חזרה מסכמת
בתחילת השיעור חזרנו על שאלות מרכזיות מכל החלק של נזקי DNA ותיקוני DNA.
TLS ועמידות לטיפול
תאים סרטניים יכולים להיות עמידים יותר לתרופות שפוגעות ב־DNA כאשר יש בהם פעילות מוגברת של TLS (Translesion Synthesis).
TLS לא מתקן את הנזק, ומאפשר לתא לשכפל מעליו. לכן, אם תרופה כמו cisplatin יוצרת bulky adducts ב־DNA, פעילות מוגברת של TLS יכולה לאפשר לתא להמשיך להתחלק למרות הנזק. זה יוצר סבילות לנזקי DNA ועלול לתרום לעמידות לכימותרפיה.
הקשר בין BER ל־SSB repair
ב־BER (Base Excision Repair) נוצרים לפעמים שברים חד־גדיליים כתוצר ביניים. לדוגמה, אחרי הסרת בסיס פגום ופעילות של AP endonuclease, נוצר nick בגדיל.
לכן יש קשר הדוק בין BER לבין SSBR (Single-Strand Break Repair). אבל ב־BER רגיל התהליך יכול להיות פחות תלוי ב־PARP1, כי יש העברת שרביט ישירה לפקטורים כמו XRCC1.
מה לא שייך ל־BER?
BER הוא מנגנון מאוד ממוקד. הוא מטפל בבסיס פגום אחד או באתר חסר־בסיס.
ב־BER אין הסרה של מקטע DNA ארוך. הסרה של מקטע ארוך מאפיינת יותר:
- NER (Nucleotide Excision Repair)
- MMR (Mismatch Repair)
MSI ו־Lynch syndrome
ב־Lynch syndrome יש פגיעה תורשתית במנגנון MMR. כאשר MMR לא מתפקד, רצפים חוזרניים קצרים כמו microsatellites צוברים טעויות בגלל replication slippage.
התוצאה היא MSI (Microsatellite Instability) - הצטברות שגיאות ברצפי חזרות קצרים. לכן גידולים עם MSI-high נוטים להיות קשורים ל־Lynch syndrome.
MSI משקף הצטברות שגיאות ברצפי חזרות קצרים בגלל פגיעה ב־MMR.
8-oxoG ו־BER
נזק חמצוני לגואנין, למשל 8-oxoG, מתוקן על ידי BER.
הגליקוזילאז הראשון שמזהה את הנזק הוא OGG1. אם הנזק לא מתוקן לפני שכפול, 8-oxoG יכול להזדווג בצורה לא תקינה, ובסוף להוביל למוטציה מסוג: G->T.
AP endonuclease
ב־BER, אחרי שנוצר AP site, צריך להסיר גם את השלד הסוכרי-פוספטי באזור חסר־הבסיס.
התפקיד של AP endonuclease, הוא לחתוך את שלד הסוכר-פוספט באתר חסר־הבסיס, כדי לאפשר לפולימראז למלא את החסר.
NER ו־XP
המחלה התורשתית האופיינית לפגם ב־NER היא XP (Xeroderma Pigmentosum). אנשים עם XP רגישים מאוד לקרני UV, כי הם לא יכולים לתקן דימרים של תימין שנוצרים ב־DNA.
זו הסיבה שחלבוני NER רבים נקראים XPA עד XPG.
XP-V ו־TLS
אם יש רגישות לשמש, אבל NER לא מופיע כאפשרות, אפשר לחשוב על XP-V.
ב־XP-V הפגם אינו ב־NER עצמו, אלא ב־Polymerase η, כלומר ב־TLS. במצב כזה אין סבילות יעילה לדימרים של תימין שלא תוקנו בזמן על ידי NER.
AID ו־MMR
ב־somatic hypermutation, חלק מהמוטציות שתלויות ב־AID אינן נמצאות בדיוק באתר הדה־אמינציה.
הסיבה היא שמנגנון MMR מסיר מקטע שלם סביב ה־mismatch, ואז סינתזה מחדש בעזרת פולימראזות פחות מדויקות, יכולה להכניס מוטציות גם בבסיסים סמוכים, בעיקר ב־A וב־T.
