פתיחה - משכפול לשעתוק
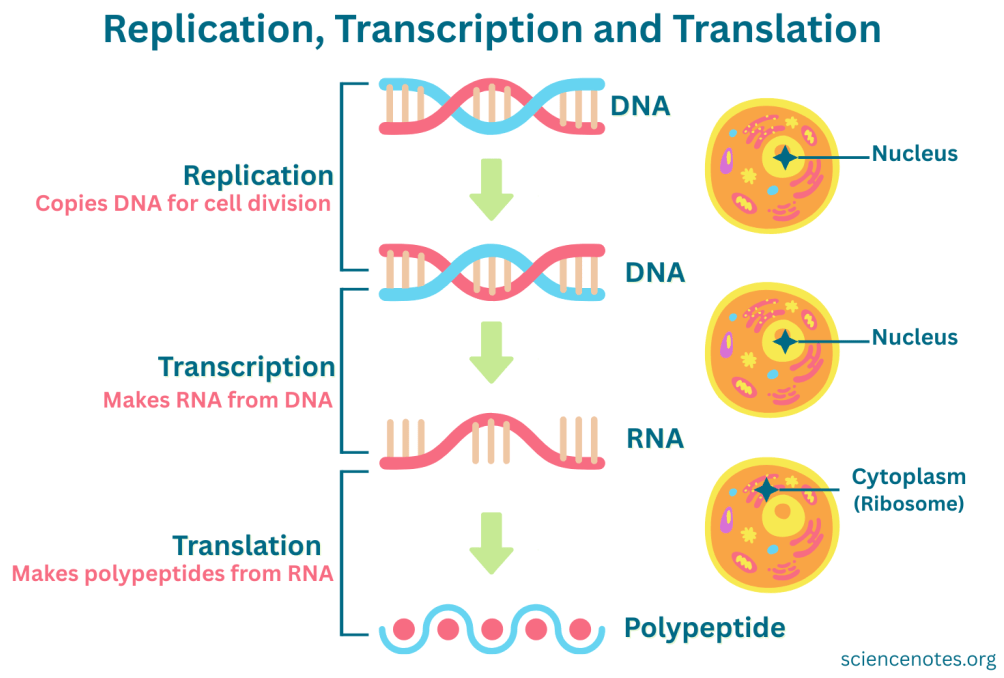
הזרימה הכללית של מידע גנטי היא:
DNA → RNA → Protein
- Replication - שכפול DNA ל־DNA.
- Transcription - שעתוק DNA ל־RNA.
- Translation - תרגום RNA לחלבון.
שכפול DNA מתרחש ב־S phase (שלב הסינתזה) ומטרתו להעביר עותק שלם של החומר הגנטי לתאי הבת. לעומת זאת, שעתוק מעתיק בכל פעם גנים מסוימים לפי צורכי התא.
עיקר השעתוק והתרגום מתרחשים ב־G1, אך הם מתקיימים גם בשלבים נוספים:
- ב־S phase משועתקים ומתורגמים, בין היתר, היסטונים הדרושים לאריזת ה־DNA החדש.
- ב־G2 נוצרים גם חלבונים המשתתפים בהכנה לחלוקה ובבקרת מחזור התא.
לכן שעתוק ותרגום אינם שייכים לשלב יחיד, אלא משתנים לפי סוג התא, הגן ומצב התא.
תזכורת קצרה: שלבי שכפול DNA
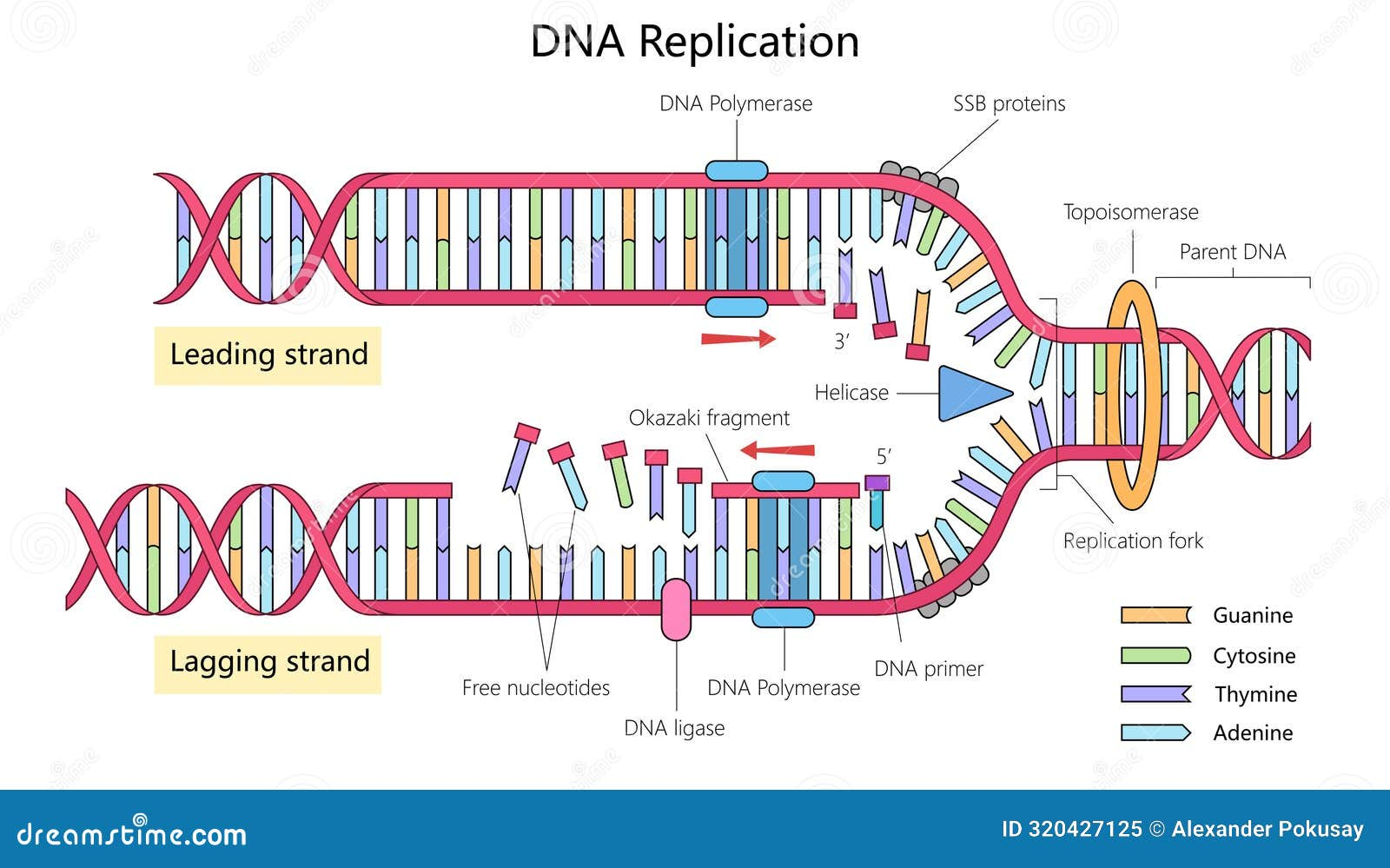
- Initiation - ה־Helicase פותח את ה־DNA הדו־גדילי.
