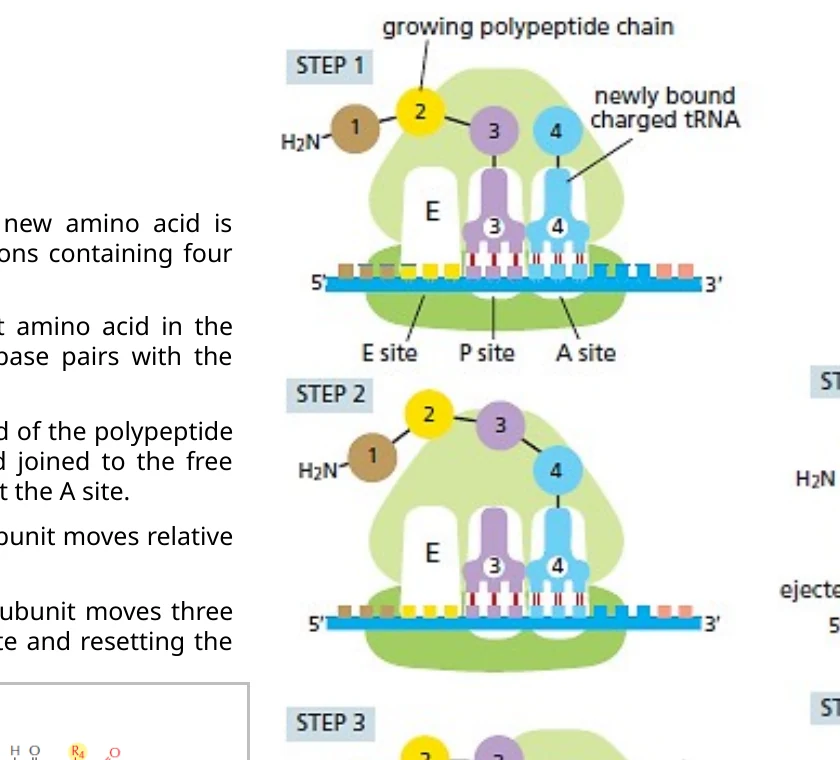
המחזור כולל ארבעה צעדים:
- כניסת aminoacyl-tRNA ל־A site בעזרת eEF1A-GTP.
- בדיקת codon-anticodon. התאמה טובה גורמת להידרוליזת GTP ולעזיבת eEF1A.
- יצירת קשר פפטידי. השרשרת עוברת מה־tRNA ב־P site אל חומצת האמינו שעל ה־tRNA ב־A site.
- Translocation. הריבוזום מתקדם codon אחד. ה־tRNA שהיה ב־P site עובר ל־E site ויוצא, וה־tRNA עם השרשרת עובר ל־P site.
Proofreading בזמן elongation
דיוק התרגום מתבסס על שני שלבי בדיקה:
- eEF1A-GTP מכניס aminoacyl-tRNA ל־A site. התאמה טובה בין codon ל־anticodon מפעילה הידרוליזת GTP.
- לאחר עזיבת eEF1A, הריבוזום עובר שינוי מבני. tRNA מתאים מתייצב באתר A, ו־tRNA לא מתאים נוטה להשתחרר לפני יצירת קשר פפטידי.