סוגי נזקי DNA ומנגנוני התיקון שלהם
בחלק הזה של הקורס למדנו נזקים בדרגות חומרה שונות: מנזק נקודתי בבסיס אחד, ועד שברים דו־גדיליים.
| סוג נזק | דוגמאות | מנגנון תיקון מרכזי |
|---|---|---|
| Base-level lesions | AP sites, deamination, oxidation | BER |
| Bulky lesions | T-dimers, platinum adducts | NER |
| Replication over damage | נזק שלא תוקן לפני שכפול | TLS |
| Mismatch | טעות בזיווג בסיסים אחרי שכפול | MMR |
| Single-strand breaks | Nick בגדיל אחד | SSBR/PARP1 |
| Double-strand breaks | שבר בשני הגדילים | NHEJ או HR |
הנקודה החשובה היא שלא מדובר באוסף מנגנונים מנותקים. זו רשת תיקון. מנגנון אחד יכול להיכשל, ואז מנגנון אחר נכנס לתמונה.
לדוגמה, אם נוצר T-dimer:
- המסלול הראשון שאמור לטפל בו הוא NER.
- אם NER לא הספיק לתקן לפני שכפול, התא יכול להשתמש ב־TLS כדי לעבור מעל הנזק.
- אם TLS הכניס טעות, MMR יכול לנסות לתקן אותה.
- אם במהלך NER או מנגנון אחר נוצר שבר חד־גדילי שלא תוקן, הוא יכול להפוך בזמן שכפול לשבר דו־גדילי.
- שבר דו־גדילי כבר ידרוש NHEJ או HR.
תבנית משותפת למנגנוני תיקון DNA
למרות שהמנגנונים שונים מאוד, כמעט כולם מתאימים לאותה תבנית כללית:
- Recognition - זיהוי הנזק.
- Processing - עיבוד האתר הפגוע והכנתו לתיקון.
- Repair - סינתזה מחדש וליגציה.
1. זיהוי (Recognition)
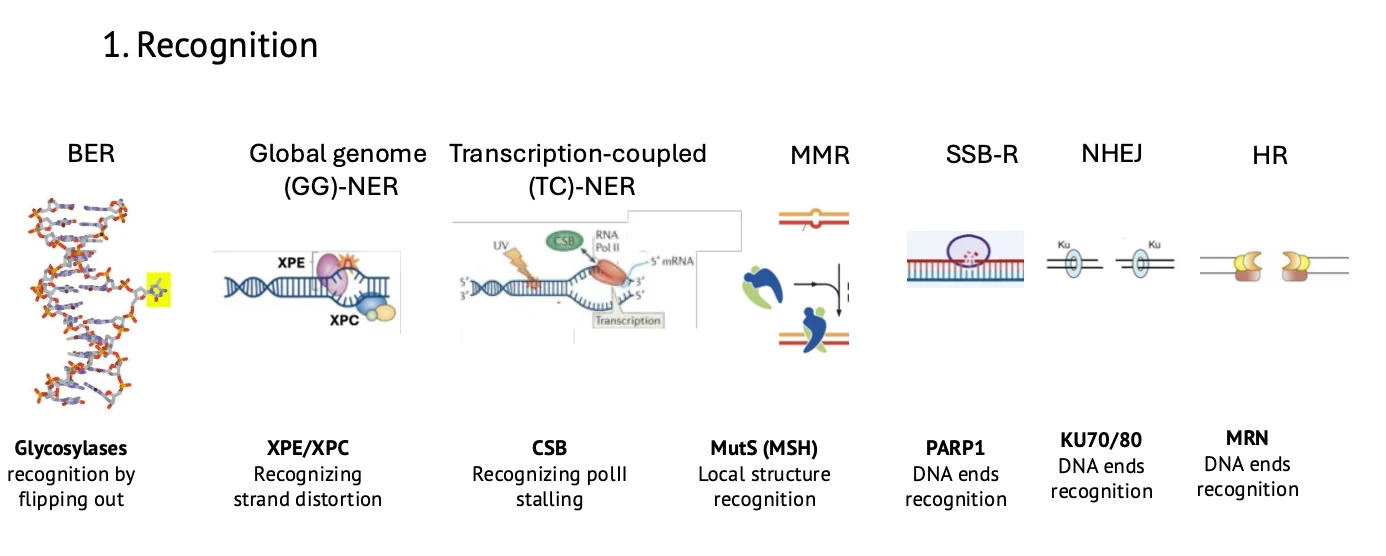
השלב הראשון הוא לזהות שיש בעיה. כל מנגנון מזהה משהו אחר.