- Priming - ה־Primase מייצר RNA primers שמספקים קצה התחלתי ל־DNA polymerase.
- Elongation - ה־DNA polymerase מוסיף נוקלאוטידים משלימים ומסנתז גדילים חדשים.
- Termination - מנגנון השכפול מתפרק לאחר השלמת ההעתקה; מנגנוני proofreading ותיקון מפחיתים טעויות.
בהמשך יחידת הלימוד נעסוק בשעתוק בחיידקים ובתאים אאוקריוטיים, עיבוד RNA, בקרה על שעתוק, תרגום וסוגי RNA שאינם מקודדים לחלבון.
DNA לעומת RNA
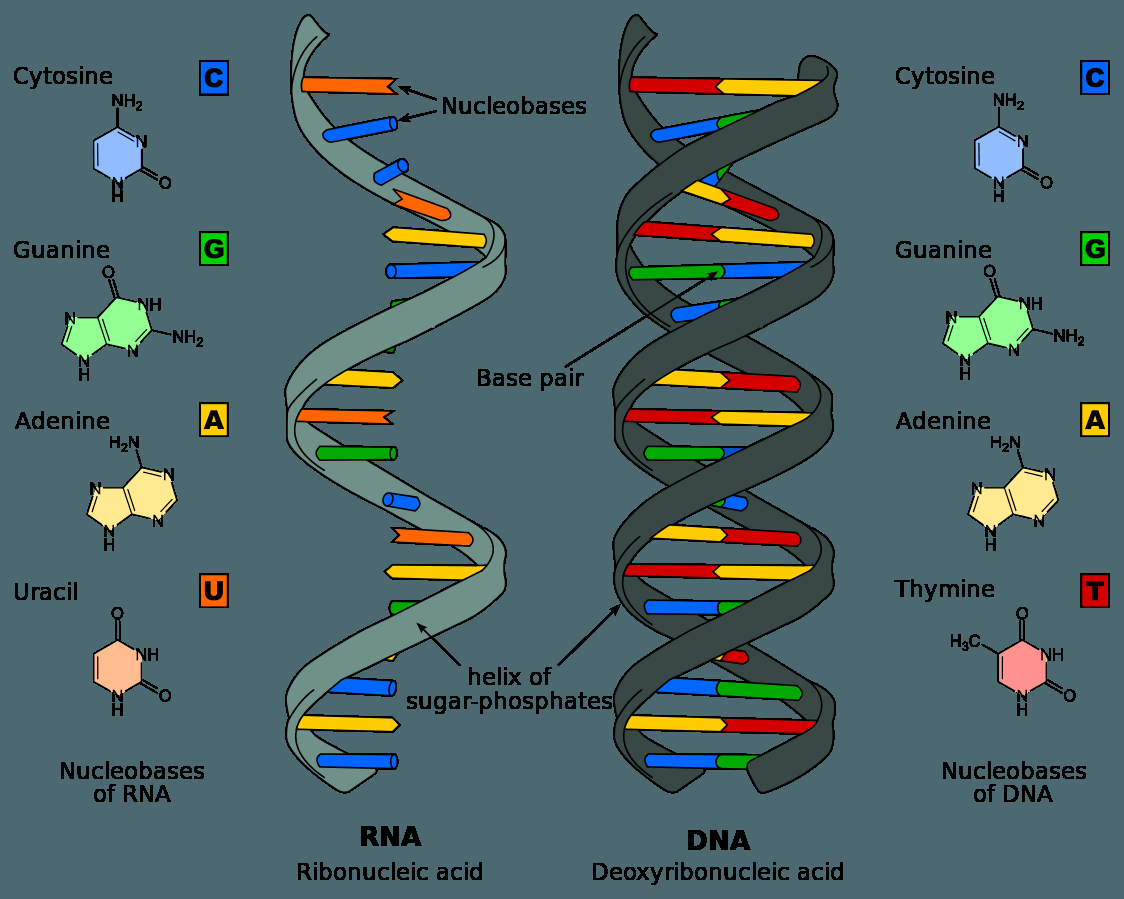
תזכורת למבנה ה־DNA
ה־DNA בנוי משני גדילים אנטי-מקבילים (Anti-parallel) היוצרים מבנה של סליל כפול (Double helix).
כל נוקלאוטיד מורכב מ:
- סוכר דאוקסיריבוז (Deoxyribose), אשר בעמדה מספר 2 שלו יש אטום מימן (H).
- אחד מארבעת הבסיסים החנקניים: אדנין (A), גואנין (G), ציטוזין (C) או תימין (T).
| מאפיין | DNA | RNA |
|---|---|---|
| מספר גדילים | דו־גדילי (יוצר סליל כפול) | חד־גדילי |
| מבנה מרחבי | סליל כפול (Double helix) | מתקפל במרחב למבנים תלת־ממדיים שונים |
| סוכר | דאוקסיריבוז (בעל H בעמדה 2) | ריבוז (בעל OH בעמדה 2) |
| בסיסים | A, T, C, G | A, U, C, G |
| יציבות | יציב מאוד | פחות יציב, כדי לאפשר לתא לשלוט בו |
המבנה החד־גדילי ויציבות ה־RNA
בניגוד ל־DNA שהוא דו־גדילי, ה־RNA הוא חד־גדילי, ולכן הוא מסתדר במרחב בכל מיני צורות תלת־ממדיות. המבנים הללו עוזרים ליציבות של ה־RNA או לחוסר היציבות שלו, בהתאם למה שהתא צריך: אם צריך שה־RNA יהיה יציב, הוא ייעזר במבנים האלה; ואם התא צריך לפרק אותו, הוא יישאר פחות יציב.