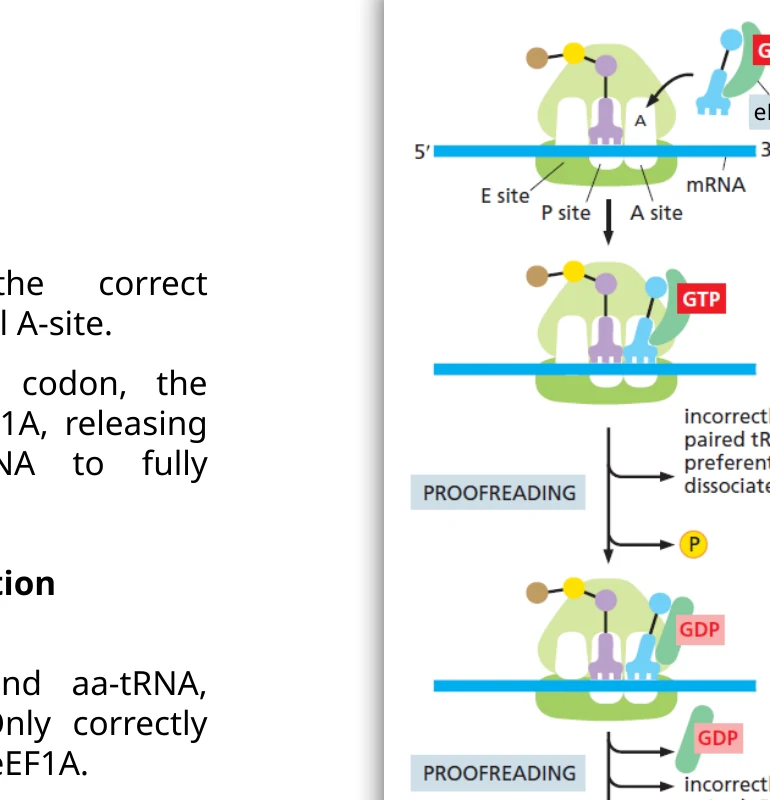
בדיקת הריבוזום מתמקדת בהתאמת codon-anticodon. לכן הטעינה הנכונה של ה־tRNA בחומצת אמינו נעשית מוקדם יותר, על ידי aminoacyl-tRNA synthetase.
Translocation
אחרי יצירת הקשר הפפטידי, תת־היחידה הגדולה זזה ביחס ל־tRNA. לאחר מכן eEF2-GTP מקדם את התזוזה של תת־היחידה הקטנה ושל ה־mRNA בשלושה נוקלאוטידים.
כך ה־A site מתפנה לקליטת aminoacyl-tRNA נוסף, והמחזור חוזר על עצמו.
Termination: סיום התרגום
כאשר stop codon נכנס ל־A site, אין tRNA רגיל שמתאים אליו. במקום tRNA מגיעים release factors.
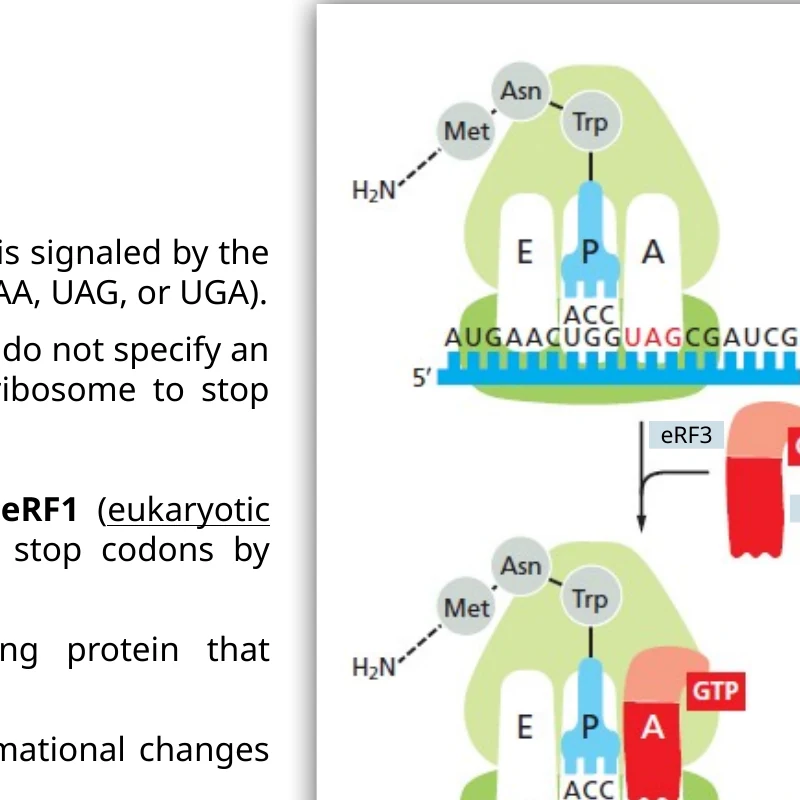
שלבי הסיום:
- eRF1 מזהה את אחד ה־stop codons: UAA, UAG או UGA.