| מנגנון | מה מזוהה? | חלבונים / עיקרון זיהוי |
|---|---|---|
| BER | בסיס פגום או בסיס לא מתאים | DNA glycosylases, למשל UNG או OGG1 |
| NER | עיוות במבנה ההליקס | XPC, XPE |
| Transcription-coupled NER | RNA polymerase שנתקע על נזק | CSB |
| MMR | mismatch אחרי שכפול | MutS/MSH |
| SSB repair | קצה חד־גדילי / nick | PARP1 |
| NHEJ | קצוות DNA דו־גדיליים | Ku70/Ku80 |
| HR | שבר דו־גדילי שצריך תיקון לפי תבנית | MRN complex |
ב־BER הזיהוי הוא כימי יחסית: גליקוזילאז מזהה בסיס פגום מסוים.
ב־NER הזיהוי הוא מבני יותר: החלבונים מזהים עיוות בדאבל הליקס.
ב־transcription-coupled NER, הזיהוי מתחיל מכך ש־RNA polymerase נתקע על הנזק.
ב־SSB, NHEJ ו־HR הזיהוי קשור לקצוות DNA שנוצרו בעקבות שבר.
2. עיבוד (Processing)
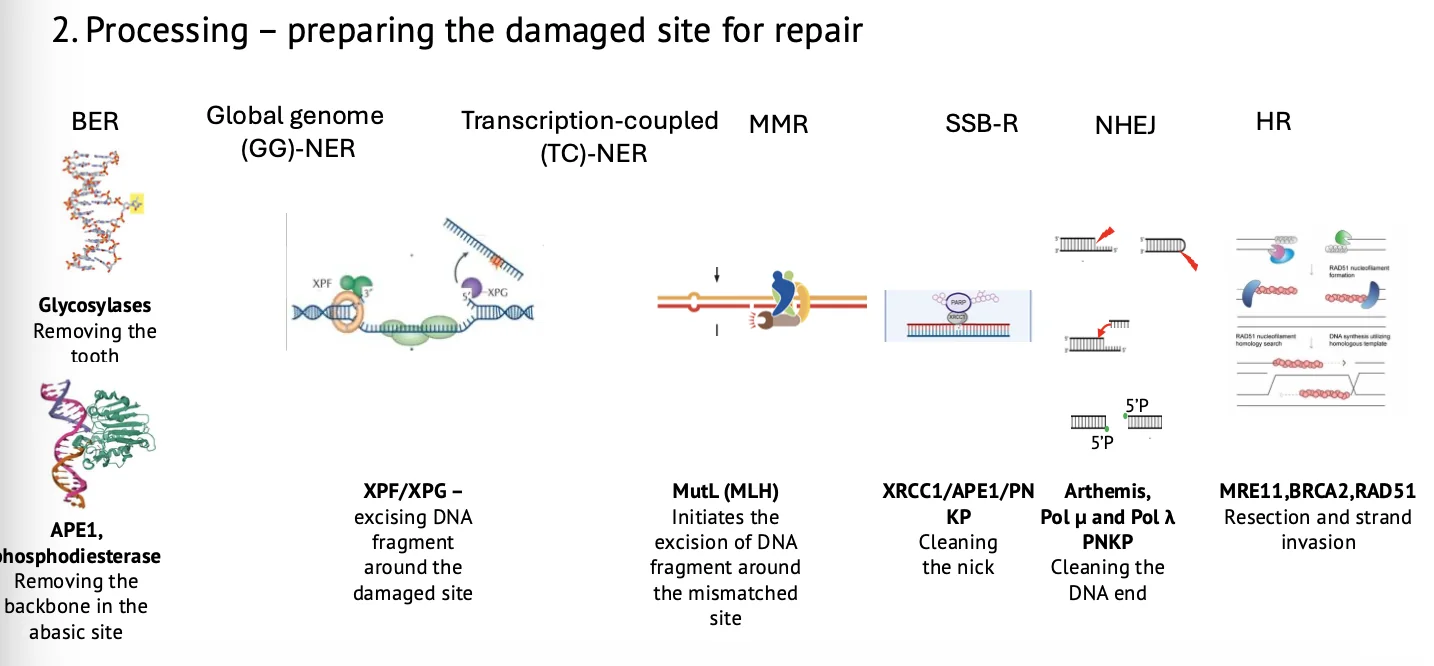
אחרי הזיהוי, צריך להכין את האתר לתיקון. הנזק בדרך כלל לא ״נקי״ מספיק כדי שפולימראז וליגאז יוכלו לעבוד מיד.