בנוסף, ב־RNA נמצא הבסיס Uracil (U) במקום Thymine (T). הבסיס T הוא הרבה יותר יציב מהבסיס U. מכיוון שהתא רוצה לשלוט ב־RNA והוא צריך להיות פחות יציב, יש בו U (שיכול גם לעבור כל מיני מודיפיקציות) כדי להקל על הבקרה עליו.
עקרון השעתוק
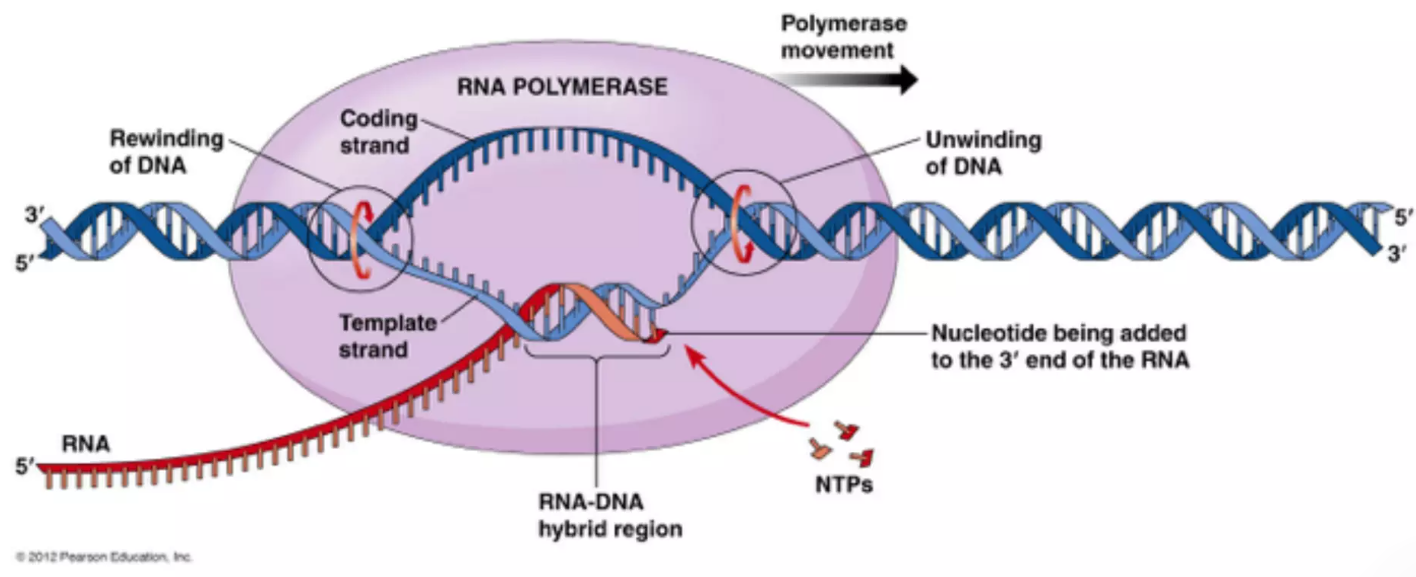 Transcription (שעתוק) הוא תהליך שבו RNA polymerase מגיע לאזור שלפני הגן, פותח את שני גדילי ה־DNA ומתחיל לשעתק את הגן.
Transcription (שעתוק) הוא תהליך שבו RNA polymerase מגיע לאזור שלפני הגן, פותח את שני גדילי ה־DNA ומתחיל לשעתק את הגן. במהלך התהליך הפולימראז מסנתז RNA חדש תמיד בכיוון אחד בלבד: מ־5'→3'.
גדיל התבנית והגדיל המקודד
- הגדיל של ה־DNA שממנו נוצר ה־RNA נקרא Template strand או Antisense strand.
- הגדיל השני וההופכי לו נקרא Coding strand או Sense strand.
ה־RNA שנוצר משלים בבסיסיו לגדיל התבנית. כתוצאה מכך, הרצף של ה־RNA החדש כמעט זהה לחלוטין לזה של ה־Coding strand, כאשר ההבדל היחיד הוא שב־RNA מופיע הבסיס U במקום T.
כאשר משעתקים (ומקודדים) RNA מתוך גדיל התבנית:
- מול הבסיס A מציבים U
- מול הבסיס T מציבים A
- מול הבסיס C מציבים G
- מול הבסיס G מציבים C
RNA polymerase
RNA polymerase הוא האנזים שמסנתז את ה־RNA לפי גדיל ה־template של ה־DNA. כיאה לאנזים, יש לו אזור קטליטי בתוך המבנה החלבוני, שאחראי על סינתזת השרשרת.
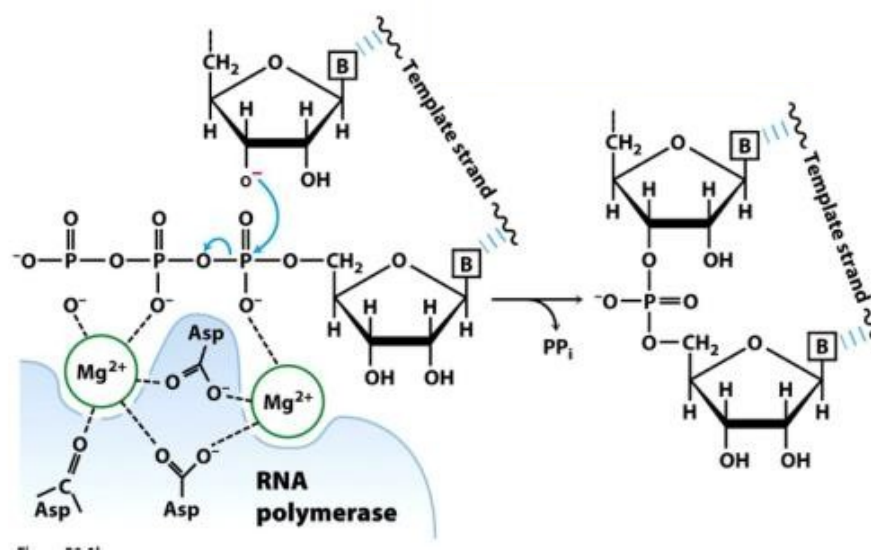
תהליך הקטליזה ויצירת הקשר
- יצירת הקשר: האנזים מוריד דו־פוספט (פירופוספט) מהנוקלאוטיד הנכנס (השני), ומחבר אותו לנוקלאוטיד הראשון בשרשרת באמצעות קשר פוספודיאסטרי.
