המשך סומיטוגנזה: שעון וגרדיאנט
בשיעור הקודם הוגדרו שני מרכיבים מרכזיים של סומיטוגנזה:
- גרדיאנט FGF8 במזודרם הפרה־סומיטי (Presomitic Mesoderm, PSM).
- גנים אוסילטוריים, כלומר גנים שהביטוי שלהם עולה ויורד במחזוריות.
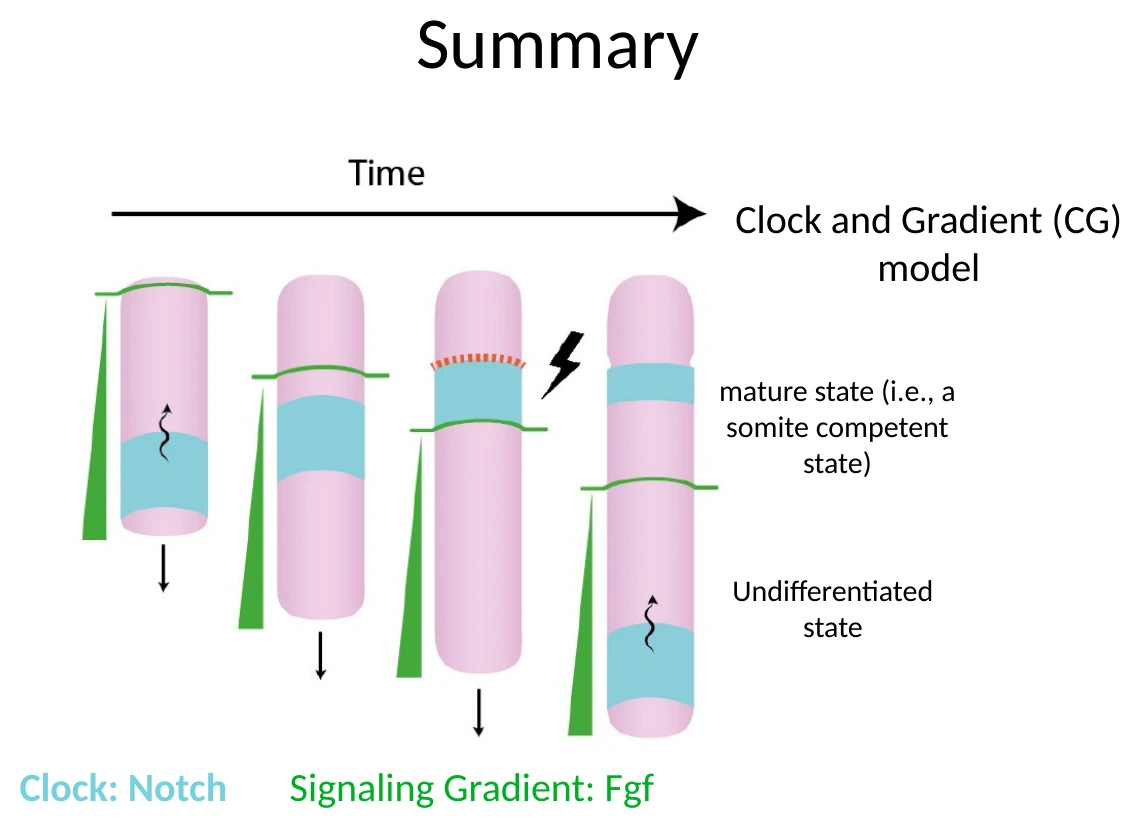
שני המרכיבים האלה יוצרים יחד את מודל Clock-and-Gradient. המודל המקורי נקרא Clock-and-Wavefront, אבל בשיעור הודגש שה״החזית״ אינה רק קו שמתקדם אחורה, אלא גרדיאנט סיגנלי, בעיקר של FGF signaling. לכן המודל המדויק יותר כאן הוא שעון וגרדיאנט.
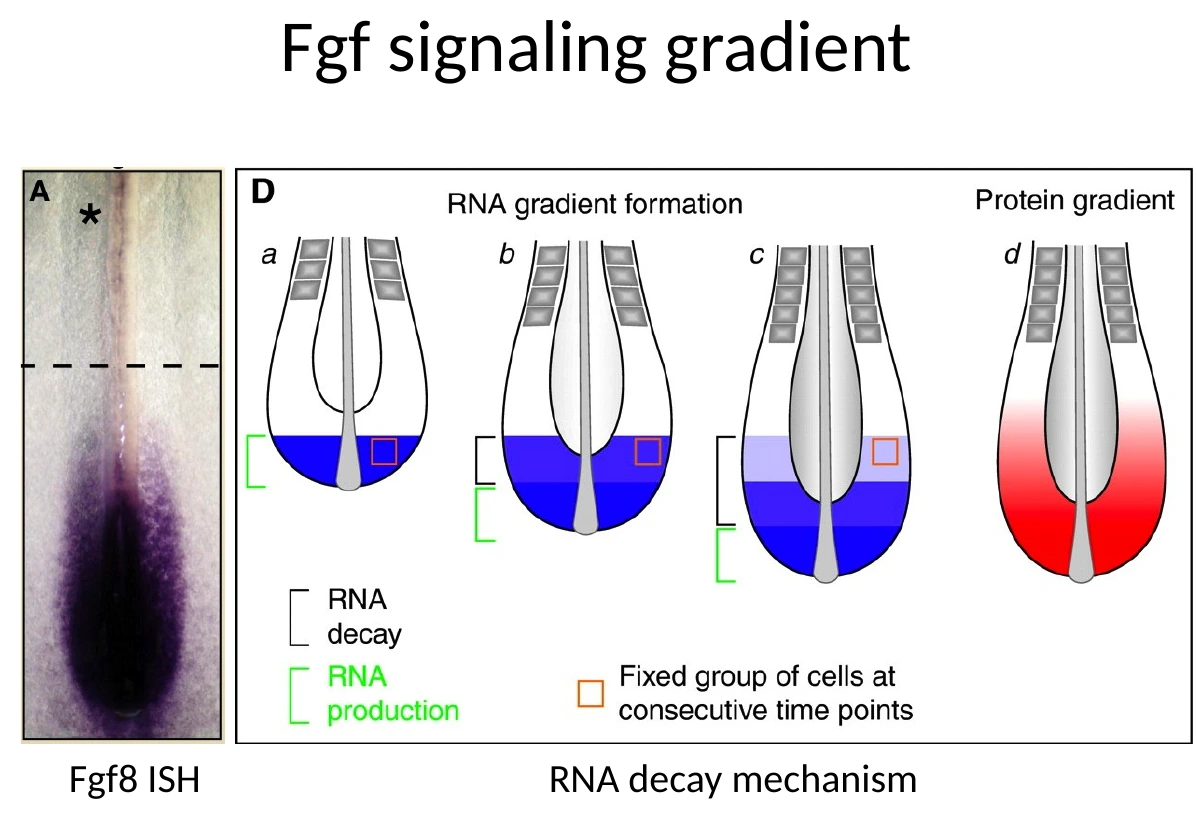
איך נוצר גרדיאנט FGF8
המקור של FGF8 נמצא באזור ה־Tailbud. שם יש שעתוק חדש של Fgf8. במצגת מופיע גם ציר של FGF/Wnt gradient; בשיעור הדגש היה בעיקר על FGF כגרדיאנט שמחזיק את ה־PSM במצב לא ממוין ומגדיר את אזור הבשלות. בזמן Axis elongation, תאים שהיו קרובים ל־Tailbud ״נשארים מאחור״ יחסית, כי הציר מתארך לכיוון פוסטריורי. בתאים האלה השעתוק החדש של Fgf8 נפסק, וה־mRNA שכבר קיים מתחיל להתפרק.
לכן מתקבל גרדיאנט:
- באזור הפוסטריורי, קרוב ל־Tailbud - רמות FGF8 גבוהות.
- ככל שמתקדמים אנטריורית בתוך ה־PSM - רמות FGF8 יורדות.
זהו RNA decay mechanism: לא צריך לייצר שעתוק חדש בכל נקודה; מספיק שהשעתוק יישאר ב־Tailbud, ושה־mRNA יתפרק בהדרגה בתאים שמתרחקים ממנו.
תפקיד גרדיאנט ה־FGF
רמות גבוהות של FGF signaling שומרות על תאי ה־PSM במצב פחות ממוין, שבו הם עדיין לא מוכנים להפוך לסומיט.
כאשר ה־Tailbud מתרחק פוסטריורית, גם גרדיאנט FGF ״נסוג״ לאחור. כך נוצר אזור אנטריורי ב־PSM שבו FGF כבר נמוך. באזור הזה התאים נמצאים במצב בוגר יותר (somite-competent state): הם יכולים להפוך לסומיט, אבל הם עדיין צריכים לקבל את הפולס של השעון.
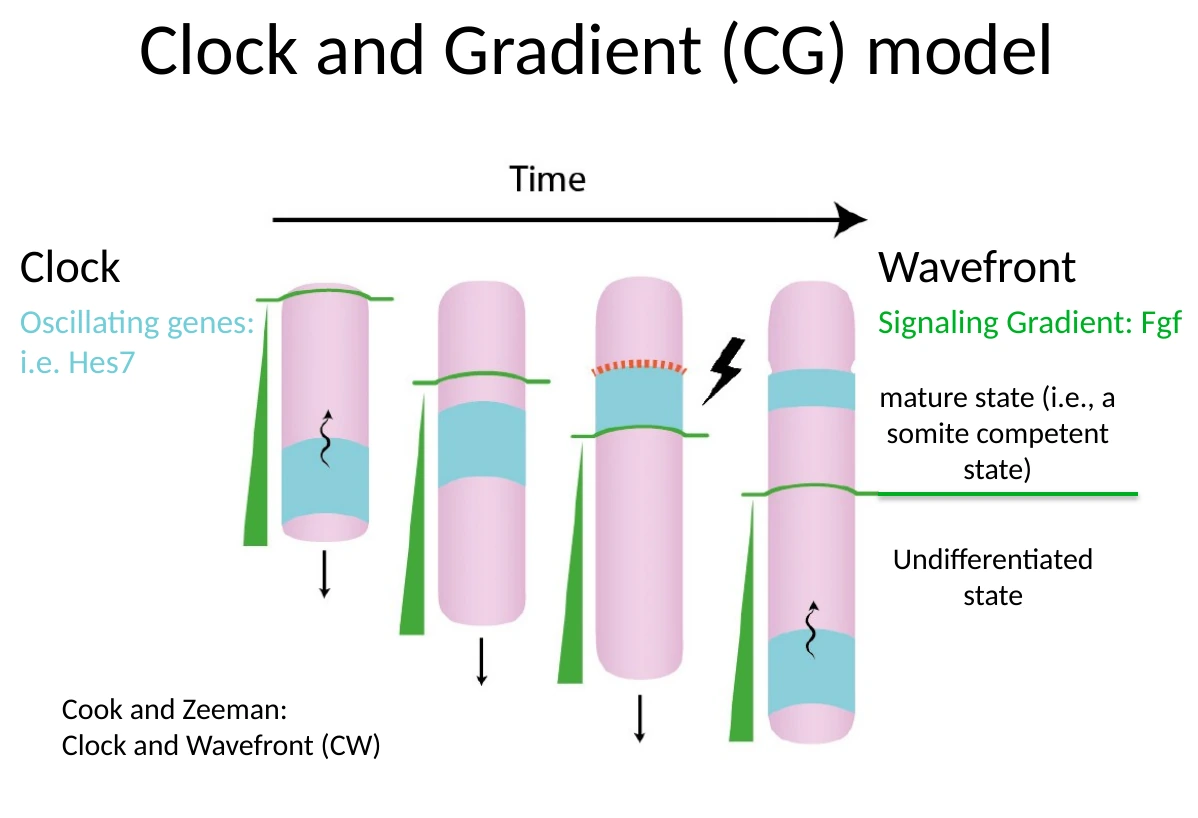
העיקרון:
| מרכיב | תפקיד |
|---|---|
| FGF gradient | קובע איפה התאים כבר מספיק בוגרים כדי ליצור סומיט |
| שעון אוסילטורי | קובע מתי תופעל תוכנית הסגמנטציה |
| Mesp2 | מסמן את תחילת תוכנית הסגמנטציה באזור שהבשיל |
Hes7: גן שעון ולא גן גורל
בעובר תרנגולת, אחד הגנים האוסילטוריים שהוצגו הוא cHairy1. בעכבר ובאדם הדוגמה המרכזית היא Hes7.