| מנגנון | Processing |
|---|---|
| BER | AP endonuclease ופוספודיאסטראז מנקים את האתר חסר־הבסיס |
| NER | XPF ו־XPG חותכים סביב הנזק ומסירים מקטע DNA |
| MMR | הסרת מקטע סביב ה־mismatch |
| SSB repair | XRCC1, APE1, PNKP ופקטורים נוספים מנקים את הקצוות |
| NHEJ | עיבוד קצוות: הסרת overhangs, פתיחת hairpins והכנת קצוות לליגציה |
| HR | end resection ליצירת גדיל חד־גדילי שיכול לחדור לכרומטידה האחות |
הבדל חשוב:
- ב־BER התיקון מאוד כירורגי, לרוב סביב בסיס אחד.
- ב־NER וב־MMR מסירים מקטע ארוך יותר.
- ב־NHEJ רוצים להכין קצוות לחיבור.
- ב־HR דווקא רוצים לבצע resection כדי להשתמש בכרומטידה האחות כתבנית.
3. תיקון (Repair)
בשלב האחרון יש סינתזה מחדש וסגירה של השרשרת.
הפולימראזות והליגאזות משתנות לפי סוג התיקון:
- Polymerase β מתאים לתיקון קצר וממוקד, כמו ב־BER.
- Polymerase δ/ε מתאימים לסינתזה של מקטעים ארוכים יותר.
- Ligase I/III סוגרים nick בגדיל אחד.
- Ligase IV מתאים ל־NHEJ, שבו צריך לחבר קצוות של שבר דו־גדילי.
תזמון לפי מחזור התא
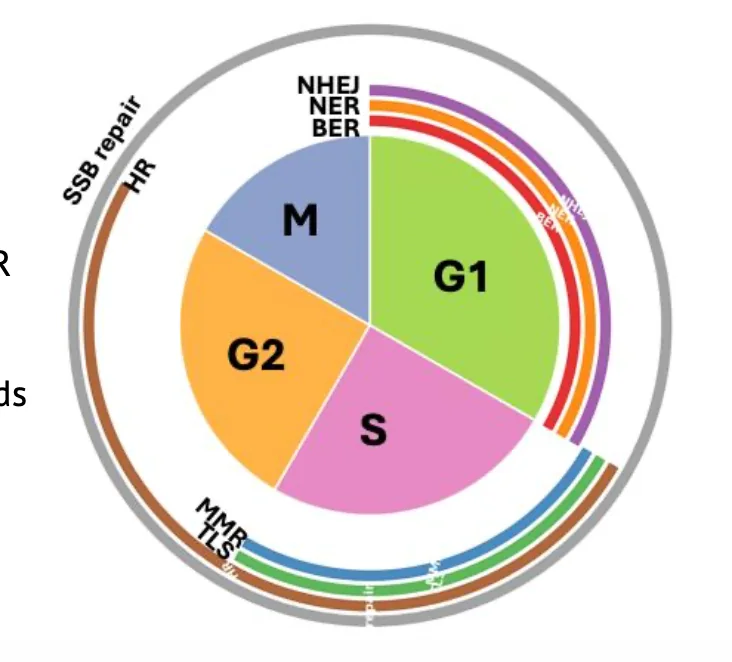
התא לא בוחר מנגנון תיקון רק לפי סוג הנזק. הוא גם תלוי במה שזמין לו באותו שלב של מחזור התא.
G1
ב־G1 אין כרומטידה אחות. לכן אם יש שבר דו־גדילי, התא לא יכול להשתמש ב־HR.
בשלב הזה בולטים:
- NHEJ
- BER
- NER
NHEJ מתאים במיוחד ל־G1 כי הוא לא דורש תבנית. הוא פשוט מחבר את הקצוות, גם אם זה פחות מדויק.
S phase
ב־S phase מתרחשת רפליקציה, ולכן מופיעים מנגנונים שקשורים ישירות לשכפול:
- MMR - תיקון טעויות שנוצרות בזמן שכפול.
- TLS - סבילות לנזק כאשר הפולימראז הרפליקטיבי נתקע.