- eRF3-GTP נקשר ל־eRF1 ומקדם שינוי קונפורמציה.
- מרכז ה־peptidyl transferase מבצע הידרוליזה של הקשר בין הפוליפפטיד לבין ה־tRNA.
- החלבון משתחרר.
- תת־היחידות של הריבוזום, ה־mRNA וה־tRNA מתפרקים למחזור תרגום נוסף.
לזכור למבחן: stop codons מזוהים על ידי release factors, בעיקר eRF1, ולא על ידי tRNA רגיל.
Polyribosomes
על אותו mRNA יכולים לשבת מספר ריבוזומים בו־זמנית. מבנה כזה נקרא polyribosome או polysome.
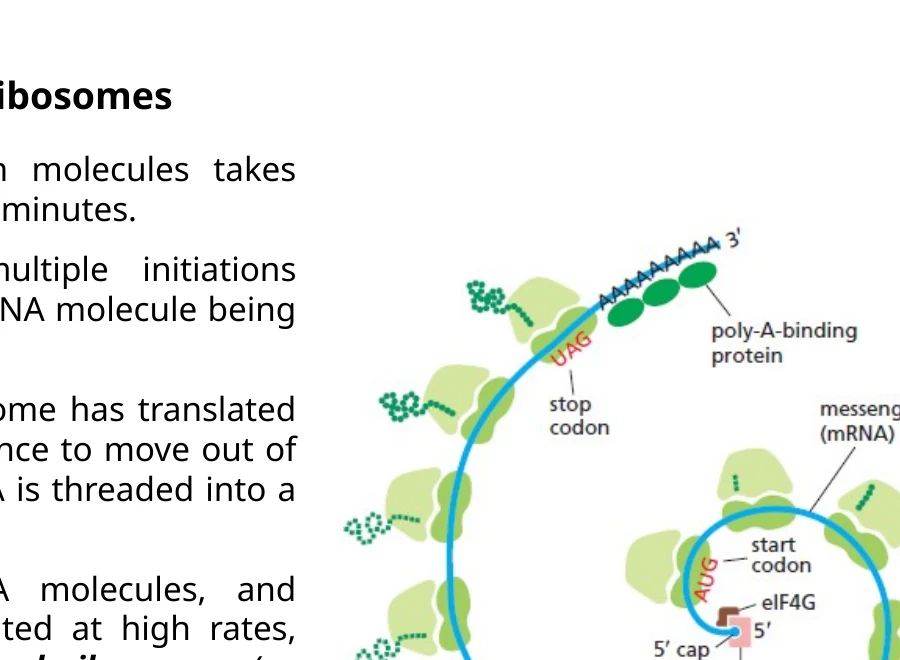
המשמעות היא שמולקולת mRNA אחת יכולה לייצר עותקים רבים של אותו חלבון בזמן קצר. ריבוזום שמתחיל ראשון יהיה קרוב יותר לקצה $3’$, והריבוזומים שמתחילים אחריו יהיו קרובים יותר לקצה $5’$.
אחרי התרגום: החלבון צריך להגיע למבנה פעיל
סיום התרגום נותן פוליפפטיד, אבל חלבון פעיל דורש שלבים נוספים:
- קיפול למבנה תלת־ממדי.
- הרכבה עם תת־יחידות נוספות, אם החלבון פועל כקומפלקס.
- קישור cofactors, למשל יוני מתכת או coenzymes.
- מודיפיקציות post-translational.
- בקרת איכות וסילוק חלבונים שלא התקפלו בצורה תקינה.