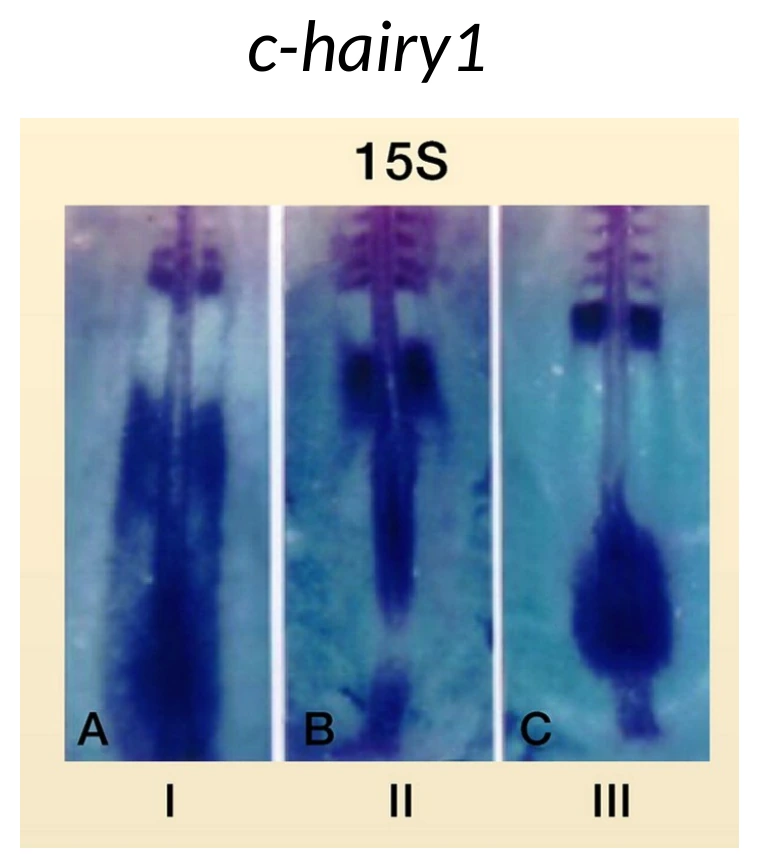 | 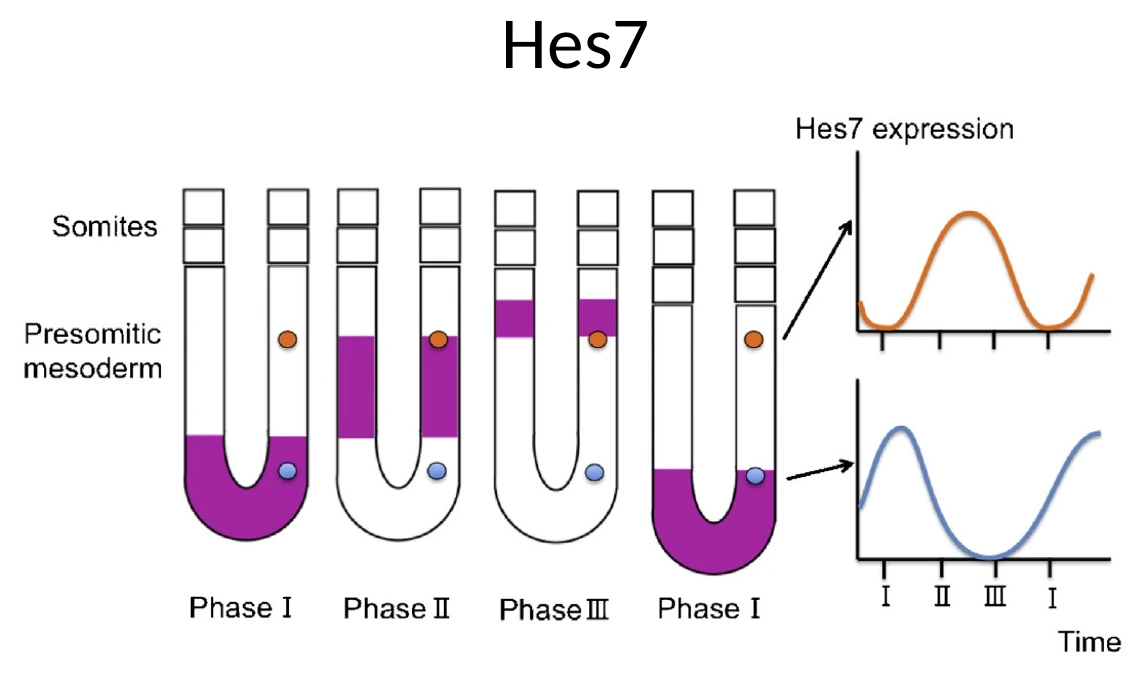 |
חשוב להבין: זה לא RNA ש״זורם״ פיזית מה־Tailbud לכיוון אנטריורי. כל תא ב־PSM מבטא את Hes7 באופן מחזורי, אבל תאים שונים נמצאים בפאזות שונות של המחזור.
לדוגמה:
- תא אחד יכול להיות בשלב שבו Hes7 גבוה.
- תא אחר, באותו זמן בדיוק, יכול להיות בשלב שבו Hes7 נמוך.
- לאורך זמן, כל תא עובר עלייה וירידה בביטוי.
לכן, בתמונות של in situ, זה נראה כאילו גל ביטוי מתקדם לאורך ה־PSM, אבל בפועל מדובר באוכלוסיית תאים שכל אחד מהם מתנדנד, והפאזות שלהם מסודרות במרחב.
Loss of function של Hes7
כאשר עושים loss of function ל־Hes7, העוברים יכולים להגיע לשלב מאוחר ולהיוולד, אבל יש פגיעה קשה בסגמנטציה של השלד.
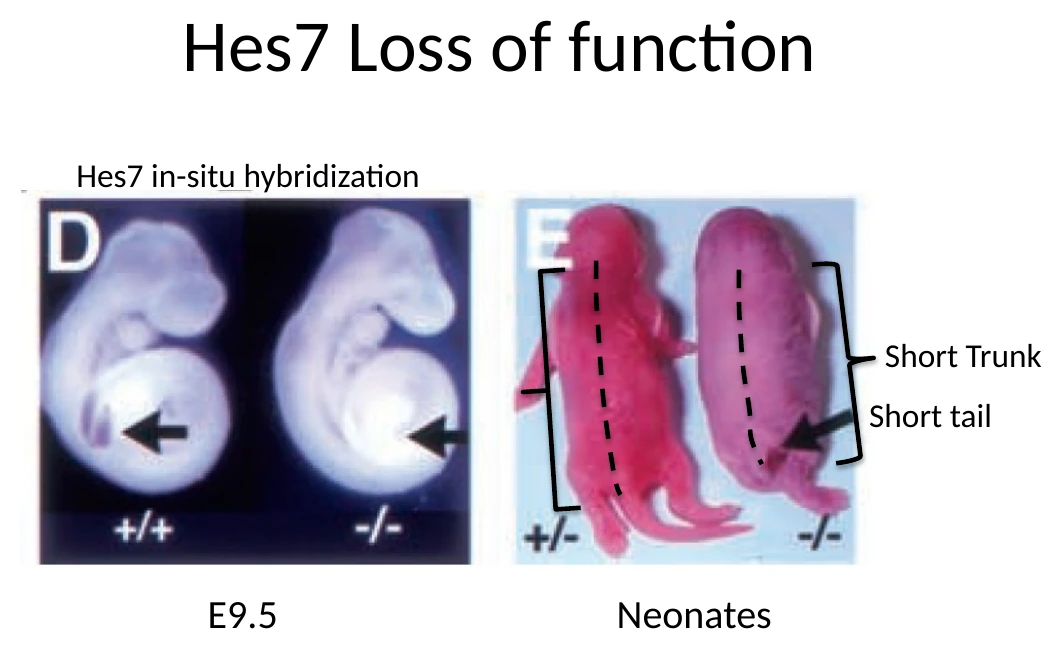
הפנוטיפ המרכזי:
| תצפית | פירוש |
|---|---|
| עכברים נולדים עם גֵּו קצר וזנב קצר | יש פגיעה בבניית הציר האחורי והחוליות |
| סומיטים נוצרים בצורה לא סדירה ב־E9.5 | יש בעיה בפרגמנטציה של ה־PSM |
| חוליות וצלעות נראות מאוחות או לא מסודרות | הסגמנטציה של השלד נפגעה |
| חוט השדרה עדיין קיים | הבעיה אינה יצירת חוט השדרה עצמו, אלא תבנית החוליות סביבו |
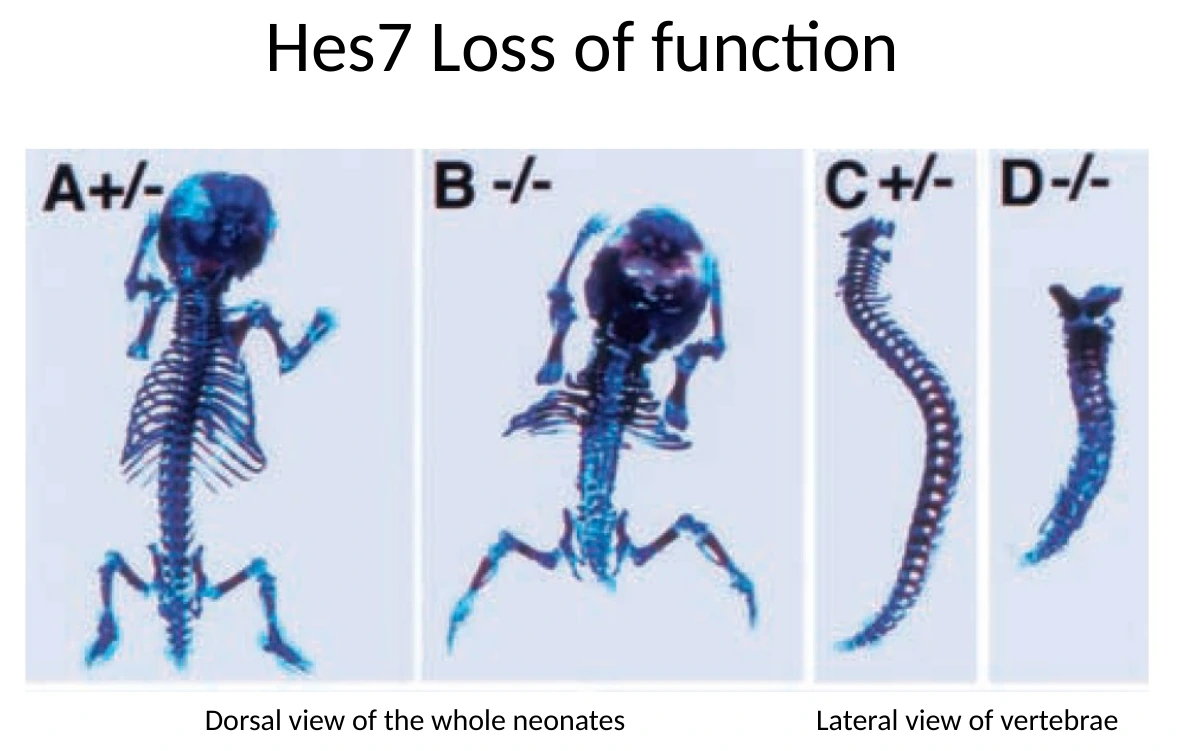 | 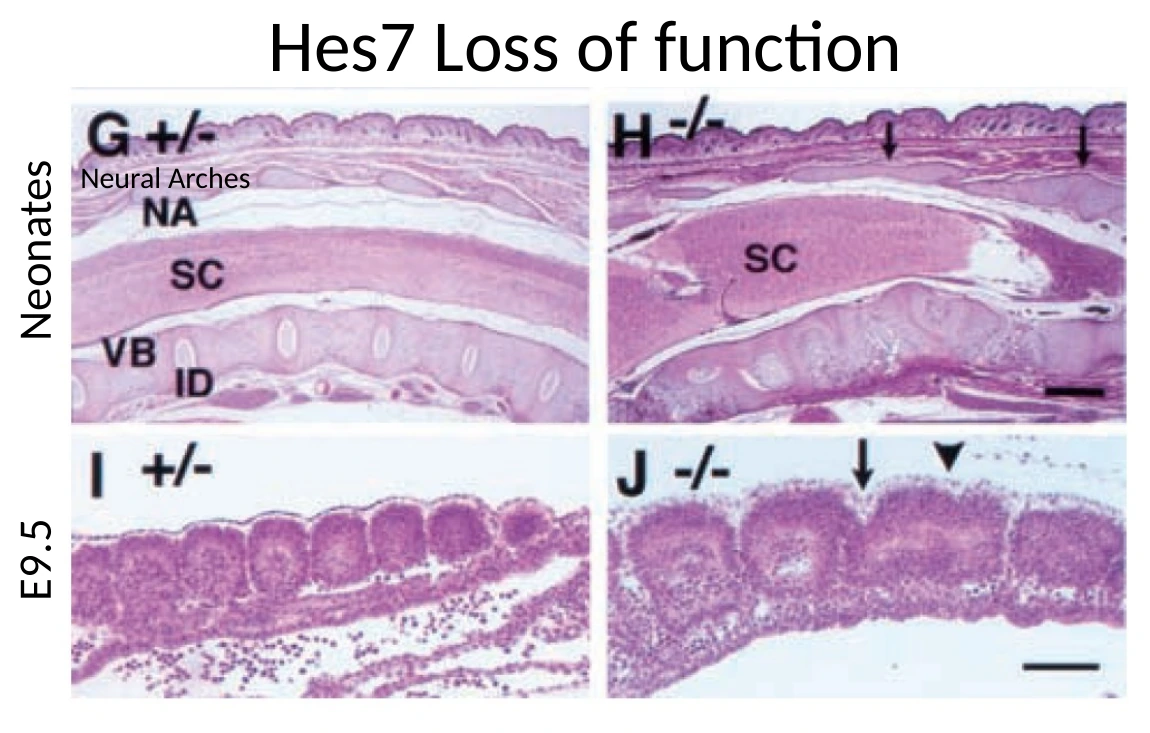 |
המסקנה אינה ש־Hes7 ״יוצר שלד״. השלד עדיין יכול להיווצר במידה מסוימת. הבעיה היא שאין סגמנטציה תקינה: במקום הרבה יחידות מסודרות, מתקבל רצף מאוחה ולא מאורגן.
Gain of function של Hes7
בניסוי gain of function, הביטוי של Hes7 נעשה קבוע ומתמשך בכל ה־PSM. גם כאן מתקבל פנוטיפ דומה: סומיטים וחוליות מתאחים.

המסקנה החשובה היא שצריך את המחזוריות של Hes7, לא רק את הנוכחות שלו. Hes7 אינו גן שמכתיב ״מה יהיה הסומיט״, אלא גן שמשמש כחלק מהשעון שקובע מתי תתרחש הפרגמנטציה.
אפשר לנסח את זה כך:
- בלי Hes7 - אין שעון תקין.
- עם Hes7 קבוע ולא מחזורי - גם אין שעון תקין.
- רק ביטוי מחזורי של Hes7 מאפשר סגמנטציה תקינה.