- HR - כי יש כרומטידה אחות זמינה.
G2
ב־G2 השכפול כבר הושלם, ועדיין קיימת כרומטידה אחות. לכן HR עדיין אפשרי.
מיטוזה
אם התא כבר נכנס לחלוקה עצמה, תיקון שברים נעשה בעייתי יותר. לכן עצירת מחזור התא לפני מיטוזה חשובה מאוד כאשר יש נזק DNA משמעותי.
פתולוגיות וטיפולים שנגזרים ממנגנוני התיקון
הידע על מנגנוני תיקון DNA מסביר מחלות, סיכון לסרטן ובחירת טיפול.
| הקשר קליני | מנגנון רלוונטי | עיקרון |
|---|---|---|
| Skin cancer / XP | NER | פגיעה בתיקון דימרים של תימין וב־bulky lesions |
| XP-V | TLS / Polymerase η | פגיעה בסבילות לדימרים של תימין שלא תוקנו |
| Colorectal cancer MSI-high | MMR | עומס מוטציות ויצירת neoantigens |
| Breast / ovarian cancer עם BRCA deficiency | HR | פגיעה בתיקון שברים דו־גדיליים מדויק |
| B-cell leukemia | RAG / NHEJ / antigen receptor editing | חיתוכים לא רצויים ומחיקות גנומיות |
| עמידות ל־cisplatin | TLS | סבילות מוגברת ל־bulky adducts |
Colorectal cancer, MSI-high ואימונותרפיה
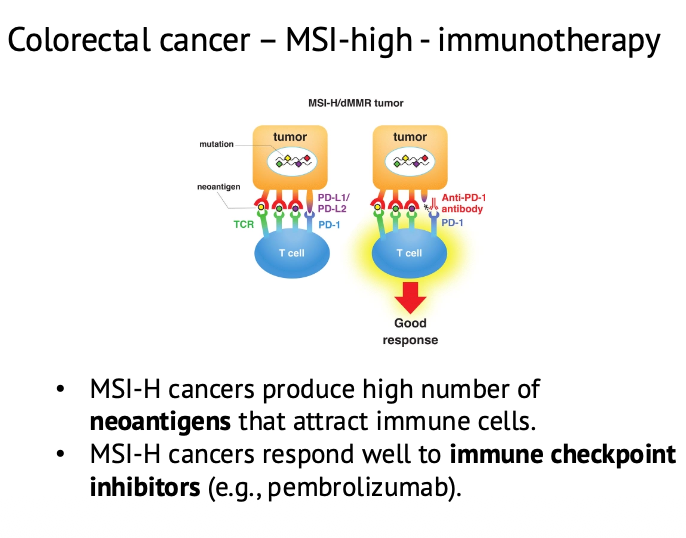
בגידולים עם MMR deficiency מצטברות טעויות שכפול. אחת התוצאות היא MSI-high, אבל המשמעות הרחבה יותר היא עומס מוטציות גבוה.
עומס מוטציות גבוה יכול ליצור neoantigens - חלבונים או פפטידים חדשים שהתא מציג, מערכת החיסון יכולה לזהות כלא־עצמיים.
לכן גידולי MSI-high, למשל בחלק ממקרי colorectal cancer, יכולים להגיב טוב יותר ל־immunotherapy.
אחד הכיוונים הוא checkpoint inhibitors. תאים סרטניים יכולים לבטא מולקולות שמעכבות את מערכת החיסון, כמו PD-1/PD-L1 axis. טיפול שמעכב את המחסום הזה מאפשר למערכת החיסון לפעול טוב יותר נגד התא הסרטני.
BRCA-deficient tumors ומעכבי PARP
בגידולים עם BRCA deficiency יש פגיעה ב־HR. כלומר, התא מתקשה לתקן שברים דו־גדיליים בצורה מדויקת.
מעכבי PARP יוצרים מצב שבו שברים חד־גדיליים אינם מתוקנים היטב, וחלקם הופכים בזמן שכפול לשברים דו־גדיליים. תא עם HR תקין יכול להתמודד עם חלק מהשברים האלה. תא עם BRCA deficiency לא מצליח.
זהו העיקרון של: Synthetic lethality.
שתי פגיעות יחד יוצרות קטלניות:
- פגיעה קיימת ב־HR בגלל BRCA deficiency.