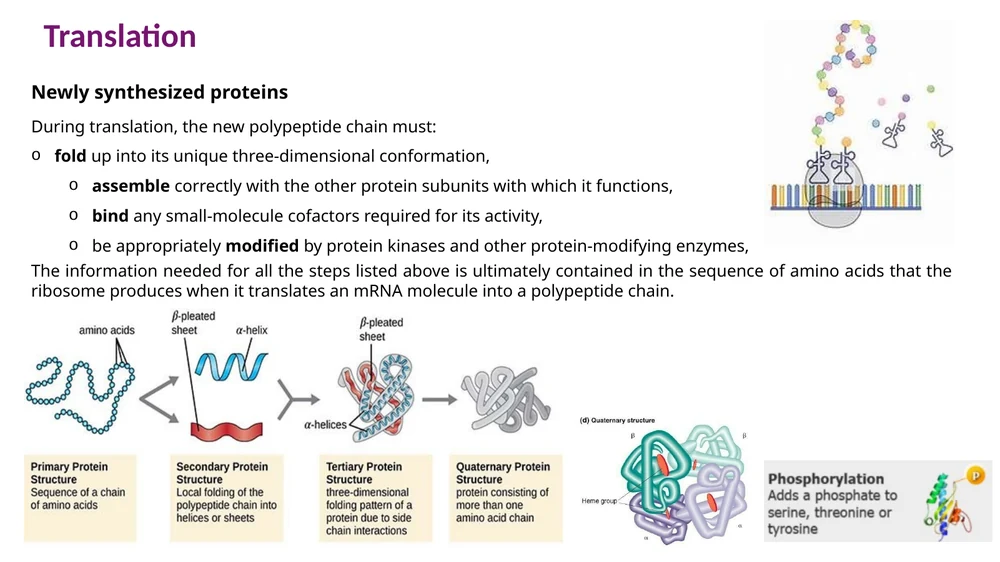
עקרונות קיפול
רצף חומצות האמינו מכיל את המידע הבסיסי לקיפול. במהלך הקיפול:
- אזורים הידרופוביים נוטים להיקבר בליבת החלבון.
- אזורים פולריים וטעונים נוטים להיות חשופים יותר לסביבה מימית.
- קשרי מימן, אינטראקציות יוניות, אינטראקציות הידרופוביות וקשרי Van der Waals מייצבים את המבנה.
הקיפול מתחיל כבר כאשר השרשרת יוצאת מתעלת היציאה של הריבוזום. בחלבונים רבים, Methionine הראשון מוסר לאחר מכן, ולעיתים הקצה ה־N-terminal עובר acetylation.
Cofactors ומודיפיקציות לאחר תרגום
חלבונים רבים זקוקים למרכיבים נוספים כדי לפעול:
| סוג | דוגמאות | תפקיד כללי |
|---|---|---|
| Metal ions | $\ce{Zn^{2+}}$, $\ce{Fe^{2+}/Fe^{3+}}$, $\ce{Cu^{2+}}$, $\ce{Mg^{2+}}$ | יציבות מבנית, קטליזה, קישור מצעים |
| Coenzymes | ביוטין, Coenzyme A, ויטמינים שונים | נשיאת קבוצות כימיות או אלקטרונים |
| Post-translational modifications | phosphorylation, acetylation, methylation, ubiquitination | שינוי פעילות, מיקום, יציבות או אינטראקציות |
מודיפיקציות לאחר תרגום יכולות לשנות במהירות את פעילות החלבון בלי לייצר חלבון חדש מהתחלה. לדוגמה, phosphorylation יכולה להפעיל או לעכב אנזים, ו־ubiquitination יכולה לסמן חלבון לפירוק או לשנות את פעילותו, תלוי בסוג השרשרת.
Chaperones: עזרה בקיפול ומניעת אגרגציה
חלבונים צעירים או חלבונים שנפגעו יכולים לחשוף אזורים הידרופוביים. אזורים כאלה נוטים להידבק לחלבונים אחרים וליצור aggregates. ה־Molecular chaperones מזהים מצבים כאלה ונותנים לחלבון הזדמנויות חוזרות לקיפול נכון.
Hsp70
Hsp70 הוא chaperone שמור מאוד. הוא נקשר לרצפים קצרים והידרופוביים בחלבון שעדיין לא התקפל בצורה יציבה.