יצירת ביטוי מחזורי של Hes7
Hes7 הוא transcription factor שפועל כרפרסור. הוא מדכא את השעתוק של עצמו, ולכן יוצר משוב שלילי.
 | 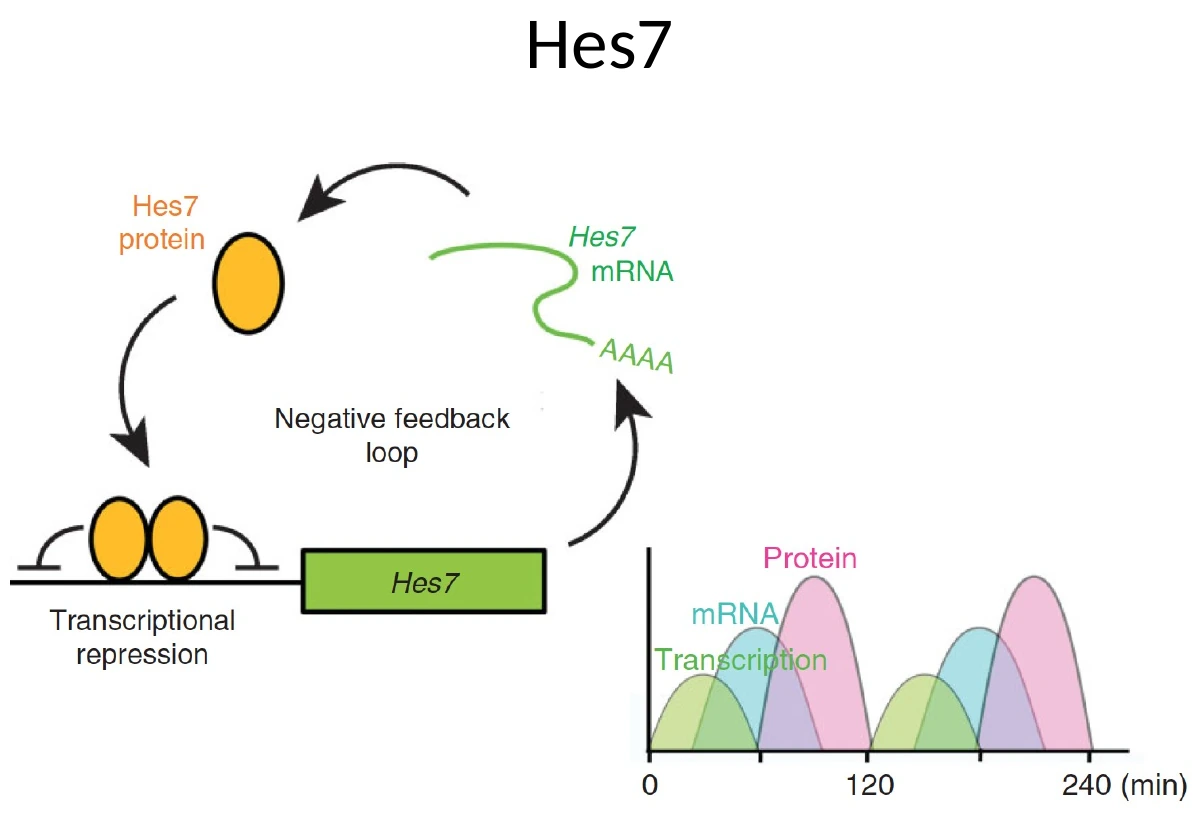 |
הרצף הבסיסי:
- Notch ו־FGF מאפשרים שעתוק של Hes7.
- נוצר mRNA של Hes7.
- ה־mRNA עובר splicing ויוצא לציטופלזמה.
- מתורגם חלבון Hes7.
- החלבון חוזר לגרעין.
- Hes7 מדכא את הפרומוטור של Hes7 עצמו.
- בגלל שה־mRNA והחלבון לא יציבים, הם מתפרקים.
- לאחר שהחלבון נעלם, הדיכוי משתחרר, ומתחיל מחזור חדש.
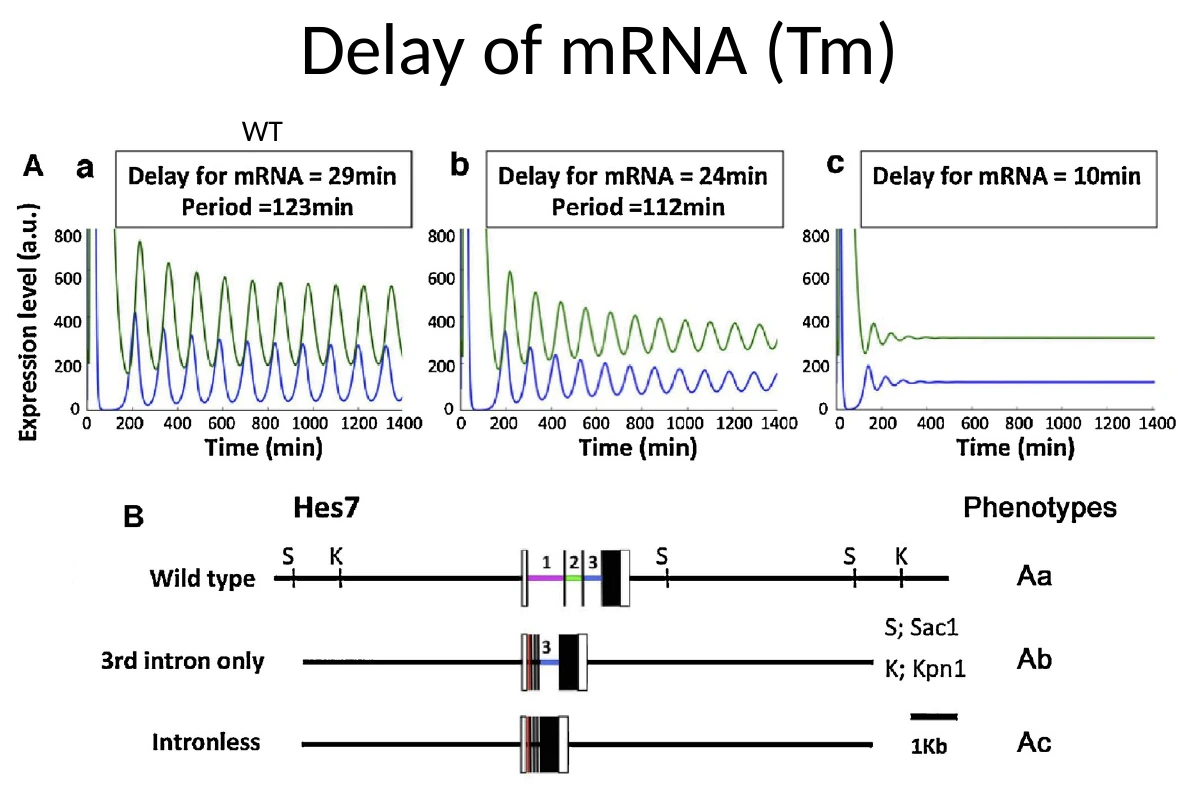
למה צריך דיליי
כדי שמשוב שלילי ייצור אוסילציה, לא מספיק שהחלבון ידכא את עצמו. צריך גם דיליי בין תחילת השעתוק לבין הופעת החלבון הפעיל.
הדיליי כולל כמה שלבים:
- שעתוק
- splicing
- יציאת mRNA מהגרעין
- תרגום
- הבשלת החלבון והגעה לגרעין
ניתן לפרמל את המודל בעזרת delay differential equations. הנקודה הביולוגית העיקרית היא שמשך החיים של ה־mRNA והחלבון, יחד עם הדיליי, קובעים אם תתקבל אוסילציה יציבה.
\[\begin{aligned} &\mathrm{d}p(t)/\mathrm{d}t = \mathrm{a}m(t-T_p) - \mathrm{b}p(t) \\[1em] &\mathrm{d}m(t)/\mathrm{d}t = \mathrm{k}/\left[1 + \left\{\mathcal{p}(t-T_m)/\mathrm{p_0}\right\}^2\right] - \mathrm{c}m(t) \end{aligned}\]הפרמטרים במודל המתמטי של Hes7: כמות mRNA, כמות חלבון, קצבי תרגום ופירוק, ודיליי של mRNA וחלבון.
| פרמטר | משמעות ביולוגית |
|---|---|
| m(t) | כמות mRNA פעיל בזמן נתון |
| p(t) | כמות חלבון Hes7 פעיל בזמן נתון |
| b | קצב פירוק של החלבון |
| c | קצב פירוק של ה־mRNA |
| Tm | הזמן מתחילת השעתוק עד mRNA בשל בציטופלזמה |
| Tp | הזמן מתחילת התרגום עד חלבון בשל ופונקציונלי |
שני תנאים חשובים לאוסילציה יציבה:
- זמן מחצית החיים של תוצרי Hes7 צריך להיות קצר מספיק - גם ה־mRNA וגם החלבון צריכים להתפרק מהר יחסית.
- הדיליי הכולל צריך להיות ארוך מספיק - חייב להיות פער זמן בין תחילת השעתוק לבין הופעת החלבון המדכא.
שני תנאים לאוסילציה של Hes/her: תוצרים לא יציבים מספיק, ודיליי כולל ארוך מספיק.
שינוי קטן בזמן מחצית החיים משנה את הסגמנטציה
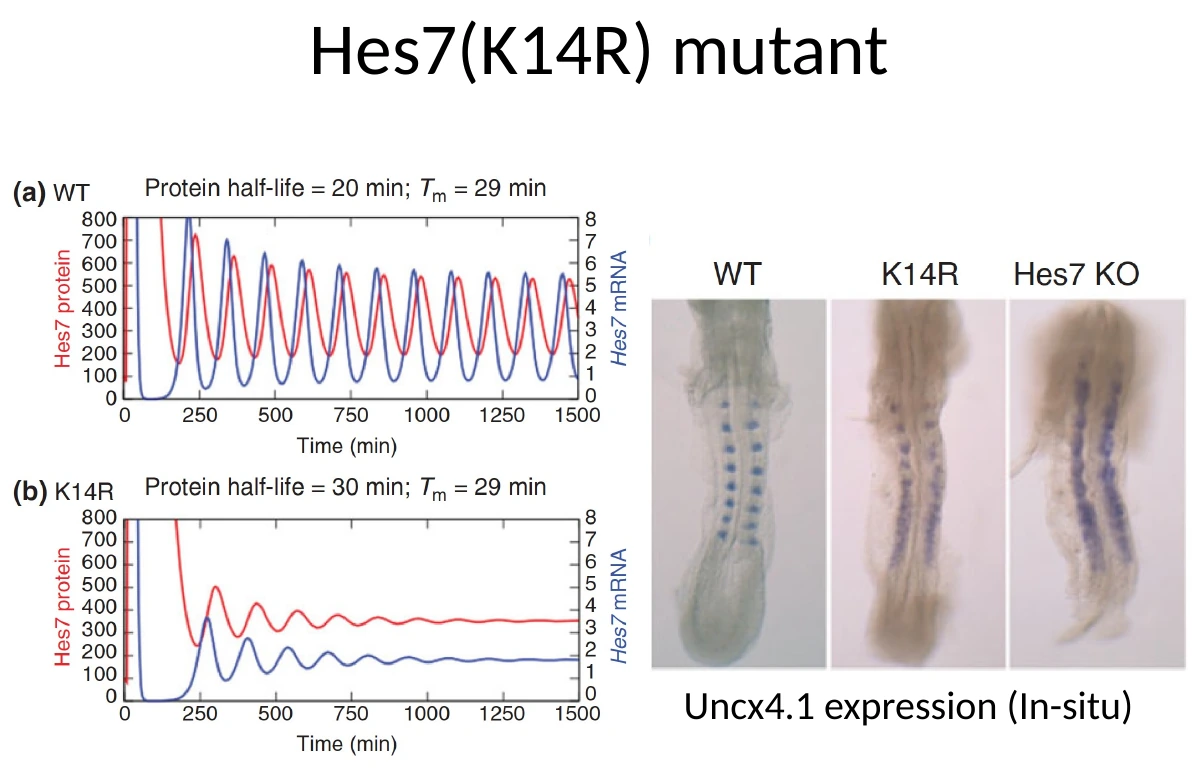
במצב תקין של Hes7 בעכבר:
- זמן מחצית החיים של החלבון הוא בערך 20 דקות.
- הדיליי של ה־mRNA הוא בערך 29 דקות.
- המחזור כולו מתאים לקצב יצירת סומיט בעכבר: בערך שעתיים.
במוטציה Hes7(K14R) מחליפים חומצת אמינו אחת: lysine בעמדה 14 מוחלפת ב־arginine. זו החלפה יחסית עדינה מבחינת מטען, אבל היא מאריכה את זמן מחצית החיים של החלבון לכ־30 דקות.
התוצאה: האוסילציה דועכת לאחר מספר מחזורים, ולכן הסומיטים הראשונים יכולים להיווצר, אבל הסומיטים הבאים מתאחים.
 | 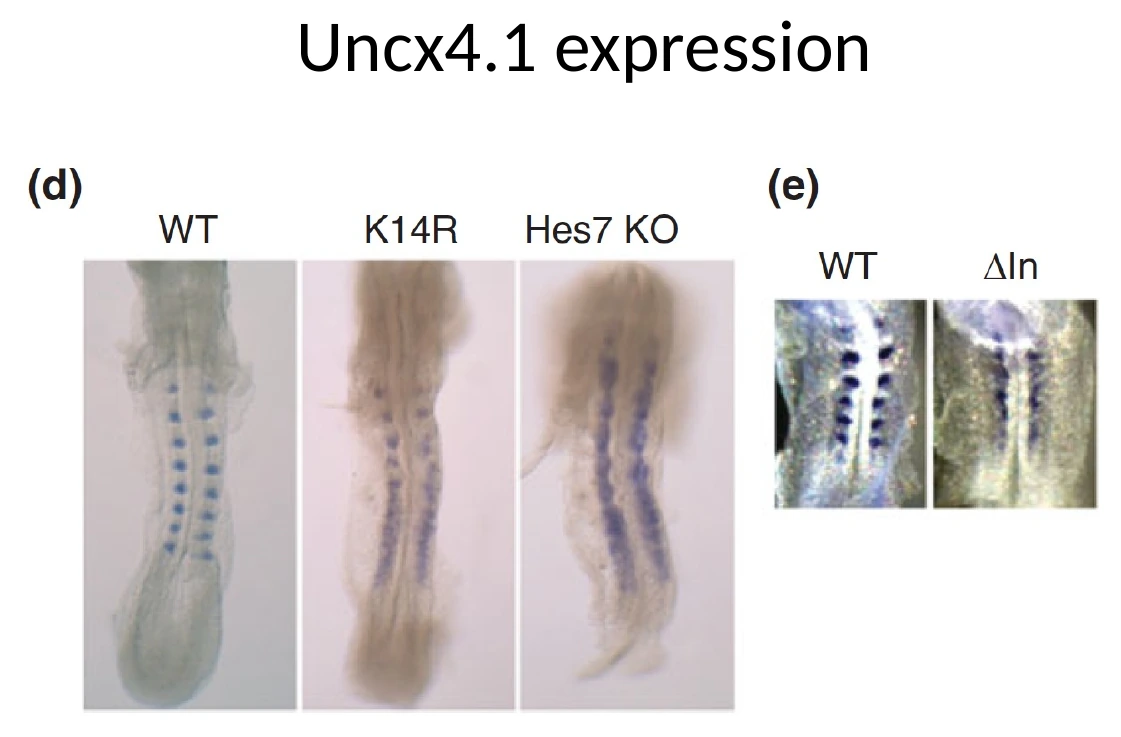 |
אינטרונים יוצרים חלק מהדיליי
חלק מהדיליי של Hes7 נובע מ־splicing. אם מוציאים אינטרונים מהגן, מקצרים את זמן העיבוד של ה־mRNA.