- האתר הפעיל: הפעולה הקטליטית מתאפשרת הודות לשלוש חומצות אמינו של אספרגין (אספרטט) הנמצאות באתר הפעיל, אשר פועלות יחד עם שני יוני מגנזיום ($\ce{Mg^{2+}}$).
- אנרגיה: הורדת הפוספטים משחררת את האנרגיה הדרושה ליצירת הקשר הפוספודיאסטרי החדש.
- הקשר הכימי נוצר ספציפית בין קבוצת ההידרוקסיל של הנוקלאוטיד הראשון לבין קבוצת הפוספט של הנוקלאוטיד השני.
כיווניות השרשרת (5’ ל־3’)
- השרשרת תמיד מסונתזת בכיווניות של 5’ ל־3’.
- הנוקלאוטיד הראשון בשרשרת מאופיין בכך שהעמדה הפנויה שלו היא פחמן מספר 5’.
- הנוקלאוטיד האחרון בשרשרת מאופיין בכך שהעמדה הפנויה שלו היא פחמן מספר 3’.
- כתוצאה מחיבורים אלו, נוצר שלד (Backbone) אחיד של ה־RNA, כאשר כל הבסיסים פונים כלפי צד אחד.
RNA polymerase בחיידקים לעומת תאים אאוקריוטיים
בחיידקים יש סוג עיקרי אחד של RNA polymerase, המסנתז את כל קבוצות ה־RNA, ובהן mRNA, rRNA, tRNA ו־RNA רגולטוריים קטנים.
בתאים אאוקריוטיים קיימים שלושה RNA polymerases גרעיניים עיקריים:
| פולימראז | תוצרים עיקריים |
|---|---|
| RNA polymerase I | רוב ה־rRNA: 28S, 18S ו־5.8S; פעילותו העיקרית ב־nucleolus |
| RNA polymerase II | כל ה־mRNA המקודד לחלבון, וכן חלק מ־snRNA, miRNA, siRNA, snoRNA ו־lncRNA |
| RNA polymerase III | tRNA, 5S rRNA וחלק מה־RNA הקטנים |
RNA polymerase II האאוקריוטי דומה מבחינה מבנית לפולימראז החיידקי, אך התחלת השעתוק באאוקריוטים מורכבת יותר:
- בחיידקים נדרש בעיקר פקטור התחלה אחד, sigma factor.
- באאוקריוטים נדרשים כמה general transcription factors.
- באאוקריוטים ה־DNA ארוז בנוקלאוזומים ובכרומטין, ולכן יש להתגבר גם על אריזת ה־DNA.
שלבי השעתוק
בכל מערכות השעתוק יש שלושה שלבים כלליים:
- אתחול (Initiation) - זיהוי הפרומוטור והתחלת סינתזת RNA.
- התארכות (Elongation) - הארכת שרשרת ה־RNA.
- סיום/טרמינציה (Termination) - עצירת השעתוק ושחרור התעתיק.
שעתוק בחיידקים
Core enzyme ו־Holoenzyme
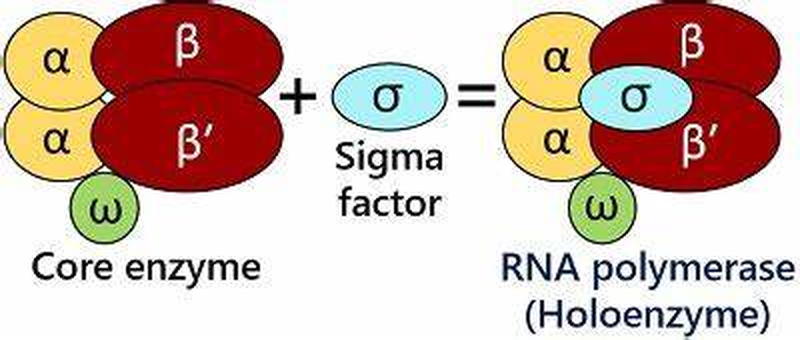
בחיידקים ישנו סוג אחד בלבד של RNA polymerase המבצע את כל תהליכי השעתוק.
האנזים הפעיל והשלם נקרא Holoenzyme, והוא נוצר מחיבור של שני מרכיבים:
-
Core enzyme (אנזים הליבה): מורכב ממספר תת־יחידות ($\mathrm{2\alpha}$, $\mathrm{\beta}$, $\mathrm{\beta’}$, ו־$\mathrm{\omega}$). ה־Core enzyme הוא מבנה קבוע שזהה בכל תהליכי השעתוק ואינו ספציפי לגן מסוים.
-
Sigma factor (פקטור $\sigma$): תת־יחידה נוספת המעניקה לפולימראז את הספציפיות שלו. קיימים סוגים שונים של פקטורי סיגמא בתא, וכל אחד מהם יודע לזהות ולהיקשר לגנים שונים בהתאם לצורכי החיידק.
שלבי השעתוק של RNA polymerase בחיידקים
ה־Holoenzyme נקשר ל־DNA ונע על גביו (מתחבר ומתנתק באופן קבוע) עד שהוא מזהה את אזור תחילת הגן. תהליך השעתוק עצמו מחולק לשלושה שלבים עיקריים:
1. איתחול (Initiation)
הפרומוטור (Promoter)
הפרומוטור הוא האזור על ה־DNA שנמצא לפני הגן (Upstream), אליו נקשר ה־Sigma factor ומביא איתו את כל הפולימראז. נקודת תחילת הגן מוגדרת כ־+1, והאזורים שלפניו בפרומוטור נספרים במינוסים.