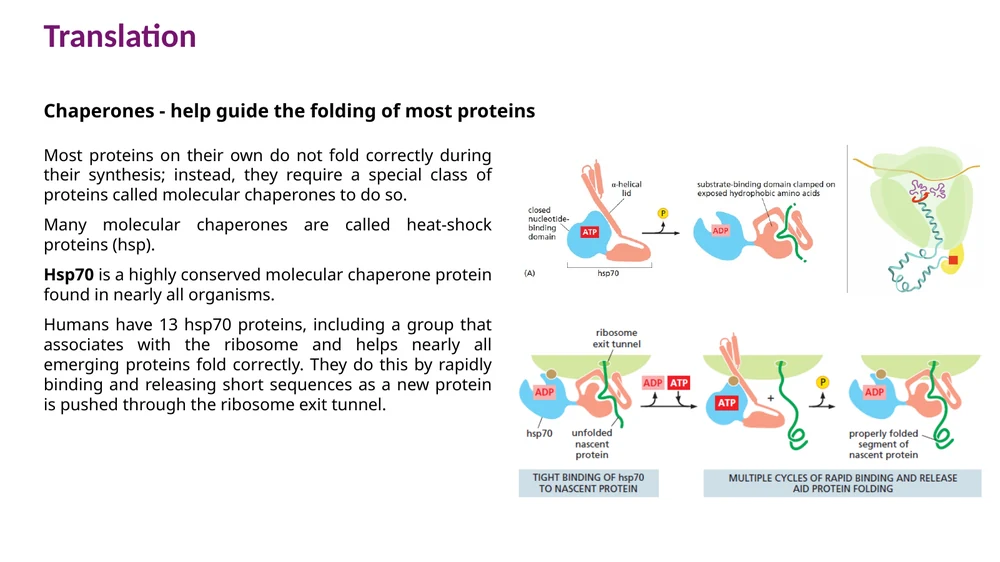
מחזור הפעולה:
- Hsp70 מזהה אזור הידרופובי חשוף.
- הידרוליזת ATP ל־ADP סוגרת את Hsp70 על החלבון ושומרת אותו במצב פרוש יחסית.
- קישור ATP חדש פותח את Hsp70 ומשחרר את החלבון.
- המחזור חוזר עד שהחלבון מצליח לקפל את האזורים ההידרופוביים פנימה או עד שהוא מועבר למסלול בקרת איכות אחר.
הפעולה הזו מצמצמת אגרגציה ונותנת לחלבון זמן לקיפול הדרגתי.
Hsp90 ו־Hsp60
חלק מהחלבונים זקוקים לעזרה נוספת אחרי Hsp70.
Hsp90 פועל בעיקר בשלבים מאוחרים של קיפול ומייצב חלבוני לקוח כמו kinases, steroid hormone receptors ו־transcription factors. הוא עובד כדימר ובמחזור תלוי ATP.
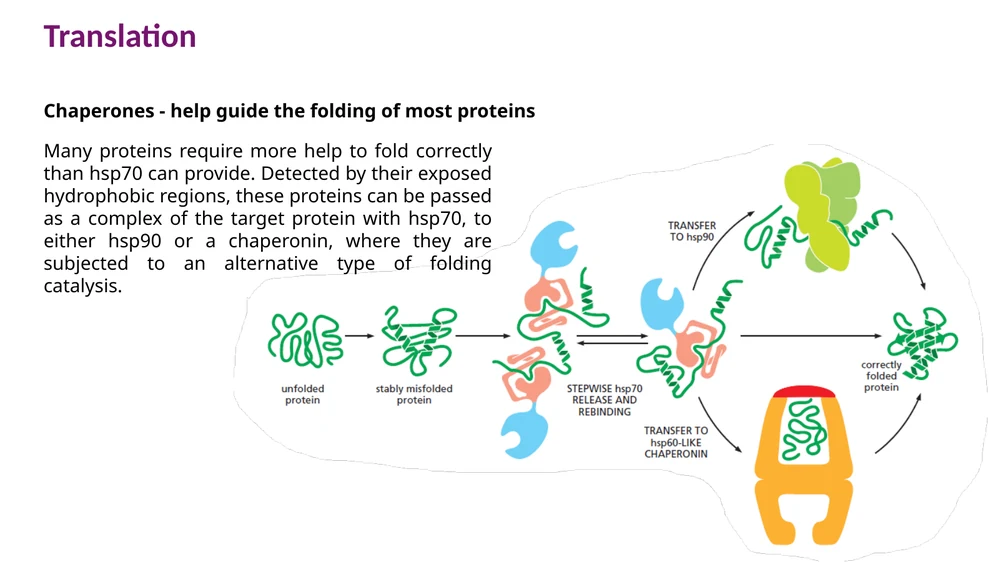
Hsp60 / chaperonin יוצר תא קיפול מבודד. החלבון נכנס לחלל, מכסה נסגר בתהליך תלוי ATP, והחלבון מקבל סביבה מבודדת שבה הוא יכול לנסות להתקפל מחדש.
