למה צריך בקרת ביטוי גנים?
כמעט לכל תאי הגוף יש אותו גנום, אבל כל תא משתמש בחלק אחר ממנו. נוירון, תא כבד ותא שריר מכילים את אותם גנים, אך מייצרים סט חלבונים ו־RNA שונה. גם אותו תא משנה את הביטוי שלו לפי שלב במחזור התא, מצב עקה, גירוי חיצוני או צורך מטבולי.
בקרת ביטוי גנים פועלת לאורך כל הדרך: מהכרומטין והתחלת השעתוק, דרך עיבוד ה־RNA, יציאה מהגרעין, תרגום, פירוק mRNA, פירוק חלבונים והפעלה או השתקה של חלבונים קיימים.
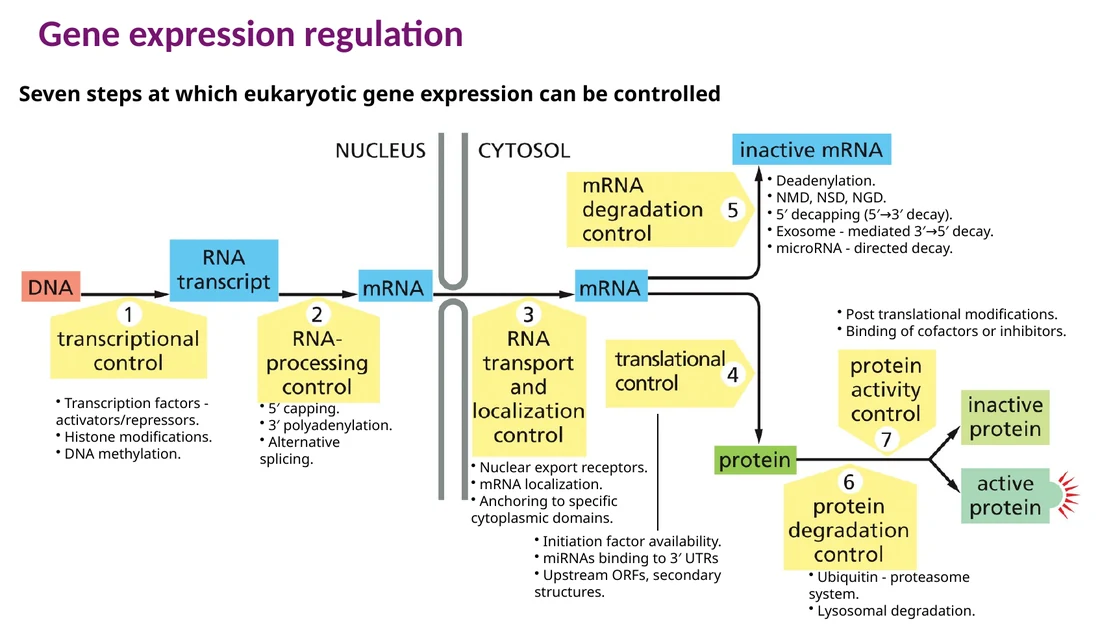
Transcriptional control - בחירה אילו גנים ישועתקו וכמה mRNA ייווצר.
RNA processing control - קאפינג, polyadenylation, splicing ו־alternative splicing.
RNA transport/localization control - יציאה מהגרעין והבאת mRNA לאזור המתאים בתא.
Translational control - שליטה בכמות החלבון שתיווצר מכל mRNA.
mRNA degradation control - קביעת זמן החיים של mRNA.
Protein degradation/activity control - פירוק חלבונים או הפעלה שלהם אחרי שכבר נוצרו.
בקרת שעתוק: פקטורי שעתוק, enhancers ו־repressors
פקטורי שעתוק רגולטוריים נקשרים לרצפי DNA קצרים, בדרך כלל באורך של כ־5-12 זוגות בסיסים. הרצפים האלה נקראים cis-regulatory sequences כי הם נמצאים על אותו כרומוזום כמו הגן שהם מבקרים.
פקטור שעתוק יכול להיקשר ל־enhancer ולקדם שעתוק, או ל־silencer ולדכא שעתוק. הפעולה מתבצעת דרך קישור ישיר או עקיף ל־general transcription factors, ל־Mediator, ל־cohesin, ל־chromatin remodelers ולחלבונים שמוסיפים או מסירים מודיפיקציות מהיסטונים.
פקטורי שעתוק עובדים לרוב כצוות
גן אאוקריוטי טיפוסי מבוקר על ידי כמה רצפים רגולטוריים וכמה פקטורי שעתוק. כל פקטור תורם חלק מהמידע: סוג התא, מצב מחזור התא, גירוי חיצוני, זמינות נוטריינטים או עקה. השילוב ביניהם קובע את רמת הביטוי.