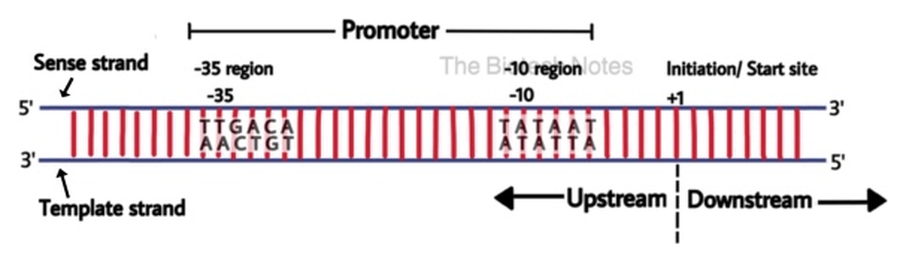
הפרומוטור הידוע והנחקר ביותר בחיידקים נקרא Pribnow box, והוא כולל שני אזורים מרכזיים:
- אזור ה־
-10: אזור המכיל רצף עשיר ב־T ו־A. מכיוון שזוגות T-A יוצרים ביניהם רק שני קשרי מימן (לעומת שלושה ב־C-G), קל יותר להפריד את הגדילים באזור זה - פעולה הכרחית לתחילת השעתוק. - אזור ה־
-35: אזור זיהוי נוסף לפקטור הסיגמא.
חוזק הפרומוטור תלוי ברצף הנוקלאוטידים שלו; שינויים ברצף (למשל יותר C ו־G) ישפיעו על חוזק הקישור של ה־Sigma factor לאזור.
Scrunching mechanism (מנגנון המשיכה)
בשלב זה, ה־RNA פולימראז כבר קשור לפרומוטור בעזרת ה־Sigma factor, ולכן הוא מקובע ואינו יכול להתקדם.
- כאנזים, הפולימראז מנסה להתחיל לשעתק, ומושך את ה־DNA לתוך עצמו (כמו חבל), אך נשאר תקוע במקומו. המרצה דימתה זאת למכונית מרוץ שלוחצת במקביל על הגז ועל הברקס.
- תהליך זה של משיכה, שחרור ומשיכה חוזרת נקרא Scrunching.
- ה”מאבק” הזה יוצר הצטברות גדולה מאוד של אנרגיה.
- רק כאשר נצברת מספיק אנרגיה והפולימראז מצליח לסנתז מקטע קצר של כ־10 נוקלאוטידים של RNA חדש, הוא “משתחרר” מהפרומוטור.
- ברגע השחרור, ה־Sigma factor מתנתק מהקומפלקס ויוצא לסירקולציה, והפולימראז ממשיך לבדו לשלב הבא.
2. התארכות (Elongation)
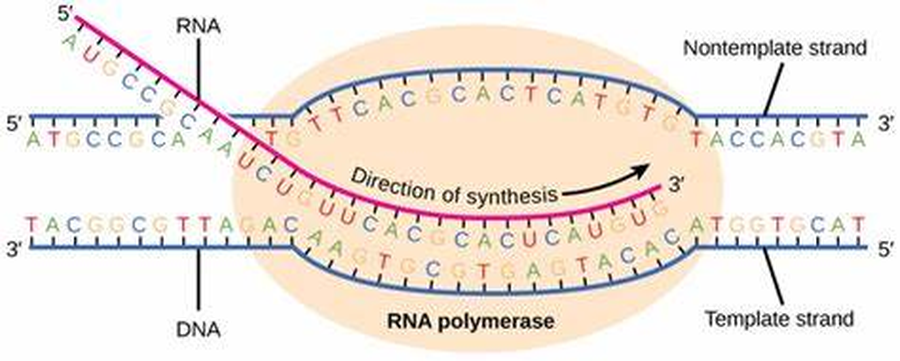
לאחר שהשתחרר מהפרומוטור, ה־RNA פולימראז מתקדם לאורך גדיל התבנית (ה־Template/Antisense strand).
הוא פותח את גדילי ה־DNA, מסנתז באופן רציף את ה־RNA החדש בכיווניות של 5'→3', וממשיך לנוע עד שהוא מגיע לסוף הגן - שם יתבצע שלב הטרמינציה (סיום השעתוק).
3. טרמינציה
בחיידקים יש שתי דרכים עיקריות לסיים שעתוק:
- Rho-independent termination, הנקרא גם intrinsic termination.
- Rho-dependent termination.
Rho-independent termination
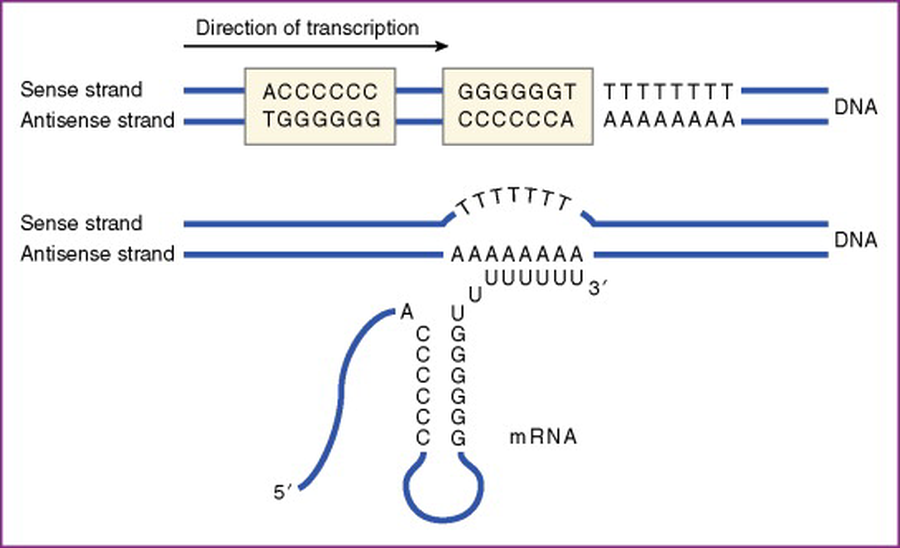
במסלול זה, סיום השעתוק אינו תלוי בחלבון Rho אלא ברצף הגן עצמו. בסוף הגן, על גבי ה־DNA (גדיל התבנית), יש רצפים של poly(G) ולאחריהם poly(C), שמיד אחריהם גם מגיע רצף ארוך של poly(A).
לאחר השעתוק מתקבלים ב־RNA:
- שני רצפים משלימים (poly(C) ו־poly(G)) שעושים אינטראקציה ומתקפלים למבנה של hairpin (סיכת ראש).
- רצף ארוך של poly(U) מיד לאחר הלופ (שנוצר מול רצף ה־poly(A) שבדנ”א).