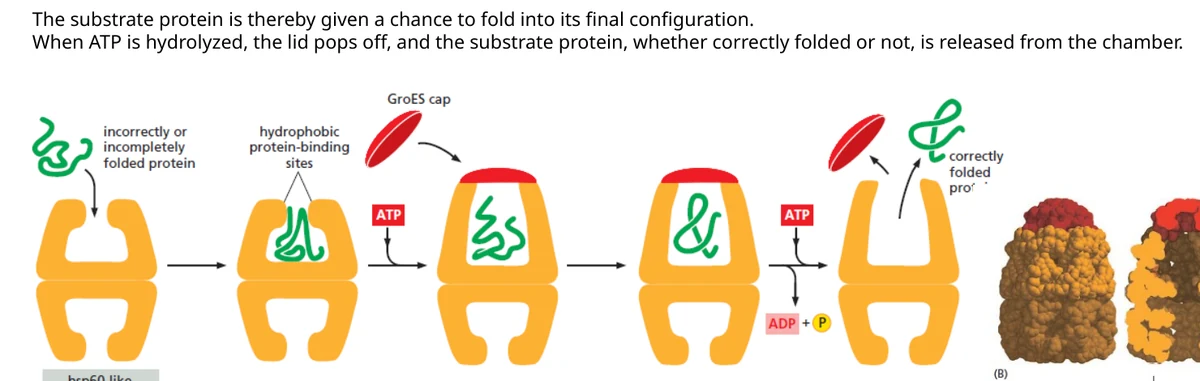
לזכור למבחן: Hsp70 מתאים לזיהוי מוקדם של אזורים הידרופוביים חשופים; Hsp60/chaperonin מספק תא קיפול מבודד; Hsp90 מלווה maturation של חלבוני signaling ולקוחות ספציפיים.
סילוק חלבונים שלא התקפלו נכון
כאשר חלבון ממשיך להציג אזורים הידרופוביים חשופים או לא מצליח להגיע למבנה תקין, התא מסמן אותו לפירוק. הסימון המרכזי לפירוק דרך proteasome הוא שרשרת polyubiquitin המקושרת דרך Lys48.
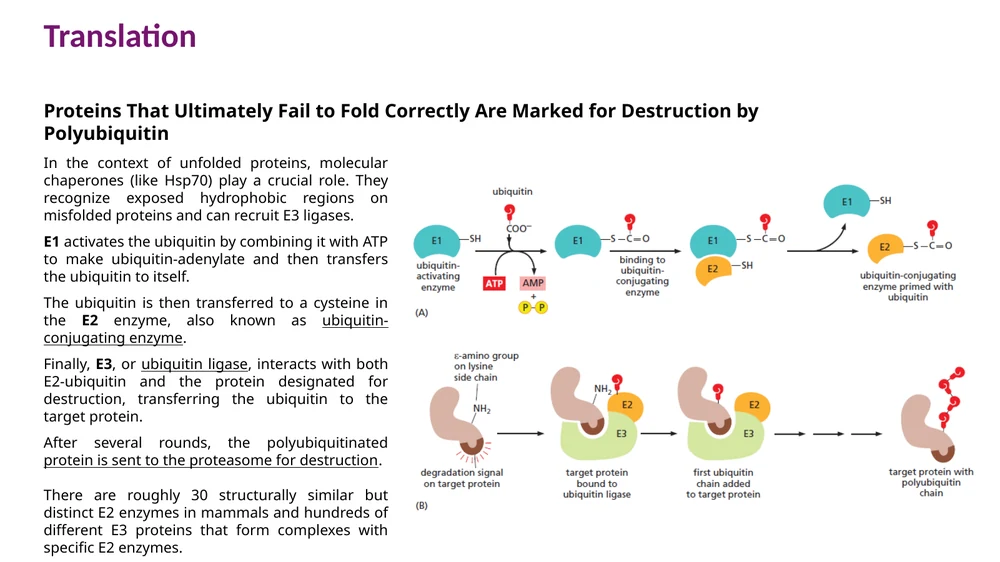
שלבי הסימון:
- E1 מפעיל ubiquitin בעזרת ATP.