במודל שהוצג:
| מצב הגן | השפעה על הדיליי | תוצאה צפויה |
|---|---|---|
| Wild type | Tm בערך 29 דקות | אוסילציה תקינה |
| נשאר רק אינטרון אחד | Tm מתקצר לכ־24 דקות | המחזור מתקצר והאוסילציה פחות יציבה |
| גן בלי אינטרונים | Tm מתקצר מאוד, לכ־10 דקות | האוסילציה מתבטלת וסומיטים מתאחים |
לכן האינטרונים בהקשר הזה תורמים לזמן העיבוד של ה־mRNA, והזמן הזה הוא חלק ממנגנון השעון.
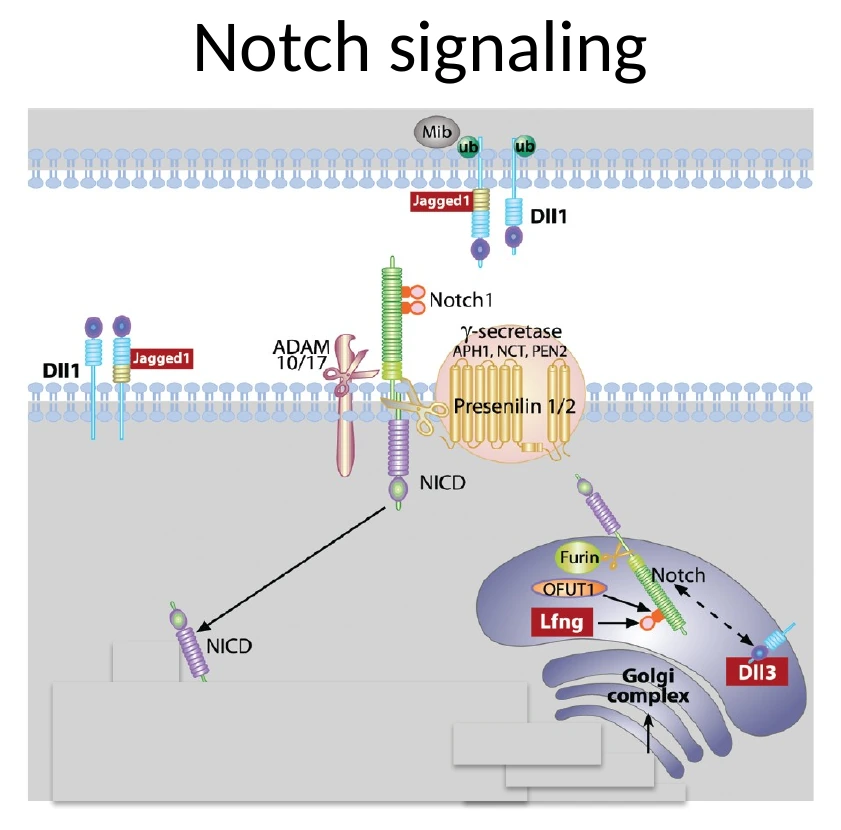
Notch, Lunatic Fringe וסנכרון השעון
עד כאן אפשר היה לחשוב שכל תא ב־PSM מפעיל לבד את השעון שלו. אבל כדי לייצר סומיטים תקינים, התאים צריכים להיות מסונכרנים זה עם זה. כאן נכנס Notch signaling.
Lunatic Fringe הוא חלק ממסלול Notch
Lunatic Fringe (Lfng) הוא אנזים בגולג׳י. הוא משנה את Notch באמצעות גליקוזילציה, וכך משפיע על היכולת של Notch להגיב לליגנדים שלו.
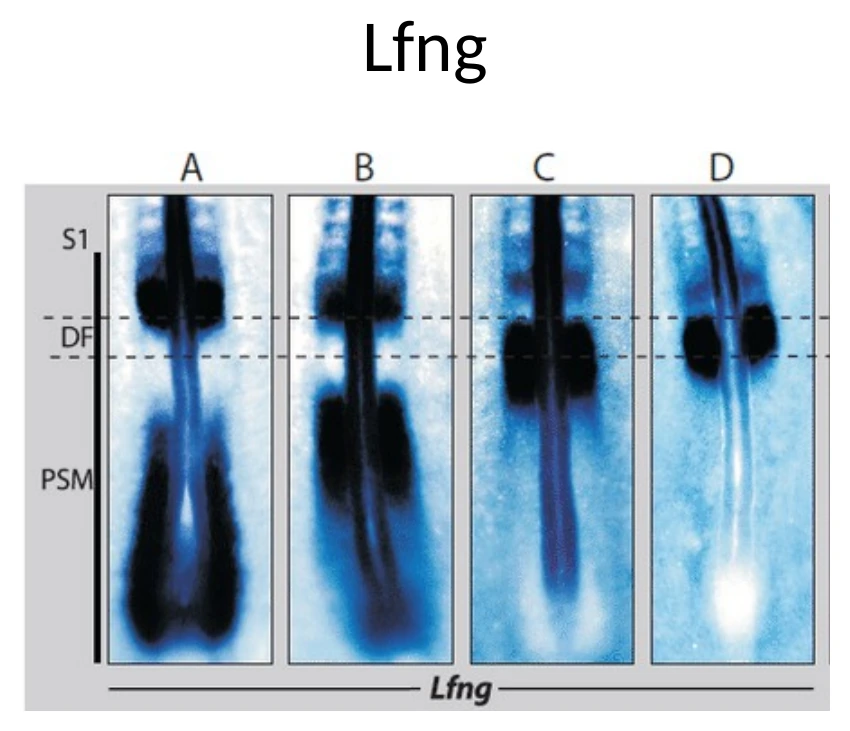
בשיעור הודגש ש־Lfng עצמו מתבטא בדפוס מחזורי, כמו Hes7. זה חשוב, כי הוא מחבר בין השעון לבין Notch signaling.
תזכורת למסלול Notch:
- תא אחד מבטא ליגנד כמו Delta-like.
- תא סמוך מבטא את הרצפטור Notch.
- קשירת הליגנד גורמת לביקועים פרוטאוליטיים ב־Notch.
- משתחרר Notch intracellular domain.
- הדומיין הזה נכנס לגרעין ומשפיע על שעתוק גנים.
Notch נדרש לסנכרון
בניסוי בדג זברה עיכבו Notch באמצעות DAPT. לאחר העיכוב, הביטוי של הגן האוסילטורי Her1 יצא מסנכרון, והפך לדגם מפוזר של salt-and-pepper.
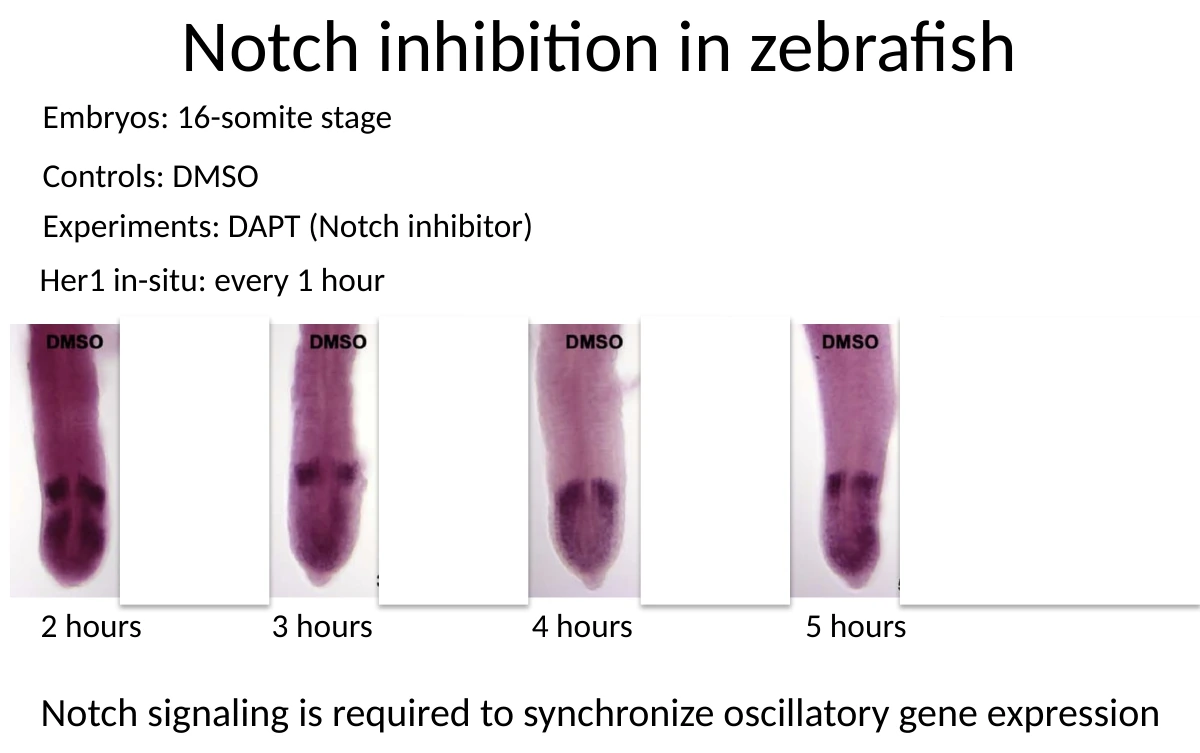
המסקנה:
- תאים יכולים להחזיק מנגנון אוסילטורי פנימי.
- Notch מסנכרן את התאים אחד עם השני.
- בלי Notch, כל תא יכול להמשיך להתנדנד, אבל הפאזות כבר לא מתואמות, ולכן לא מתקבל גל מסודר.
הקשר בין Notch, Hes7 ו־Lfng
Notch מפעיל שעתוק של Hes7 ושל Lfng. ה־Hes7, בתור רפרסור, מדכא גם את עצמו וגם את Lfng. ה־Lfng משנה את פעילות Notch, וכך נוצר משוב שמאפשר תנודתיות וסנכרון.
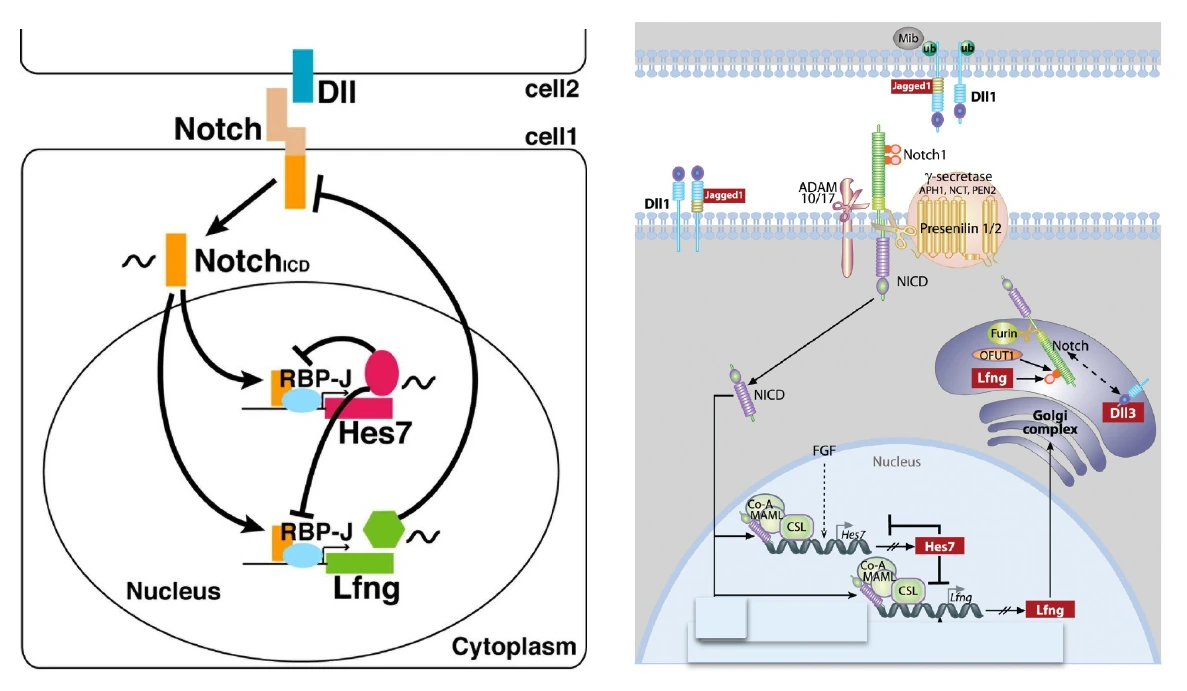
התמונה הכללית:
- FGF מאפשר ביטוי של Hes7 באזור שבו התאים עדיין ב־PSM.
- Notch מפעיל Hes7 ומסנכרן בין תאים סמוכים.
- Hes7 יוצר משוב שלילי שמייצר מחזוריות.
- Lfng משנה את Notch ומוסיף שכבת בקרה מחזורית.
Mesp2 ותוכנית הסגמנטציה
כדי שסומיט חדש ייווצר, לא מספיק שתאים יהיו ב־PSM ולא מספיק שיהיה שעון. צריך להפעיל בפועל את תוכנית הסגמנטציה.