- עיכוב PARP כטיפול.
התוצאה היא הצטברות נזקים ודחיפה של התא הסרטני למוות תאי.
TLS, סבילות ועמידות לטיפול
TLS יושב על הגבול בין הצלה לבין סכנה.
מצד אחד, הוא מאפשר לתא להמשיך שכפול כאשר יש נזק. זה מונע קריסה של מזלג השכפול ומפחית אפופטוזיס.
מצד שני, TLS עלול להיות error-prone. לכן פעילות מוגברת שלו יכולה לגרום ל:
- יותר מוטציות.
- סבילות מוגברת לנזקי DNA.
- עמידות לטיפולים אנטי-סרטניים שיוצרים bulky adducts, למשל cisplatin.
Polymerase η הוא יחסית high fidelity בהקשר של T-dimers, כי זו ההתמחות שלו. לעומתו, פולימראזות TLS אחרות כמו Pol κ או Pol ι יכולות להיות יותר error-prone.
נזקי DNA יזומים - Damage for the best
לא כל נזק DNA הוא תקלה. יש תהליכים פיזיולוגיים שבהם התא יוצר נזק בכוונה כדי להשיג תוצאה ביולוגית חשובה.
הדוגמאות המרכזיות מהקורס:
- V(D)J recombination - יצירת מגוון של רצפטורים לאנטיגן.
- Somatic hypermutation - יצירת מוטציות בנוגדנים כדי לשפר את האפיניות שלהם לאנטיגן.
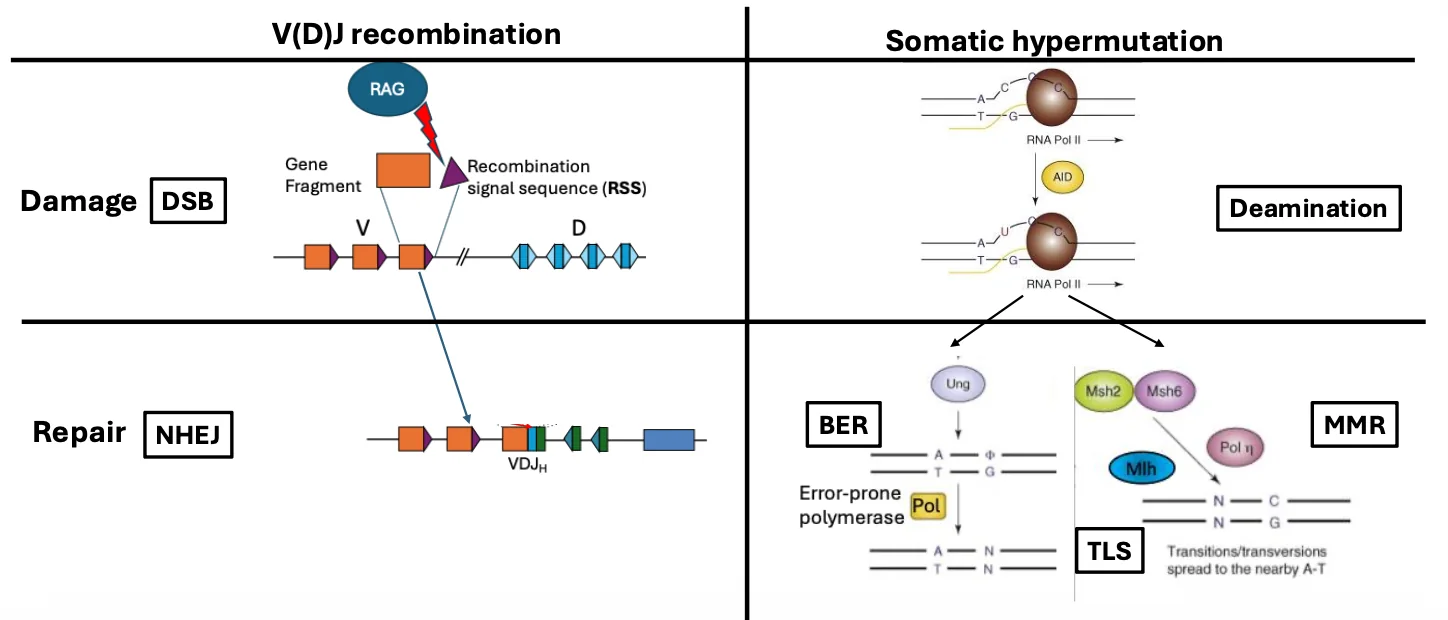
V(D)J recombination
ב־V(D)J recombination נוצרים שברים דו־גדיליים בלוקוסים שמקודדים לנוגדנים ול־T-cell receptors.