- ubiquitin עובר ל־E2.
- E3 ligase מזהה את חלבון המטרה ומביא אותו לקרבת E2.
- ubiquitin מתחבר לליזין על חלבון המטרה.
- מחזורים נוספים יוצרים שרשרת polyubiquitin.
E3 ligases נותנים למסלול את הספציפיות. בתאים יש מאות E3 שונים, ולכן אפשר לזהות מגוון רחב של חלבונים לפירוק.
Ubiquitin יחיד יכול לשמש גם כסימון רגולטורי. שרשרת polyubiquitin דרך Lys48 היא הסימון הקלאסי לפירוק ב־proteasome.
Proteasome
ה־proteasome הוא קומפלקס פירוק חלבונים. הוא בנוי מ־20S core שבו נמצאים אתרי הפרוטאזות, ומשני כובעי 19S שמזהים חלבונים מסומנים.
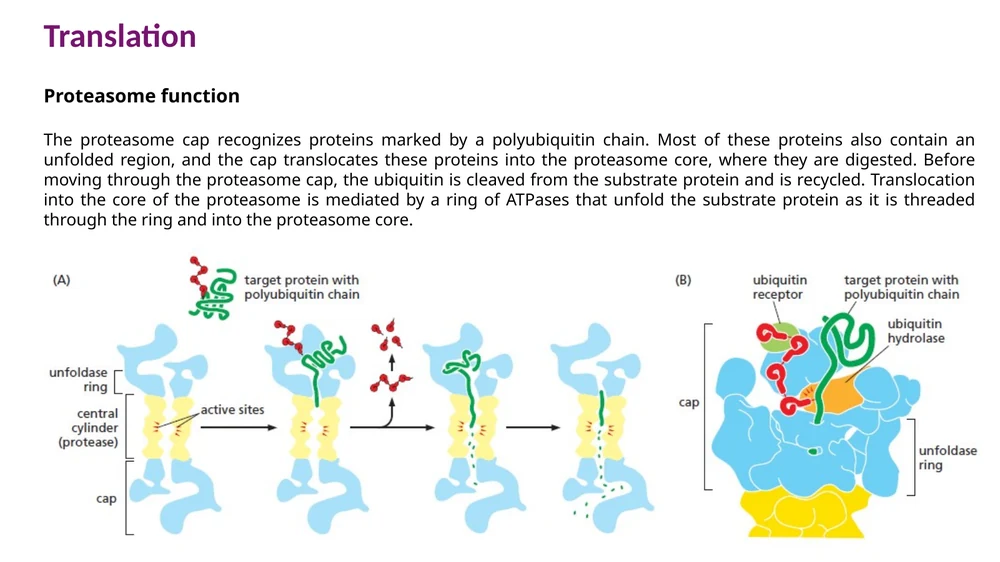
תהליך הפירוק:
- כובע 19S מזהה שרשרת polyubiquitin.
- ubiquitin מוסר וממוחזר.
- טבעת ATPases פורשת את החלבון.
- החלבון מושחל אל 20S core.
- אתרי הפרוטאזות חותכים אותו לפפטידים קצרים.