ה־hairpin יוצר הפרעה סטרית, ובמקביל, הקשרים שנוצרים בין הנוקלאוטידים של ה־U ב־RNA, ל־A ב־DNA, הם קשרים חלשים יחסית. שילוב שני הגורמים הללו מאפשר את שחרור ה־RNA מה־DNA, ואת ניתוק הפולימראז מהקומפלקס.
Rho-dependent termination
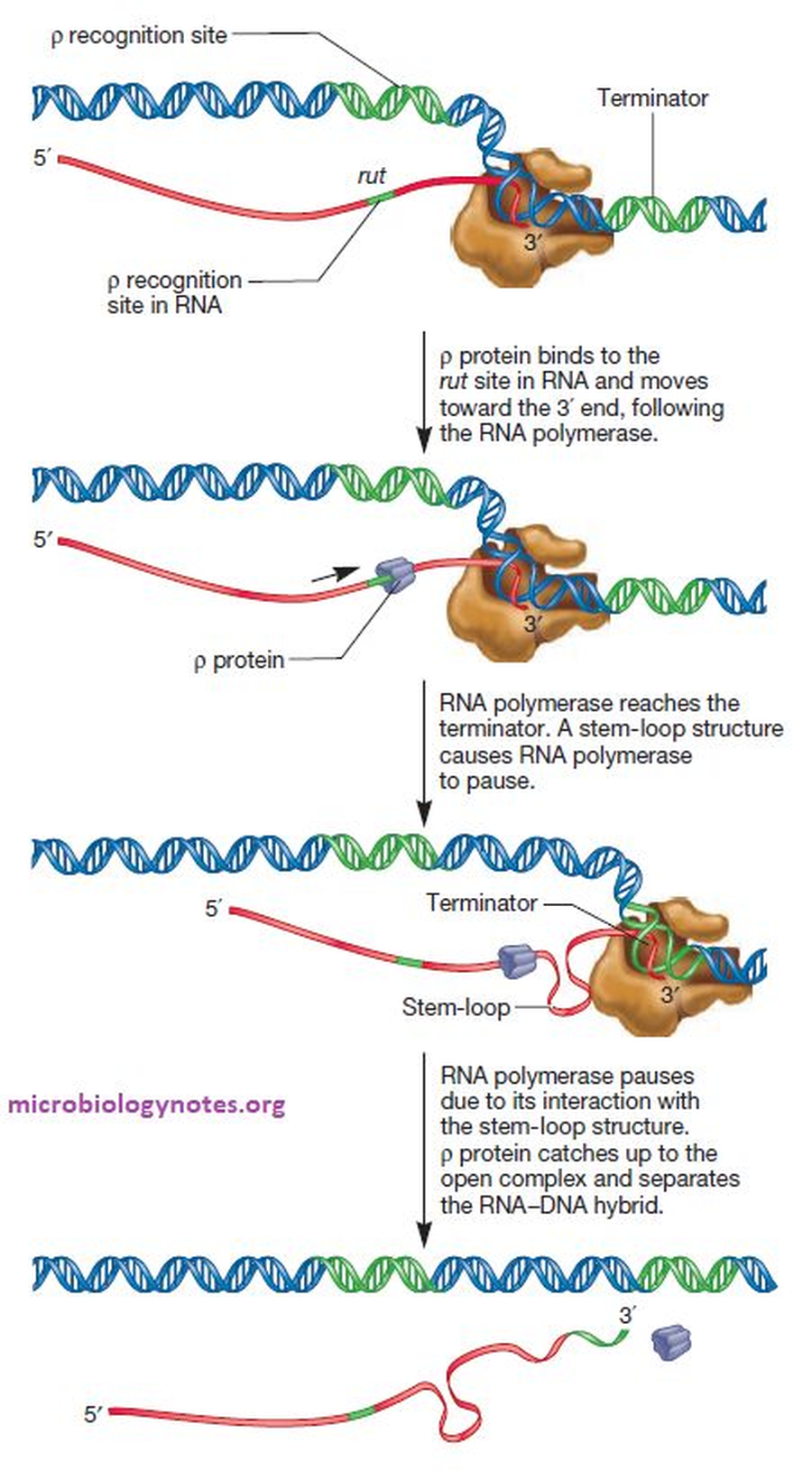
במסלול זה נדרשת פעילות של חלבון ה־Rho.
- RNA polymerase משעתק רצף בשם rut (Rho utilization site), שמצוי על הגן.
- חלבון Rho מזהה את הרצף הזה על גבי ה־RNA החדש, נקשר אליו, ומתחיל לנוע לאורכו ולרדוף אחרי ה־RNA polymerase.
- הפולימראז ממשיך לסנתז, עד שהוא מגיע לאזור שגורם ל־RNA ליצור stem-loop (לופ קטן יותר מה־hairpin במסלול הראשון). מבנה זה גורם לפולימראז להאט.
- ההאטה מאפשרת לחלבון Rho להשיג את הפולימראז.
- חלבון Rho נכנס ממש אל האזור הקטליטי (שבו ה־DNA פתוח ומסונתז ה־RNA), מפריד פיזית את ה־RNA מה־DNA, ומפרק את קומפלקס השעתוק.
| סוג טרמינציה | הגורם המסיים | עיקרון |
|---|---|---|
| Rho-independent | מבנה Hairpin ורצף poly(U) | ההפרעה הסטרית של הלופ יחד עם קשרי A-U חלשים מנתקים את הקומפלקס |
| Rho-dependent | חלבון Rho | Rho נקשר ל־rut, משיג פולימראז שהאט בגלל ה־stem-loop, נכנס לאזור הקטליטי ומפרק את הקומפלקס |
ברגע שתהליך השעתוק נגמר, ה־RNA פולימראז משתחרר, פוגש sigma factor (אותו אחד או אחר), והם יכולים ליצור אינטראקציה מחדש ולעלות שוב על ה־DNA, כדי להתחיל מחזור שעתוק נוסף.
אופרונים בחיידקים
אופרון (Operon) מאפשר לחיידק לייעל תהליכים, ולבקר יחד מספר גנים בעלי תפקיד משותף.