הגן המרכזי כאן הוא Mesp2.
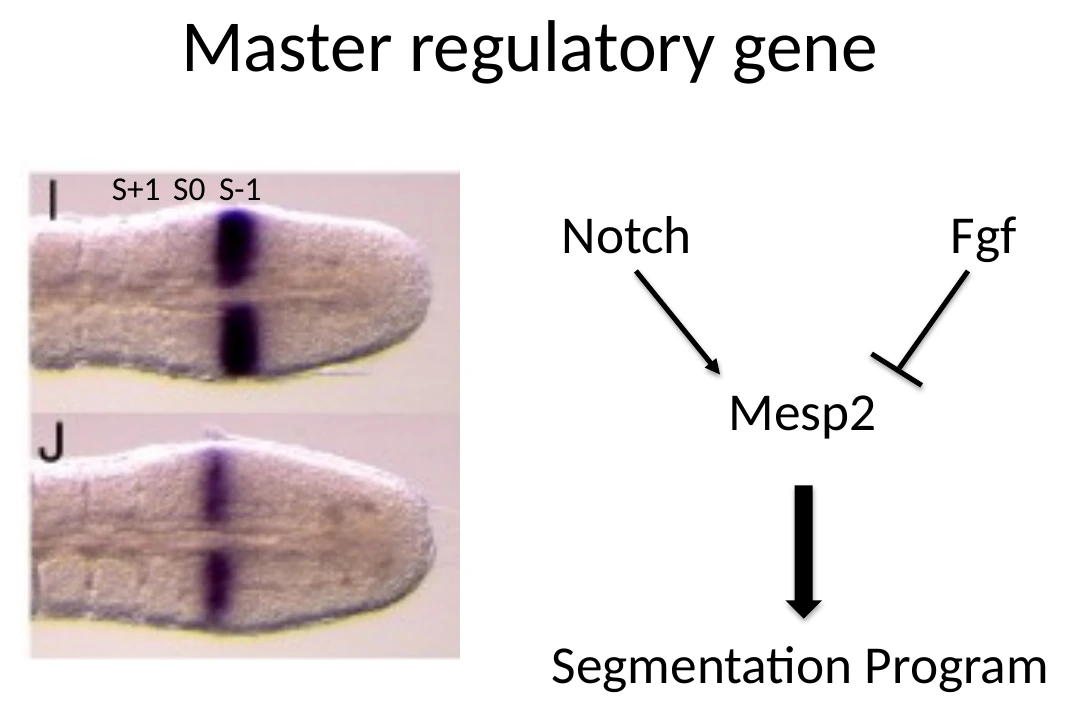
Mesp2 הוא גן בקרה ראשי של יצירת הסומיט (master regulatory gene). הוא מופיע בחלון זמן קצר, באזור שבו הסומיט הבא עומד להיווצר.
התנאים לביטוי Mesp2:
- התאים צריכים להיות באזור שבו FGF נמוך - כלומר באזור שבו הם כבר יכולים להתמיין.
- צריך להגיע פולס מתאים של Notch activity.
- קיימים גורמים נוספים, למשל TBX6, שמאפשרים את תגובת התאים.
לכן:
- הגרדיאנט הפוסטריורי, בעיקר FGF ובהקשר הרחב גם Wnt, קובע איפה התאים עדיין לא בשלים ואיפה הם כבר יכולים להיכנס לתוכנית סגמנטציה.
- השעון של Notch/Hes7 קובע מתי Mesp2 יופעל.
- Mesp2 מפעיל את תוכנית הסגמנטציה עצמה.
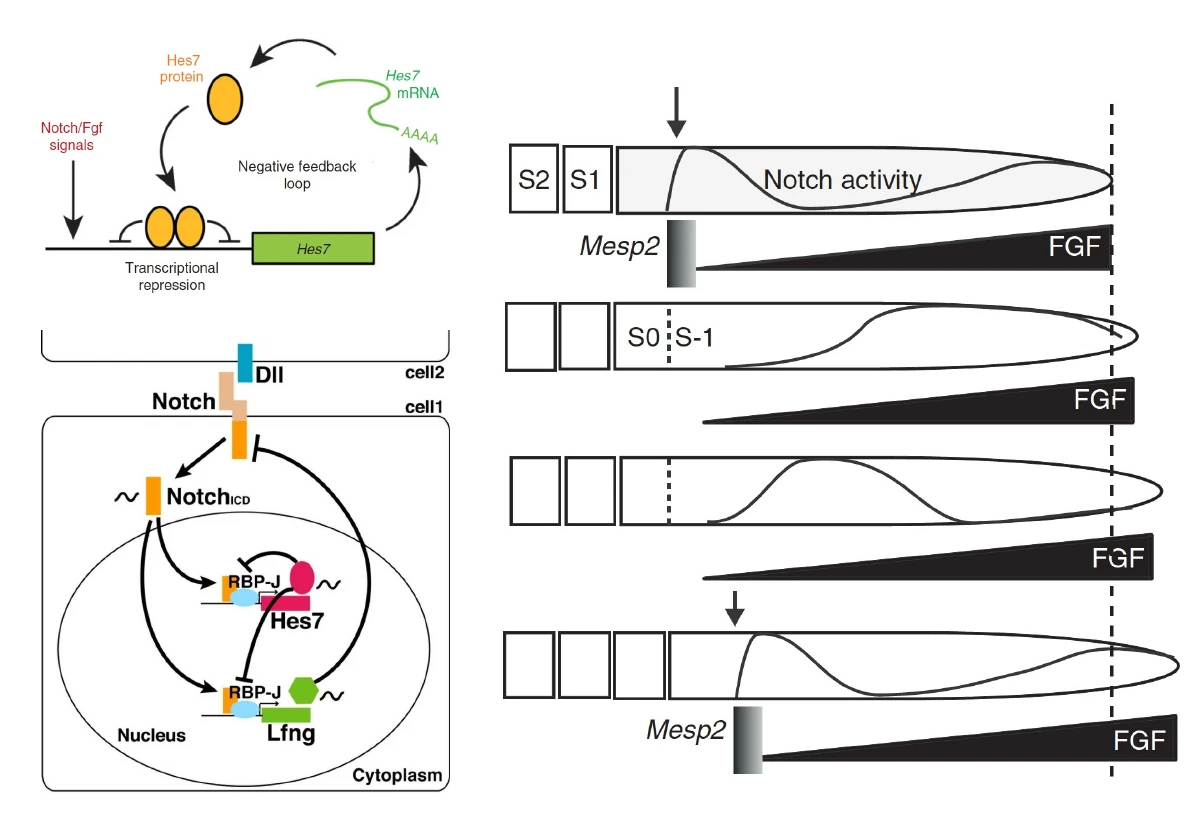
S-1, S0 ו־S1
בסומיטוגנזה משתמשים בסימון טופוגרפי:
| סימון | משמעות |
|---|---|
| S-1 | האזור ב־PSM שבו הסומיט הבא עומד להיווצר |
| S0 | הסומיט שנוצר זה עתה |
| S1 | הסומיט שכבר נוצר במחזור הקודם |
Mesp2 מופיע באזור הסומיט העתידי, ומכתיב את הגבול שבו תתרחש הפרגמנטציה. הוא לא צריך להישאר דולק לאורך זמן; להפך, ביטוי ממושך מדי יפגע בסדר של התהליך.
Spondylocostal dysostosis: כשסגמנטציה משתבשת
הקשר לרפואה מופיע בקבוצת מחלות בשם Spondylocostal dysostosis (SCDO). אלה מצבים שבהם יש פגיעה בסגמנטציה של החוליות והצלעות.
Spondylocostal dysostosis: פגיעה בסגמנטציה של חוליות וצלעות, לרוב עם גֵּו קצר, צוואר קצר וסקוליוזיס.
מאפיינים מרכזיים:
- פגמים מרובים בסגמנטציה של החוליות.
- פגיעות בצלעות.
- גֵּו (trunk) קצר ביחס לגובה.
- צוואר קצר.
- לעיתים סקוליוזיס לא מתקדם.
הגנים המעורבים קשורים בדיוק למסלולים שנלמדו בשיעור:
| גן | קשר לשיעור |
|---|---|
| DLL3 | ליגנד/רכיב במסלול Notch |
| HES7 | גן שעון אוסילטורי |
| LFNG | Lunatic Fringe, בקרה על Notch |
| MESP2 | מפעיל תוכנית סגמנטציה |
| RIPPLY2 | חלק מהתוכנית downstream של Mesp2 |
| TBX6 | גורם שמאפשר סגמנטציה תקינה |
הנקודה החשובה: אותם גנים שנראים במודל העכבר כחלק ממנגנון סומיטוגנזה הם גם גנים שמוטציות בהם גורמות למחלה אנושית.
קביעת מספר הסומיטים
מספר הסומיטים נקבע לפי היחס בין שני תהליכים:
- Axis elongation - כמה מהר ה־Tailbud מוסיף רקמה פוסטריורית.
- קצב השעון הסגמנטלי - כל כמה זמן נוצר סומיט חדש.
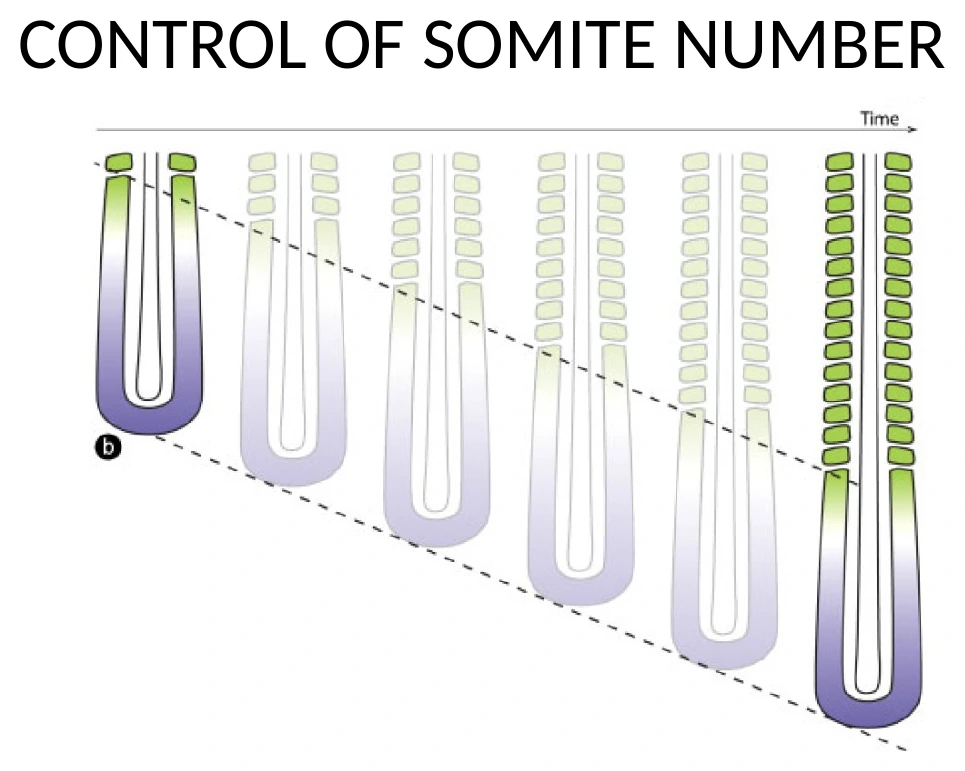
אם ציר ה־elongation מהיר ביחס לשעון, יש יותר זמן/מרחב לייצר סומיטים נוספים. אם השעון מהיר, נוצרים סומיטים בקצב גבוה יותר. אם השעון איטי, נוצרים פחות סומיטים באותו מרחב התפתחותי.
למה אי אפשר פשוט למדוד אורך בזמן אמיתי
אי אפשר להשוות ישירות ״כמה מילימטרים הציר התארך בשעה״ בין עכבר, נחש, תרנגולת ודג זברה, כי לכל מין יש קצב התפתחות כללי שונה. לכן משתמשים במדד אחר: מספר חלוקות תאים (number of generations), במהלך התקופה שבה נוצר ה־PSM והסומיטים.
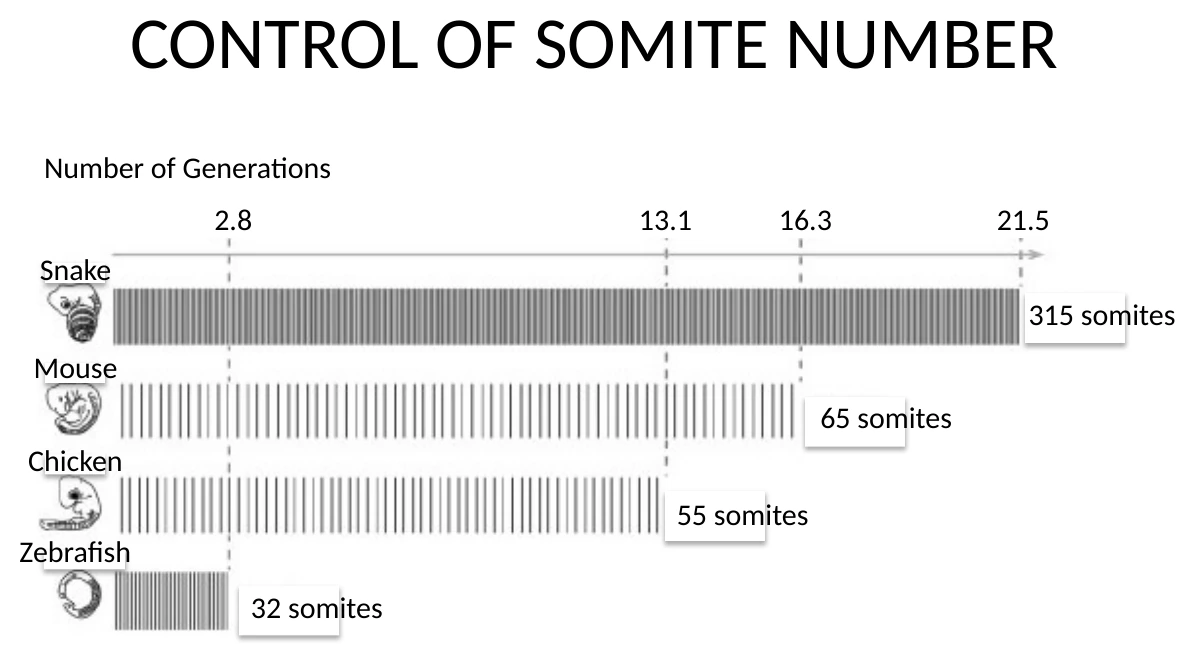
| מין | זמן ליצירת סומיט | מספר סומיטים בקירוב | עיקרון |
|---|---|---|---|
| דג זברה | כ־30 דקות | כ־32 | שעון מהיר, Axis elongation קצר יחסית |
| עכבר | כ־2 שעות | כ־65 | שעון איטי יותר |
| תרנגולת | כ־1.5 שעות | כ־55 | קצב ביניים |
| נחש | מספר סומיטים גבוה מאוד, למשל סביב 315 | הרבה יותר סומיטים | לא מוסבר רק על ידי יותר חלוקות; גם קצב השעון שונה |
הנקודה המרכזית: בנחש יש הרבה יותר סומיטים מאשר בעכבר, אבל ההבדל במספר חלוקות התאים לא מספיק גדול בשביל להסביר לבד את הפער במספר הסומיטים. לכן צריך לקחת בחשבון גם את קצב השעון.