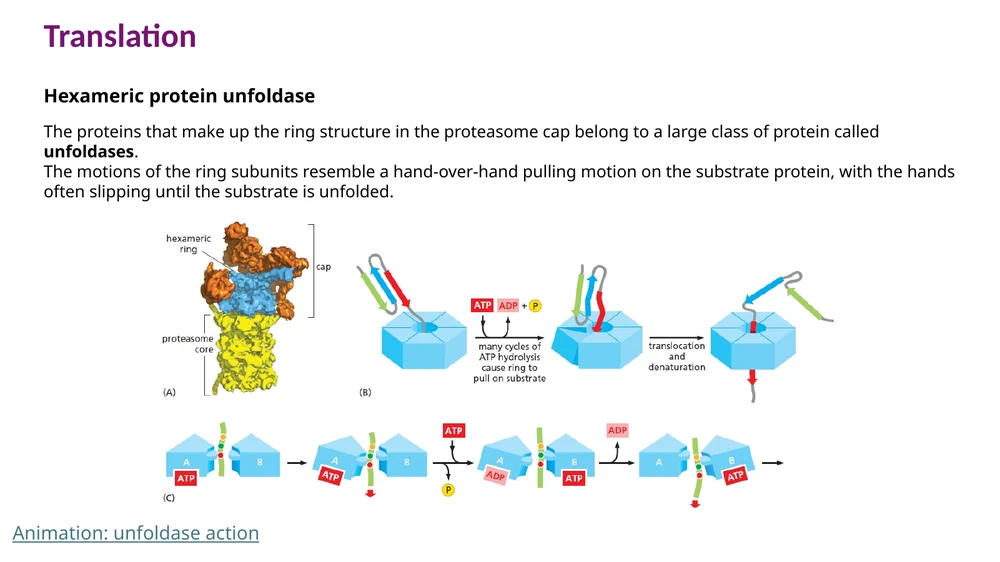
Protein unfoldase
ה־ATPases שבכובע ה־proteasome פועלים כמו unfoldase. הם מושכים את החלבון דרך טבעת צרה, מפרקים את המבנה המקופל שלו, ומשחילים אותו אל הליבה.
מהגן לחלבון פעיל
זרימת המידע מסתיימת בחלבון פעיל רק אחרי כמה שכבות של עיבוד ובקרה:
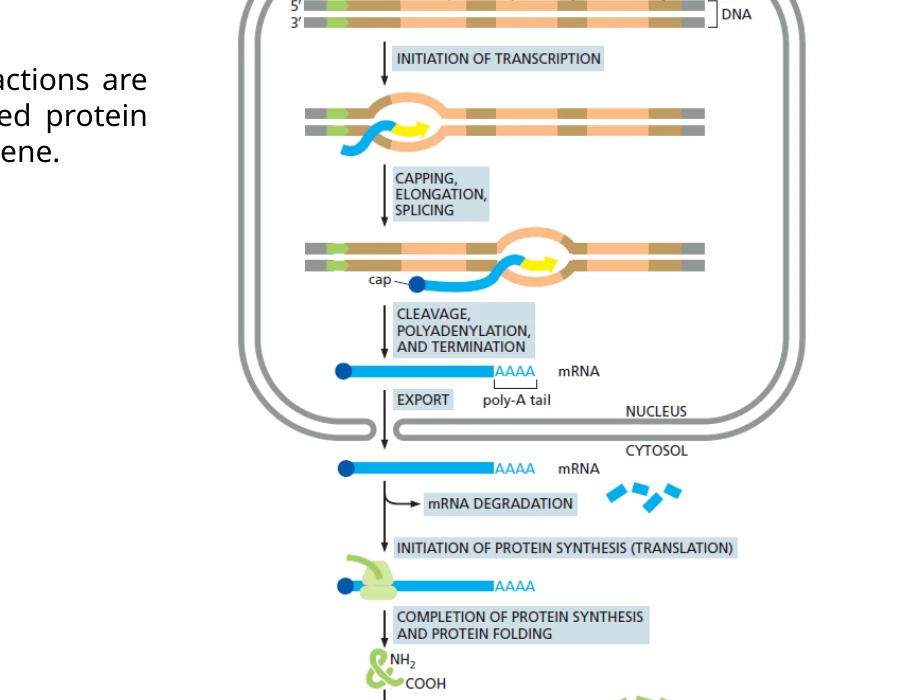
DNA → pre-mRNA: שעתוק בגרעין.
pre-mRNA → mature mRNA: ה־capping, splicing, polyadenylation ובקרת איכות.
mRNA → polypeptide: תרגום בציטופלזמה על ידי ריבוזום, tRNA ופקטורי תרגום.
polypeptide → functional protein: קיפול, cofactors, מודיפיקציות, הרכבה עם תת־יחידות ובקרת איכות.
failed protein → degradation: ה־polyubiquitination דרך Lys48 ופירוק ב־proteasome.