- החיתוך נעשה על ידי RAG.
- החיבור מחדש נעשה על ידי NHEJ.
- המטרה היא ליצור מגוון גדול של רצפטורים לאנטיגן.
זה תהליך של cut and paste ברמת ה־DNA.
בטבלה שבתמונה למעלה, V(D)J recombination שייך לעולם של DSB ושל NHEJ, כי יש יצירת שברים דו־גדיליים וחיבור מחדש של קצוות.
Somatic hypermutation
ב־somatic hypermutation הנזק הוא אחר: לא שבר דו־גדילי, אלא דה־אמינציה של ציטוזין.
- האנזים המרכזי הוא AID.
- AID עושה: $\ce{C -> U}$
- התיקון או השכפול של הנזק הזה יוצר מוטציות נקודתיות.
- המטרה היא לשפר אפיניות של נוגדנים לאנטיגן.
כאן התא מנצל מנגנוני תיקון כמו BER ו־MMR, אבל במקום תיקון מדויק, הם מגויסים ליצירת גיוון.
למה זה מסוכן?
אותם מנגנונים שמייצרים גיוון חיסוני יכולים לגרום גם לנזק לא רצוי.
- RAG יכול לזהות cryptic RSS וליצור מחיקות או טרנסלוקציות.
- AID יכול לגרום לדה־אמינציה גם מחוץ לאזורי האימונוגלובולינים.
- תיקון לא מדויק יכול לייצר מוטציות שמקדמות סרטן.
העיקרון הוא שהגוף משתמש בנזקי DNA ככלי, אבל הכלי הזה דורש בקרה חזקה מאוד.
ספריות DNA ככלי למיפוי נזקי DNA
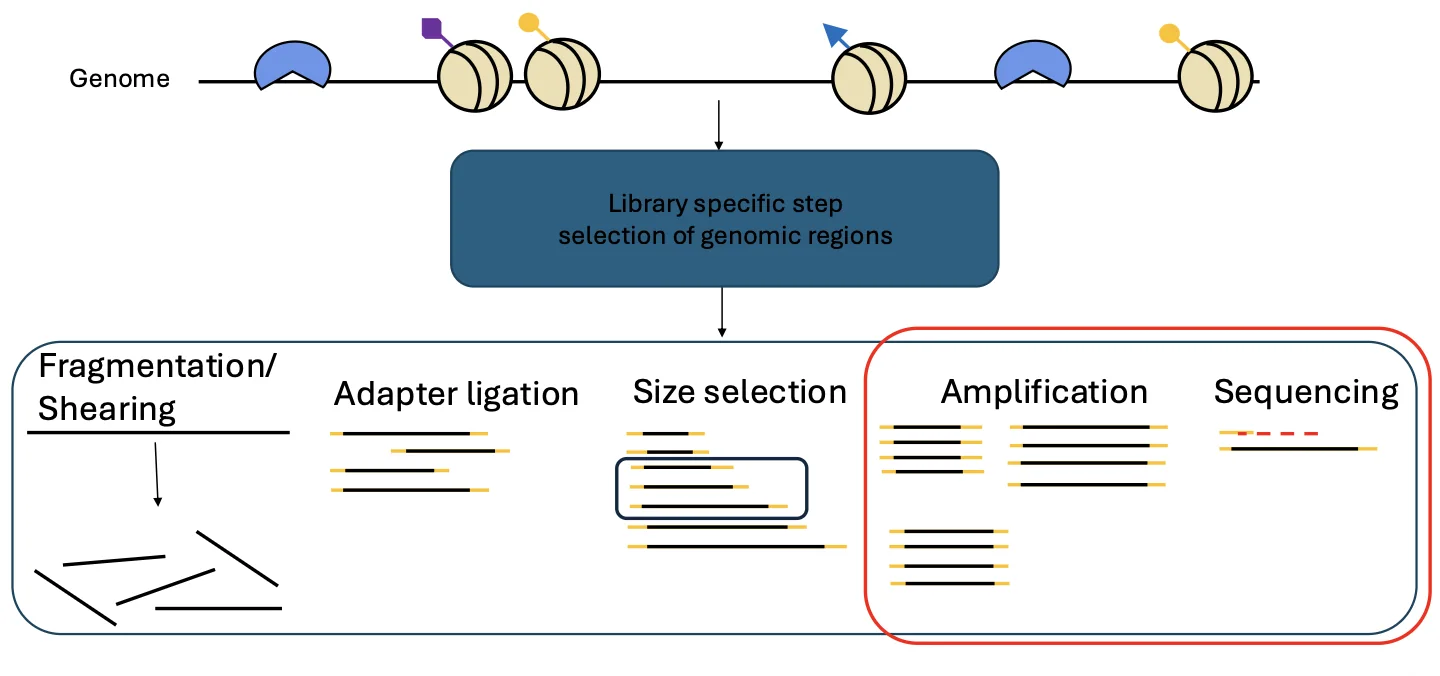
בסוף השיעור חזרנו לתחילת הקורס: הכנת ספריות DNA.