סומיטים רבים יותר לא חייבים להיות גדולים יותר. אם השעון רץ מהר יותר, אפשר ליצור יותר סומיטים, שכל אחד מהם קטן יותר בתחילת הדרך; לאחר מכן הם יכולים לגדול ולהתפתח.
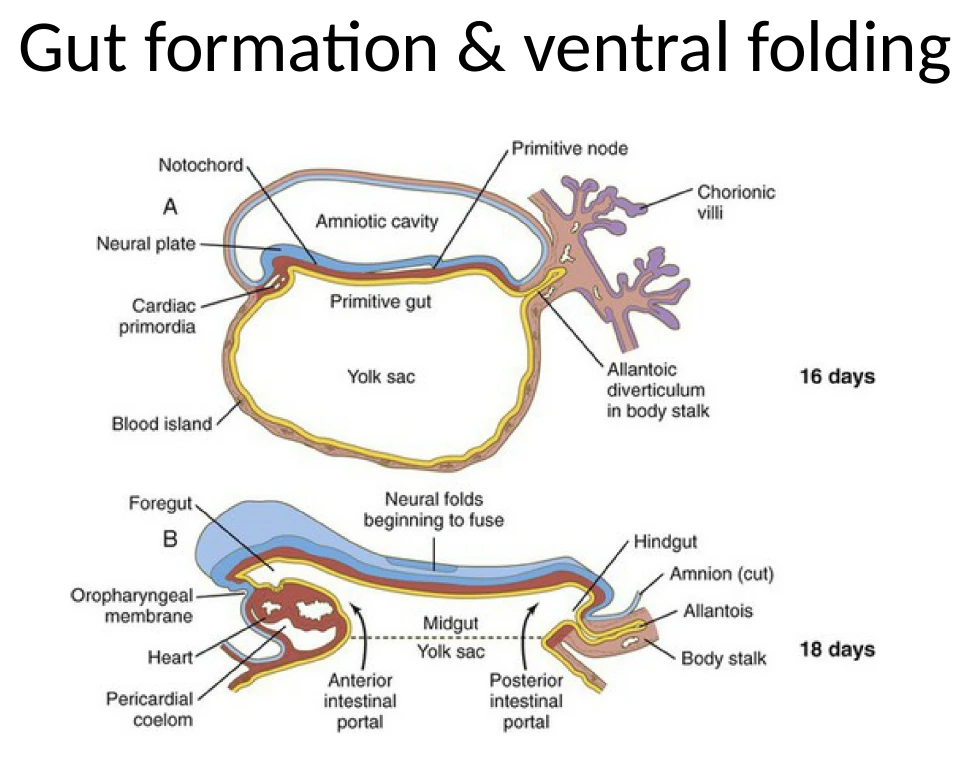
Gut Formation ו־Ventral Folding
אחרי גסטרולציה ותחילת אורגנוגנזה כבר יש:
- מערכת עצבים שמתחילה להיווצר באמצעות נוירולציה.
- סומיטים שנוצרים משני הצדדים של צינור העצבים (Neural tube).
- התחלה של עמוד השדרה ושרירים.
- Endoderm שמונח בצד הוונטרלי.
אבל העובר עדיין פתוח בצד הוונטרלי. כדי ליצור גוף סגור צריך:
- לסגור את ה־Endoderm לצינור - Gut tube.
- לסגור את דופן הגוף ואת חללי הגוף.
- למקם מחדש מבנים כמו הלב.
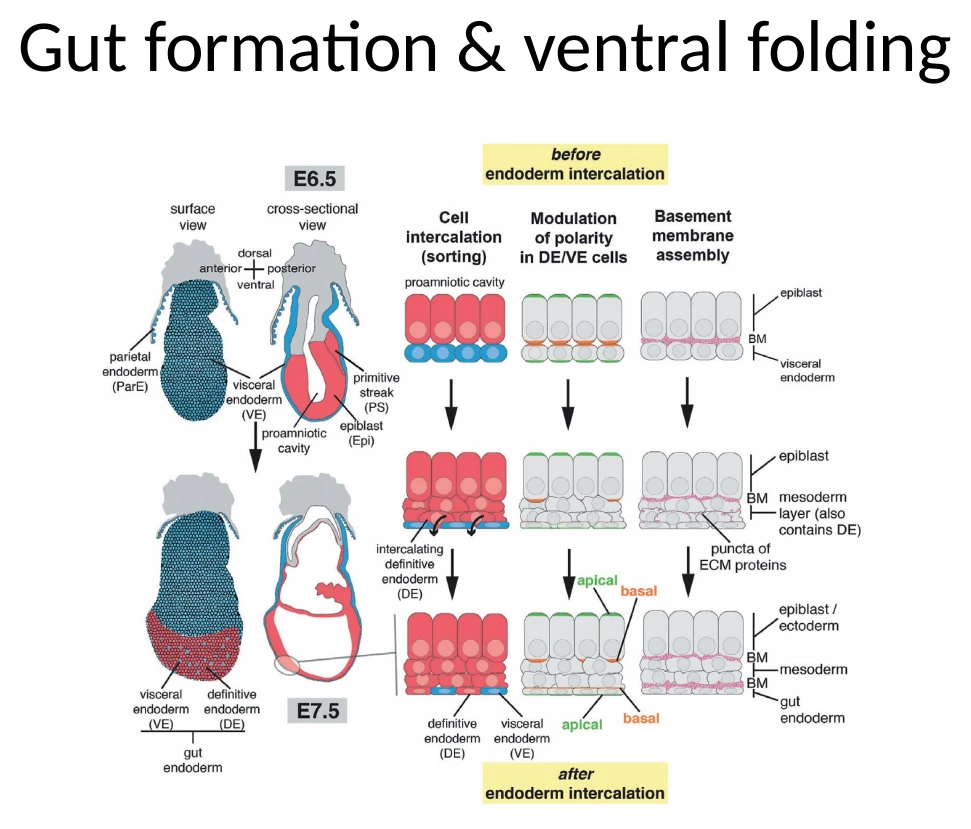
בגסטרולציה, תאים שיצאו דרך ה־Primitive Streak ותרמו ל־Definitive endoderm החליפו בהדרגה את ה־Visceral endoderm באזור העוברי. אבל ה־Definitive endoderm עדיין מחובר להמשך החוץ־עוברי שלו, כלומר ל־Visceral endoderm של ה־yolk sac.
כדי לקבל Gut tube, צריך להפוך את שכבת ה־Endoderm הפתוחה לצינור סגור.
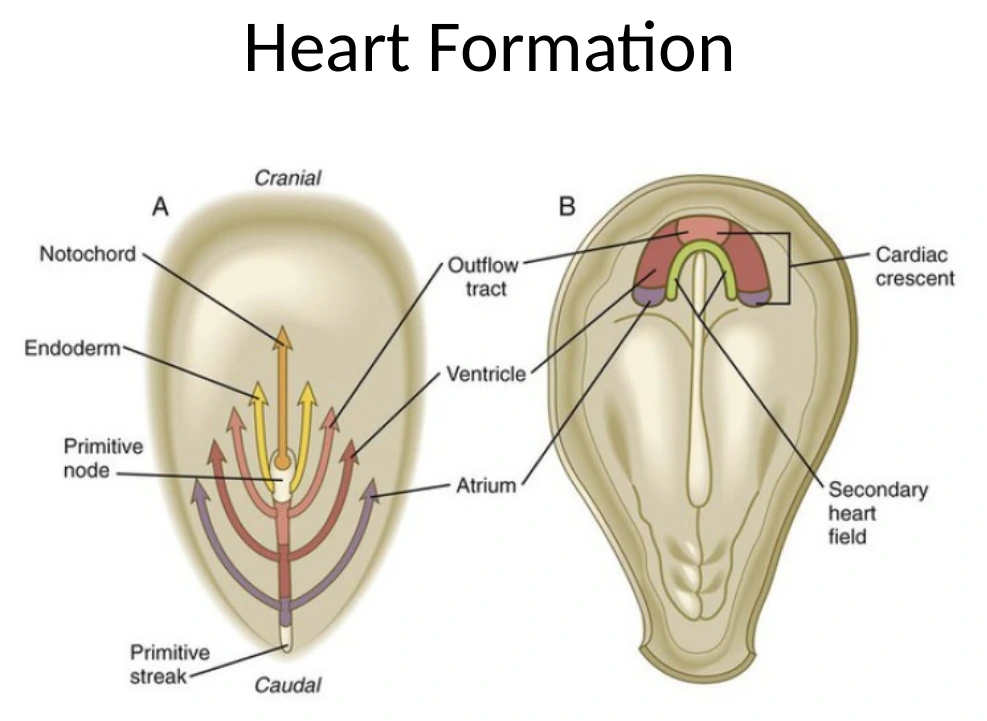
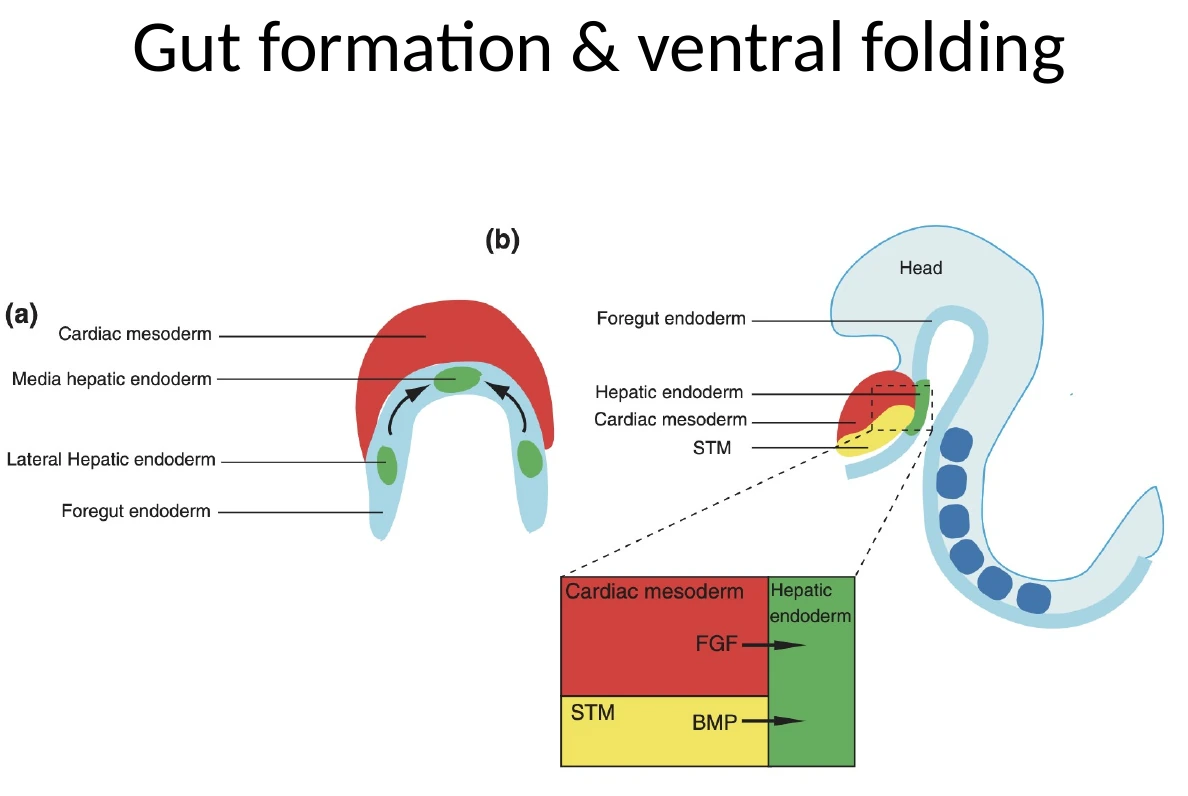
Cardiac Crescent והקשר לקיפול העובר
לפני שנעמיק בקיפול המעיים (gut), צריך לזכור איפה נמצא ה־Cardiac mesoderm.
במהלך הגסטרולציה תאי Cardiac mesoderm יוצאים מה־Primitive Streak, נודדים לטרלית וגם לכיוון אנטריורי/פרוקסימלי, ויוצרים קשת קדמית שנקראת Cardiac Crescent.