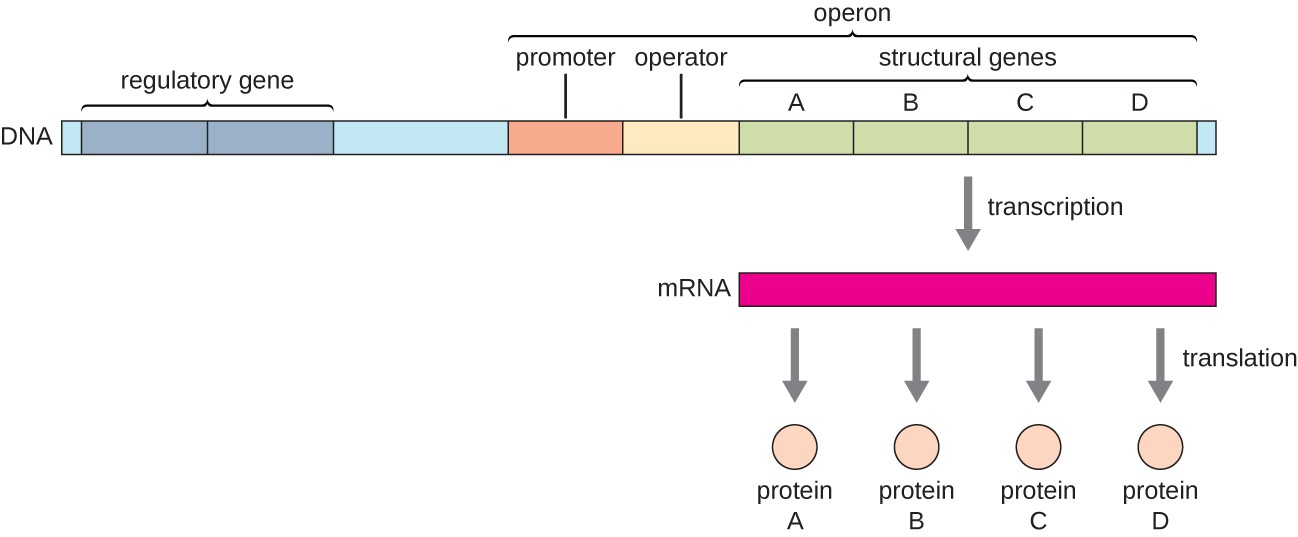
בדרך כלל, אופרון כולל:
- Promoter (פרומוטור) - האזור שאליו מגויס ה־RNA פולימראז.
- Operator (אופרטור) - אזור רגולטורי (לרוב חלק מהפרומוטור) אליו נקשר הרפרסור.
- גנים - מספר גנים הקשורים לאותו תהליך ומסונתזים יחד.
- גן רגולטורי - גן נוסף (שאינו חלק מהאופרון עצמו) המייצר חלבון בקרה שנקרא Repressor (רפרסור).
שני סוגי אופרונים:
- אינדוסיביליים - צריכים גירוי (סטימולוס) כדי לפעול. בלי הגירוי האופרון כבוי.
- רפרסביליים - הגירוי מונע מהם לפעול, קרי, פועלים עד שהגירוי מכבה אותם.
Inducible operon (אופרון אינדוסיבילי) - אופרון הלקטוז
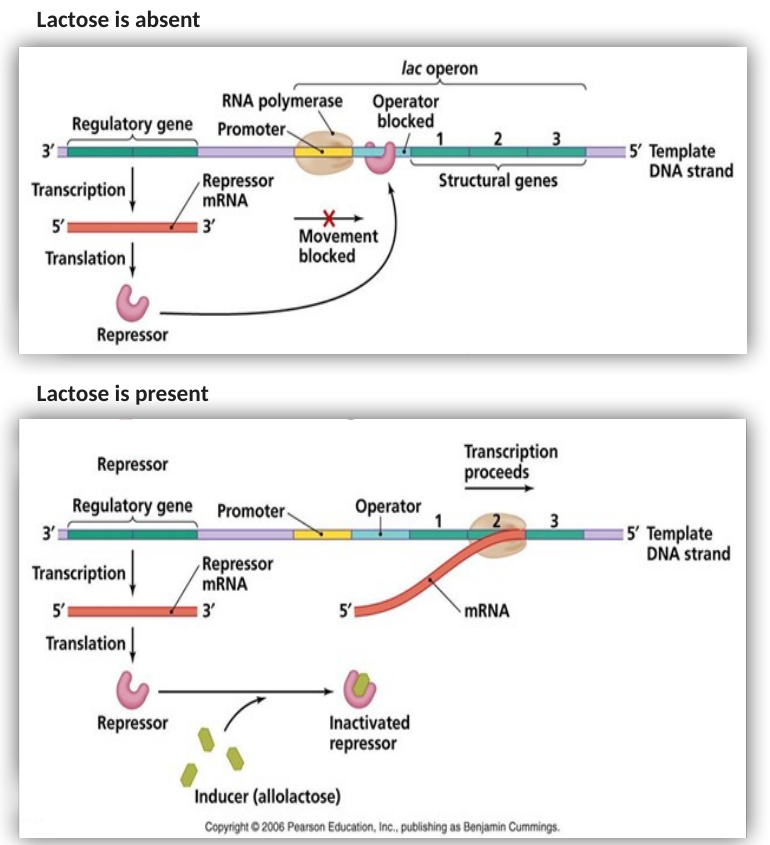
אופרון הלקטוז אחראי על ייצור חלבונים המפרקים לקטוז כדי להפיק ממנו אנרגיה. מאחר שלא תמיד יש לקטוז בסביבה, החיידק לא רוצה לבזבז אנרגיה על ייצור החלבונים האלו סתם.
- כשאין לקטוז: הגן הרגולטורי מייצר רפרסור. הרפרסור נקשר פיזית לאופרטור ומונע מה־RNA פולימראז להתקדם ולסנתז את הגנים. האופרון כבוי.
- כשיש לקטוז (הסטימולי): הלקטוז עצמו לא נקשר ישירות לרפרסור. חלבון מיוחד בשם LacZ עושה מודיפיקציה קטנה ללקטוז והופך אותו לאלו־לקטוז (Allolactose). האלו־לקטוז הוא זה שמזוהה על ידי הרפרסור. הוא נקשר אליו, משנה את הקונפורמציה שלו ומנתק אותו מהאופרטור. כעת הפולימראז חופשי לסנתז את הגנים.