המטרה הכללית של ספריית DNA היא ליצור מקטעי DNA לא ידועים עם אדפטורים ידועים, כדי שאפשר יהיה לרצף אותם במכונת NGS.
השלבים הכלליים:
- Fragmentation
- Adapter ligation
- Size selection
- Amplification
- Sequencing
אבל לפני השלבים הכלליים האלה יש שלב חשוב יותר: סלקציה.
השאלה היא איזה חלק מהגנום רוצים לרצף. בקורס פגשנו כמה דוגמאות שבהן הסלקציה מבוססת על סוג הנזק.
מיפוי AP sites
כדי למפות אתרים חסרי בסיס משתמשים במולקולה שמזהה AP sites ומחוברת ל־biotin.
לאחר מכן משתמשים ב־streptavidin beads כדי לדוג את המקטעים שסומנו.
הרעיון:
- AP site מזוהה על ידי מולקולה מתאימה.
- המולקולה מחוברת לביוטין.
- streptavidin על ביד מגנטי קושר את הביוטין.
- דגים את המקטעים.
- מרצפים וממפים לגנום.
אם רוצים למפות 8-oxoG, משתמשים קודם ב־OGG1 כדי להפוך את 8-oxoG ל־AP site, ואז משתמשים באותה אסטרטגיית AP-seq.
מיפוי T-dimers
במיפוי דימרים של תימין משתמשים באנזים שיודע לזהות את הנזק ולחתוך לידו, למשל T4 endonuclease.
החיתוך יוצר קצה שאפשר לחבר אליו אדפטור. כך תחילת הקריאה יכולה לסמן את מיקום הנזק.
העיקרון כאן שונה מ־AP sites: במקום לדוג כימית אתר חסר בסיס, משתמשים באנזים שמזהה וחותך באתר הנזק.
מיפוי שברי DNA
במיפוי שברים דו־גדיליים, למשל ב־END-seq, מנצלים את העובדה שכבר יש קצה טבעי של DNA.
בספריות רגילות יוצרים קצוות בעזרת סוניקציה. כאן רוצים קודם לתפוס את הקצוות שנוצרו ביולוגית.
לכן:
- שומרים את התאים / ה־DNA באגרוז כדי למנוע שברים מכניים.
- מחברים אדפטור לקצה הטבעי של השבר.
- מסמנים את האדפטור, למשל בעזרת ביוטין.
- דגים את המקטעים הרלוונטיים.
- רק אחר כך מבצעים שלבים נוספים של הכנת הספרייה.
האתגר הוא למנוע שברי רקע במהלך ההכנה, כי כל שבר מלאכותי יכול להיראות כמו שבר ביולוגי.
מה משותף לכל הספריות האלה?
בכל המקרים משתמשים באותו רעיון בסיסי:
- יש נזק מסוים.
- מוצאים דרך לזהות אותו או להפוך אותו לקצה שניתן לריצוף.
- עושים סלקציה למקטעים הרלוונטיים.
- מחברים אדפטורים.
- מרצפים.
- ממפים את הקריאות לגנום.
כלומר, NGS אינו רק כלי לריצוף גנום. הוא הופך לשיטה למפות איפה נזקים נוצרים, איזה אזורים רגישים יותר, ואיך טיפולים או מוטציות משנים את דפוסי הנזק.
דור פסקל