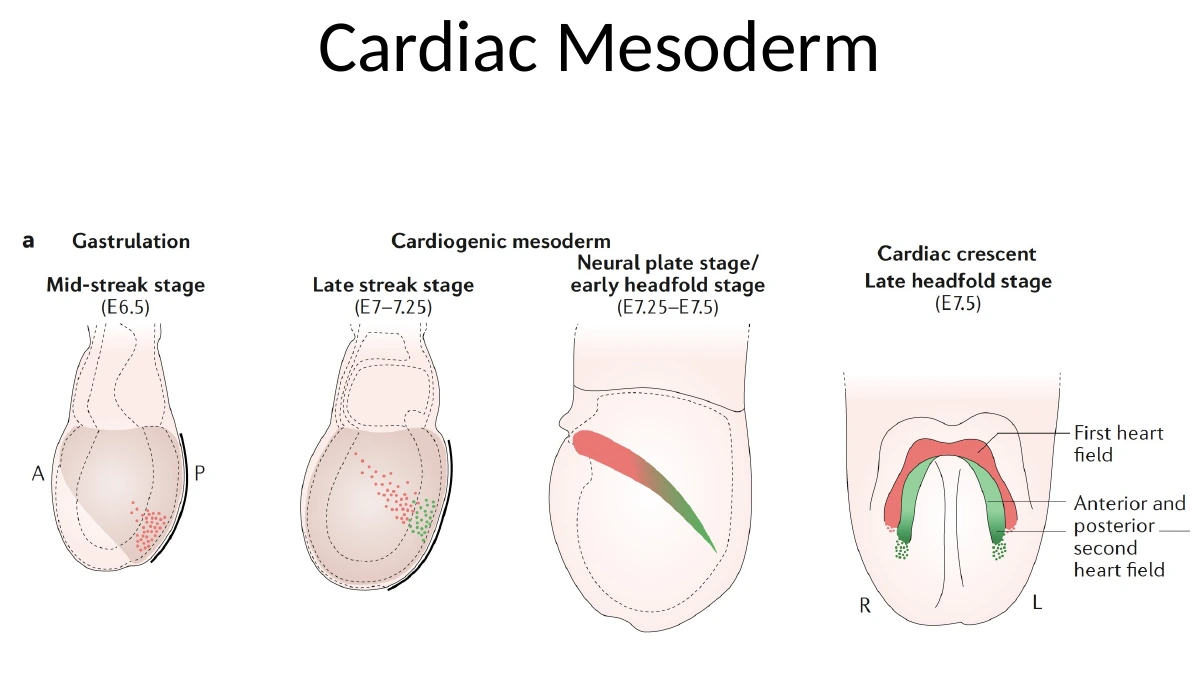 |  |
בשלב מוקדם, ה־Cardiac Crescent נמצא קדמית מאוד - למעשה מעל/לפני האזור שבו מתחיל להתפתח הראש. זה לכאורה מוזר, כי הלב הסופי לא נמצא מעל הראש. אבל בזמן הקיפול הוונטרלי והאנטריורי של העובר, האזור הזה זז למיקום הנכון באזור החזה.
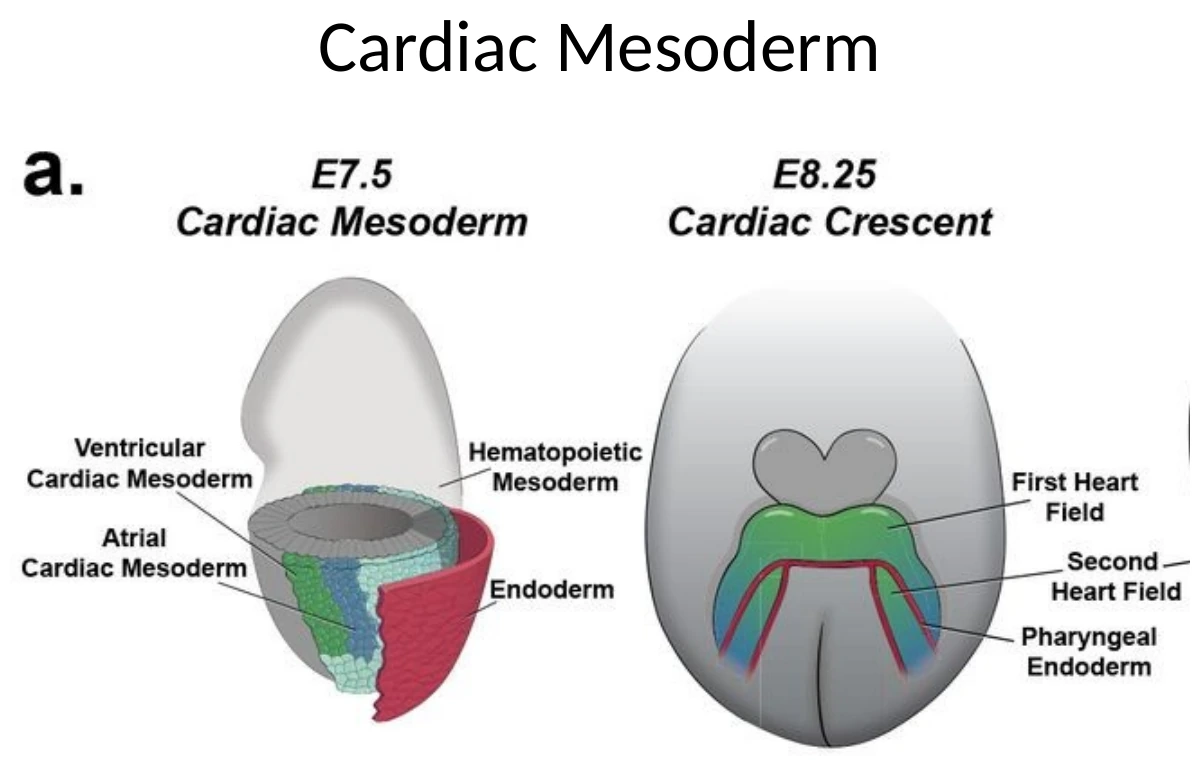
קיפול העובר לסגירת ה־Gut tube משנה את המיקום היחסי של האיברים. הלב מתחיל כאזור קדמי מאוד, ובמהלך הקיפול מגיע לאזור החזה.
Somatopleure, Splanchnopleure וחללי הגוף
בזמן שהעובר מתקדם באורגנוגנזה, ה־Lateral plate mesoderm עובר שינוי חשוב: הוא מתחלק לשתי שכבות אפיתליאליות, וביניהן נוצר חלל.
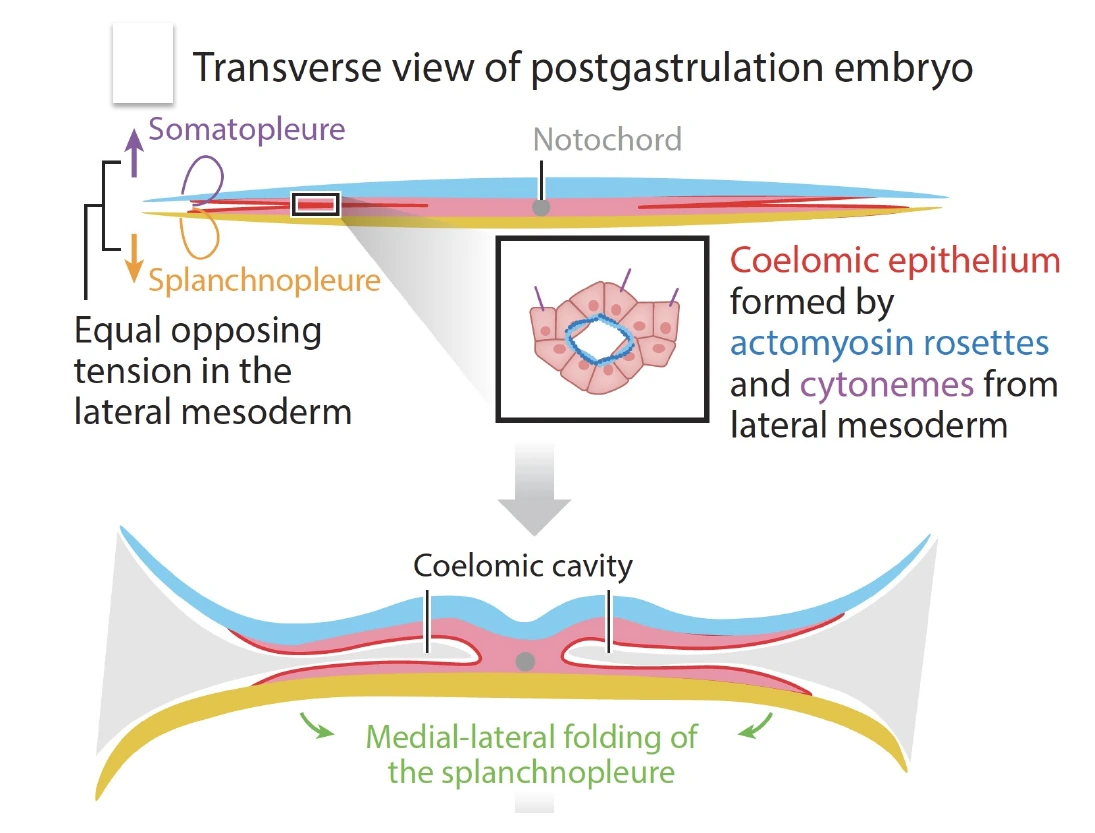
השכבות הן:
| שכבה | מיקום | מה היא צמודה אליו |
|---|---|---|
| Somatopleure | השכבה הלטרלית/דורסלית יותר | צמודה ל־Ectoderm / Surface ectoderm |
| Splanchnopleure | השכבה הפנימית/וונטרלית יותר | צמודה ל־Endoderm |
בין שתי השכבות נוצר חלל - Coelomic cavity. בהמשך יתפתחו ממנו חללי הגוף, למשל חלל החזה וחלל הבטן, לפי המיקום על הציר anterior-posterior.
חשוב לדייק:
- Somatopleure קשורה בעיקר לדופן הגוף.
- Splanchnopleure עוטפת את צינור העיכול ואת האיברים המתפתחים ממנו.
- ה־Gut tube מוקף במזודרם, וקשור לגוף דרך מבנים מזודרמליים כמו dorsal mesentery.
סגירת ה־Gut Tube ודופן הגוף
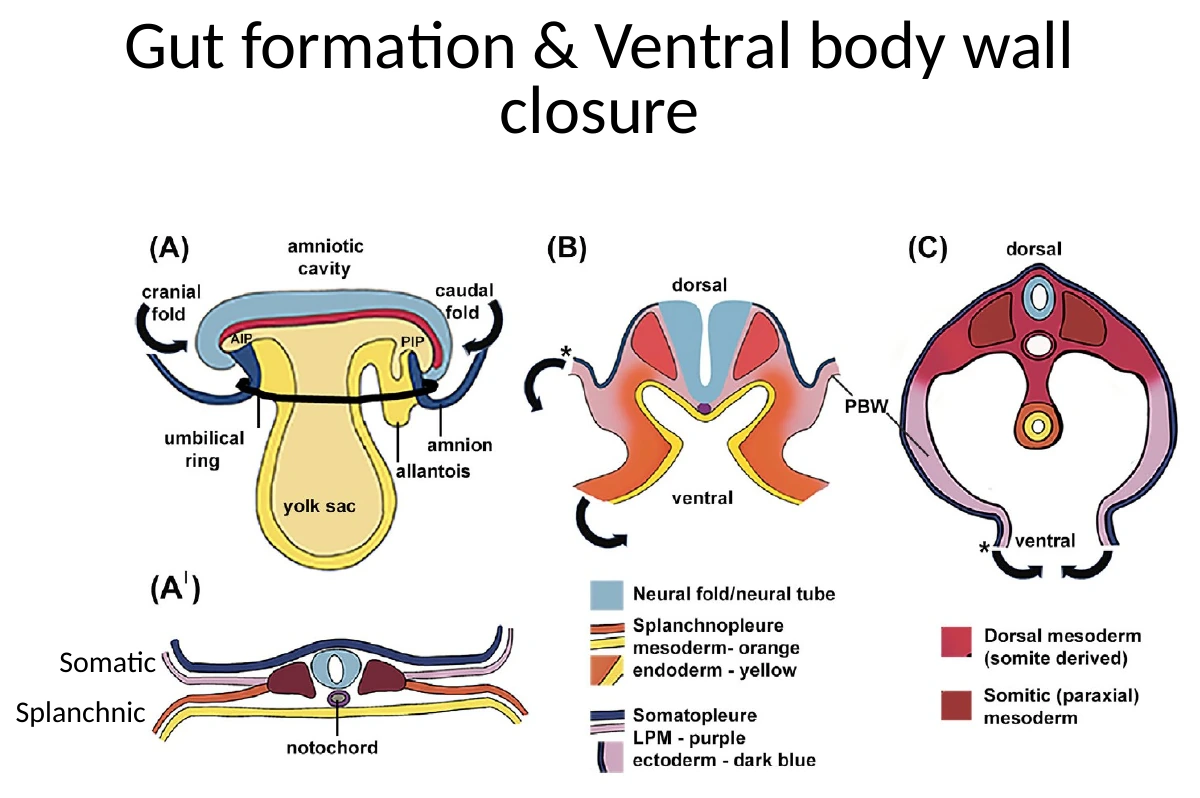
יצירת ה־Gut tube מתרחשת בכמה כיוונים, לא רק בקיפול אחד.
קיפול אנטריורי ופוסטריורי
בצד האנטריורי נוצר Anterior Intestinal Portal (AIP). זהו אזור שבו ה־Endoderm מתחיל להתקפל פנימה וליצור כיס קדמי, שממנו יתפתח ה־Foregut.
בצד הפוסטריורי נוצר Posterior Intestinal Portal (PIP), שמייצר קיפול דומה לכיוון Hindgut.