Repressible operon (אופרון רפרסיבילי) - אופרון הטריפטופן
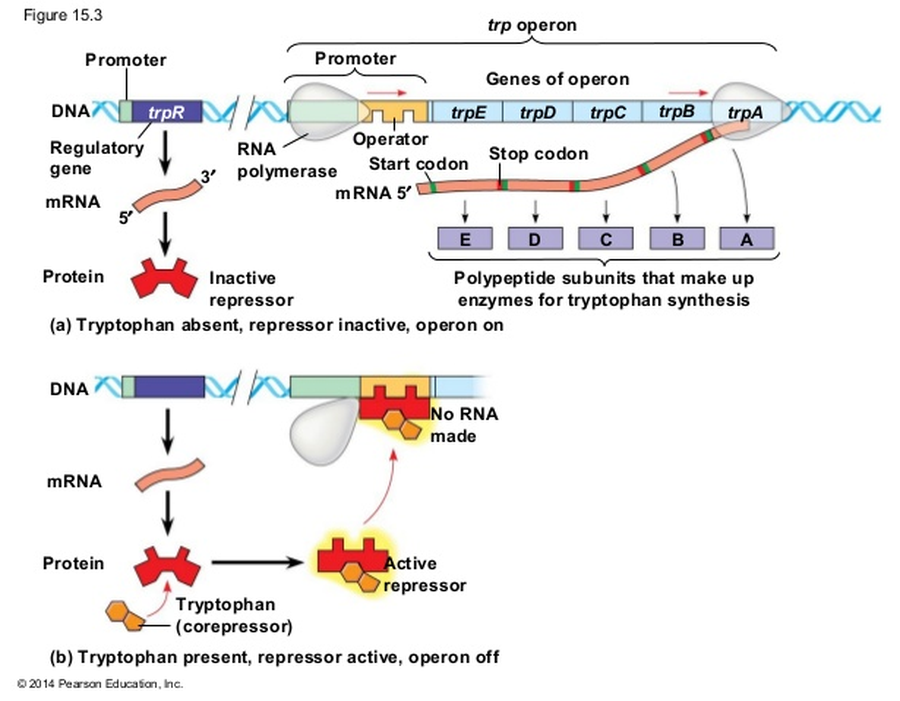
אופרון הטריפטופן כולל פרומוטור, אופרטור ו־5 גנים האחראים לסינתזה של חומצת האמינו טריפטופן, שהחיידק מייצר לעצמו (או מקבל מהסביבה). הרפרסור של אופרון זה מיוצר במצב לא פעיל.
- כשאין טריפטופן: הרפרסור לא פעיל ולא נקשר לאופרטור. האופרון פועל באופן קבוע, והחיידק משקיע אנרגיה ומסנתז לעצמו טריפטופן.
- כשיש טריפטופן (למשל, מגיע מהסביבה): לחיידק אין סיבה לבזבז אנרגיה על ייצור עצמי. הטריפטופן מהסביבה נקשר פיזית לרפרסור, משנה את הקונפורמציה שלו ומפעיל אותו. הרפרסור הפעיל נקשר לאופרטור וחוסם פיזית את ה־RNA פולימראז. האופרון כבוי.
| סוג אופרון | דוגמה | מצב חסר (אין סובסטרט בסביבה) | מצב נוכחות (יש סובסטרט בסביבה) |
|---|---|---|---|
| אינדוסיבילי | אופרון הלקטוז | כבוי - הרפרסור מונע סינתזה (אין צורך לפרק). | מופעל - לקטוז הופך לאלו־לקטוז, שמנתק את הרפרסור. |
| רפרסיבילי | אופרון הטריפטופן | פעיל - החיידק מסנתז לעצמו את המולקולה. | מדוכא - הטריפטופן החיצוני מפעיל את הרפרסור שעוצר את הסינתזה (לחיסכון באנרגיה). |
כיווניות גנים על גבי ה־DNA
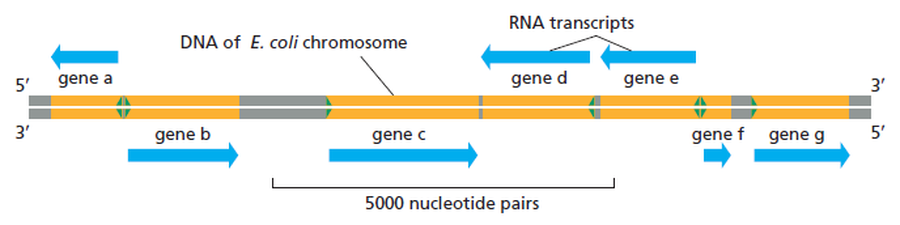
הגנום לא בנוי כך שיש גדיל אחד שהוא תמיד גדיל התבנית לכל אורכו.
גנים יכולים ללכת לשני הכיוונים על גבי ה־DNA, ולפעמים אף להימצא בחפיפה (Overlap) אחד על השני.
מה שקובע את הכיווניות הוא הפרומוטור:
- מיקומו וכיוונו של הפרומוטור קובעים לאיזה כיוון ינוע ה־RNA פולימראז.
- הפולימראז תמיד יסנתז מהפרומוטור והלאה (Downstream / במורד הזרם).
- השמות “תבנית” או “מקודד” תלויים אך ורק במיקום הגן הספציפי על הגדיל (ה־DNA עצמו אינו “מודע” לשמות האלה).
אנטיביוטיקות שפוגעות בשעתוק חיידקי
| תרופה | אתר הקישור/מטרה | מנגנון הפגיעה בשעתוק | שימושים עיקריים |
|---|---|---|---|
Rifampin (ריפאמפין)  | תת־יחידה β (בטא) של ה־RNA פולימראז | מונעת את תהליך ההתארכות (Elongation) | טיפול בשחפת (Tuberculosis) וזיהומי מעיים |
| Fidaxomicin (פידאקסומיצין) | תת־יחידה β (באזור פתיחת גדילי ה־DNA) | מונעת מהפולימראז לפתוח את גדילי ה־DNA ולכן אין סינתזה | זיהומי מעיים |
Actinomycin D (אקטינומיצין D)  | מולקולת ה־DNA עצמה | נכנסת לתוך ה־DNA, יוצרת איתו קשר ומונעת פיזית את התקדמות הפולימראז | מעבדות מחקר, טיפולים אונקולוגיים (אינה ספציפית לחיידקים ולכן פוגעת גם באדם) |
Bicyclomycin (ביציקלומיצין)  | חלבון Rho | מונעת את פעולת ה־Rho בטרמינציה - הפולימראז ממשיך לסנתז ללא הפסקה | מחלות מעיים, שימוש וטרינרי (בבעלי חיים) |