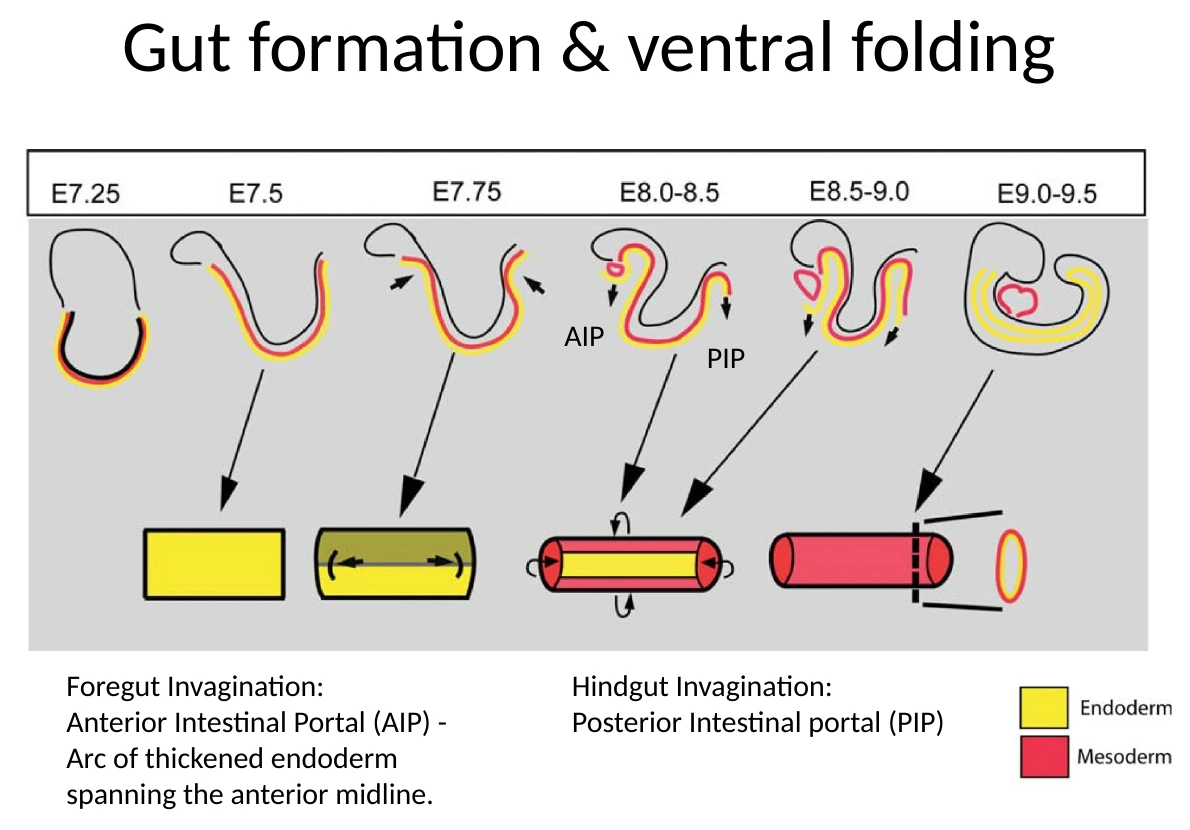
במקביל מתרחשת גם סגירה לטרלית: הצדדים של העובר מתקפלים ונעים לכיוון קו האמצע הוונטרלי.
קיפול לטרלי וסגירת צינור העיכול
בצדדים, ה־Endoderm יחד עם ה־Splanchnopleure מתקפלים ונסגרים ליצירת הצינור. בתהליך הזה, חלק מתאי ה־Splanchnopleure עוברים EMT ומייצרים מזנכימה סביב ה־Endoderm. כך נוצר צינור עיכול שמבפנים הוא Endoderm ומבחוץ עטוף במזודרם.
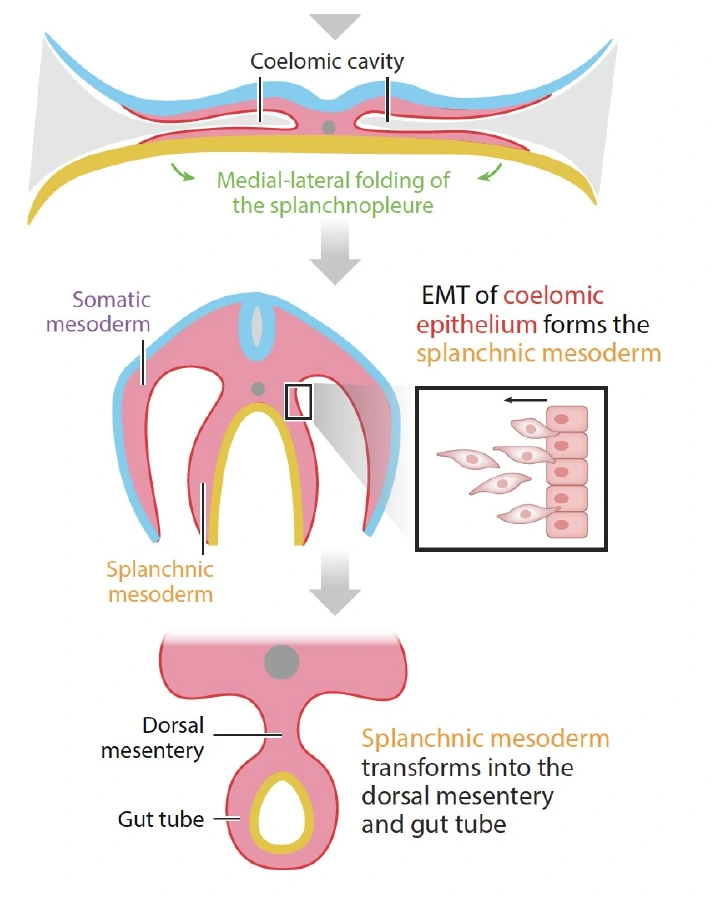
בסוף שלב זה מתקבל:
- צינור פנימי של Endoderm.
- Splanchnic mesoderm סביבו.
- Coelomic cavity סביב האיברים המתפתחים.
- חיבור מזודרמלי לגוף דרך mesentery.
סגירת דופן הגוף
בנפרד מסגירת ה־Gut tube, צריך לסגור גם את דופן הגוף הוונטרלית. כאן מעורבת בעיקר ה־Somatopleure, הצמודה ל־Ectoderm.
יש כאן שני תהליכים מקבילים אך לא זהים:
- סגירת ה־Endoderm לצינור עיכול - קשורה ל־Splanchnopleure.
- סגירת דופן הגוף - קשורה ל־Somatopleure ול־Ectoderm.
הם קורים במקביל ובאותו אזור מרחבי, ולכן קל להתבלבל ביניהם.
טבעת הטבור (Umbilical ring)
במהלך הסגירה נשאר אזור שבו העובר עדיין מחובר לרקמות החוץ־עובריות. זוהי טבעת הטבור, שדרכה יעבור חבל הטבור ומבנים נוספים.
ה־Allantois יתרום בהמשך לחבל הטבור. לכן גם לאחר סגירת דופן הגוף נשאר אזור חיבור מבוקר שדרכו עוברים מבנים חוץ־עובריים.
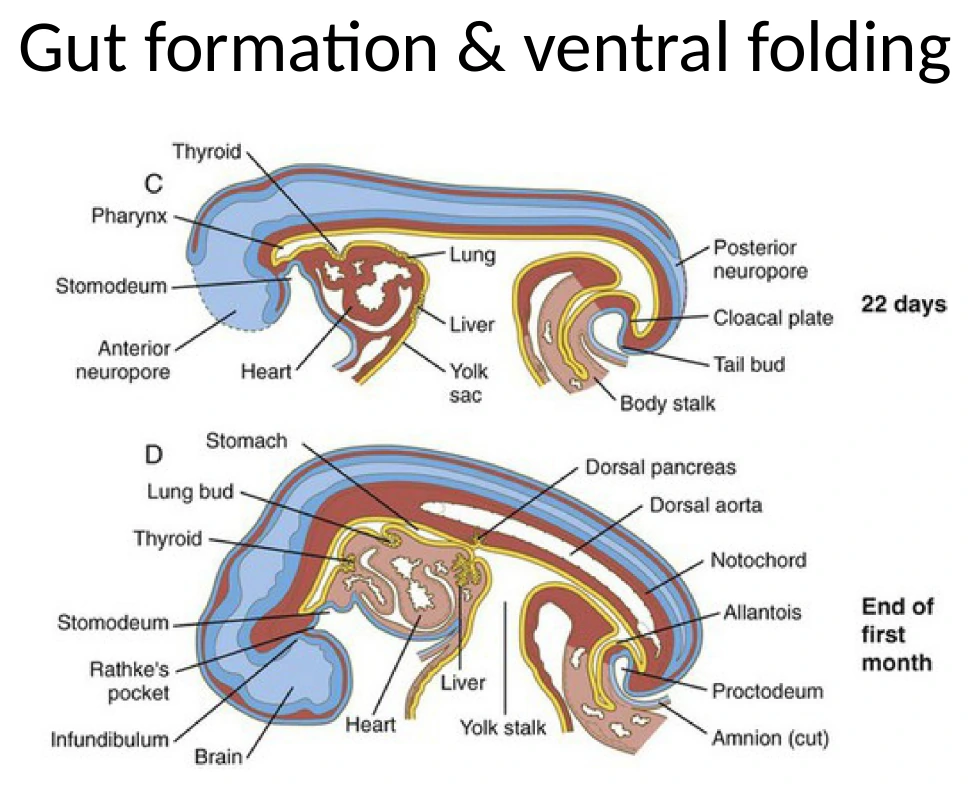
אינדוקציה של איברי אנדודרם לפי גרדיאנטים
אחרי שה־Endoderm נסגר לצינור, אפשר להתחיל להבין איך נוצרים ממנו איברים כמו Thyroid, ריאות, כבד ולבלב. העיקרון הוא אותו עיקרון שחזר לאורך כל הקורס: מיקום על ציר אנטריורי-פוסטריורי וסביבת סיגנלים קובעים גורל.
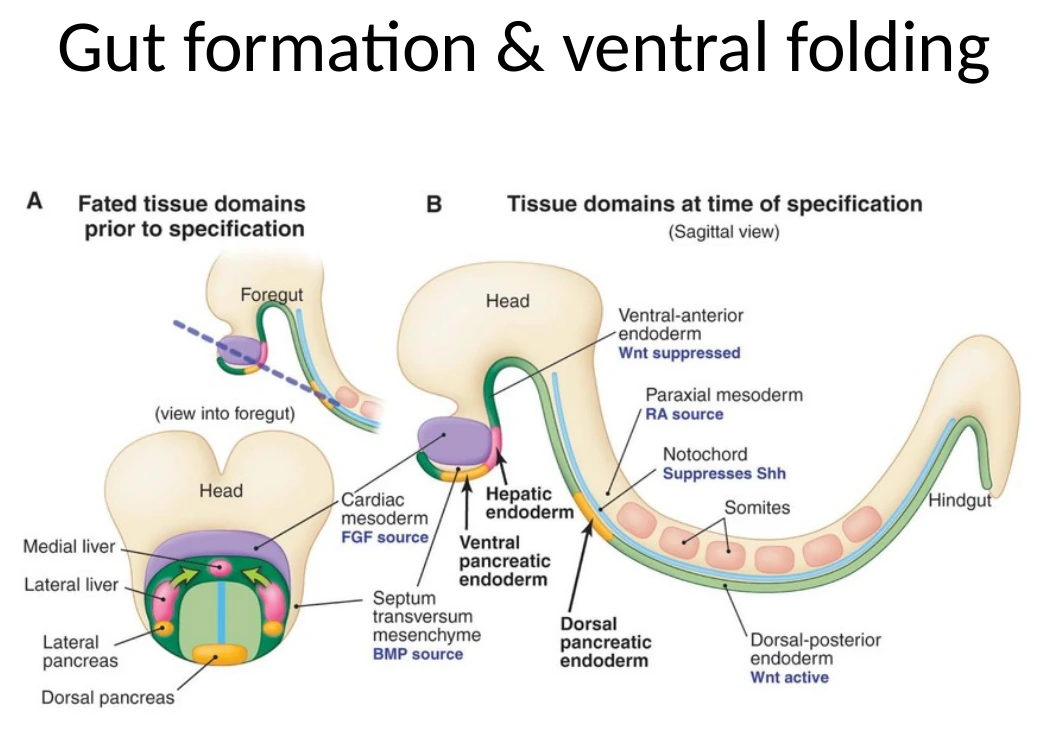
תאי Endoderm מתפתחים תחת קבלת סיגנלים מהמזודרם שסביבם וממבנים סמוכים. לדוגמה:
| איבר / אזור | עיקרון סיגנלי מהשיעור |
|---|---|
| Liver / hepatic endoderm | דורש שילוב של FGF ו־BMP |
| FGF | מגיע בין היתר מה־Cardiac Crescent |
| BMP | מגיע ממזנכימה סמוכה |
| Pancreas | מיקום ודפוסי סיגנלים שונים מאפשרים הבחנה בין dorsal ו־ventral pancreatic endoderm |
| Thyroid / lungs / pancreas | נוצרים כהנצות מאזורים שונים של ה־Gut tube לפי מיקום וסביבה |
 |  |  |
הנקודה החשובה היא הבנת העיקרון:
- לאורך הציר האנטריורי-פוסטריורי נשמרים גרדיאנטים שונים.
- בכל נקודה ה־Endoderm רואה קומבינציה אחרת של סיגנלים.
- המזודרם שסביב ה־Endoderm משתתף באינדוקציה של איברים.
- אורגנוגנזה נובעת מאינטראקציה בין שכבות הנבט
רצף האירועים המרכזי
| שלב | מה קורה |
|---|---|
| Tailbud יוצר FGF8 | נוצר גרדיאנט פוסטריורי-אנטריורי באמצעות RNA decay |
| FGF גבוה | תאי PSM נשמרים במצב לא ממוין |
| FGF יורד באזור אנטריורי | התאים הופכים ל־somite competent |
| Hes7 / cHairy1 / Lfng | גנים אוסילטוריים שיוצרים את השעון |
| Notch | מסנכרן את האוסילציות בין תאים |
| Mesp2 | מופעל באזור המתאים ומתחיל segmentation program |
| סומיט חדש | נוצר אחרי פרגמנטציה ואפיתליאליזציה |
| מספר הסומיטים | נקבע לפי היחס בין Axis elongation לבין קצב השעון |
| Gut formation | Endoderm נסגר לצינור בעזרת AIP, PIP וקיפול לטרלי |
| Ventral body wall closure | Somatopleure ו־Ectoderm סוגרים את דופן הגוף |
| אורגנוגנזה של Endoderm | מיקום וסיגנלים מהמזודרם קובעים איברים כמו כבד, פנקריאס וריאות |