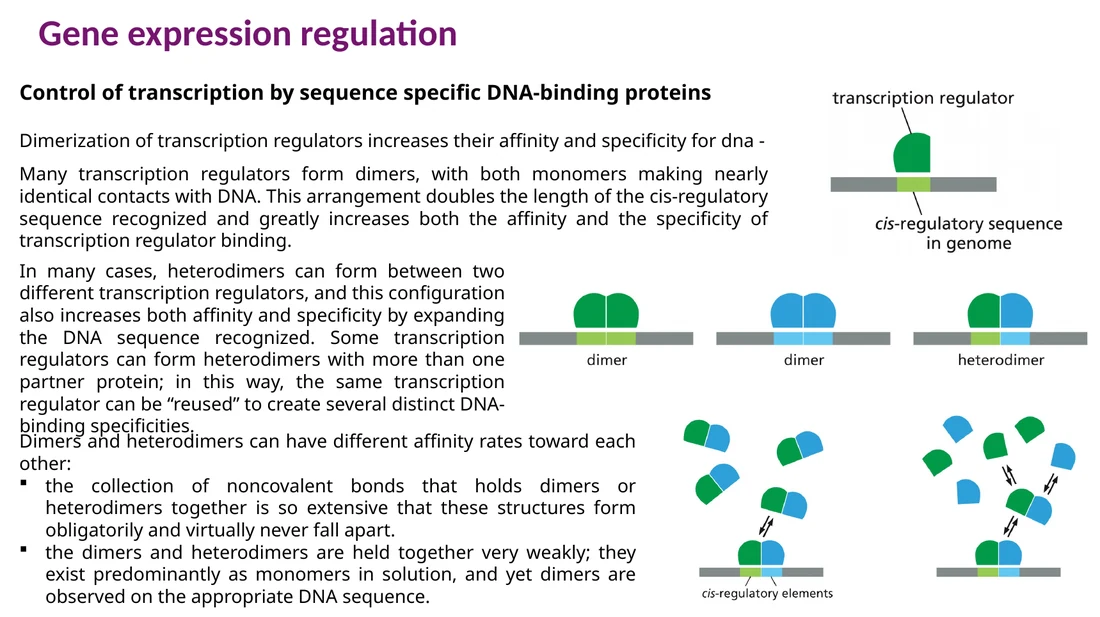
דימריזציה מעלה גם אפיניות וגם ספציפיות. כאשר שני מונומרים נקשרים יחד, הם מזהים רצף ארוך יותר על ה־DNA. הטרודימר מוסיף עוד שכבה של ספציפיות, כי אותו פקטור שעתוק יכול לעבוד עם שותפים שונים ולכוון לגנים שונים.
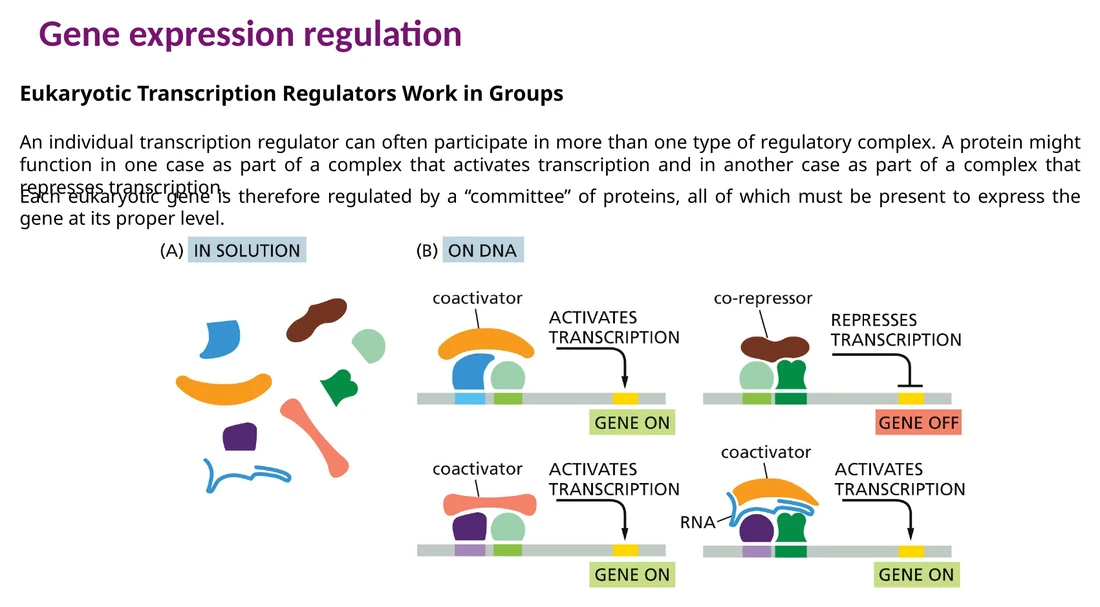
למבחן: פקטור שעתוק אחד יכול להפעיל גן בהקשר אחד ולדכא גן בהקשר אחר. הפעולה שלו תלויה בשותפים שנקשרים אליו, במיקום שלו בכרומטין וב־cofactors שהוא מגייס.
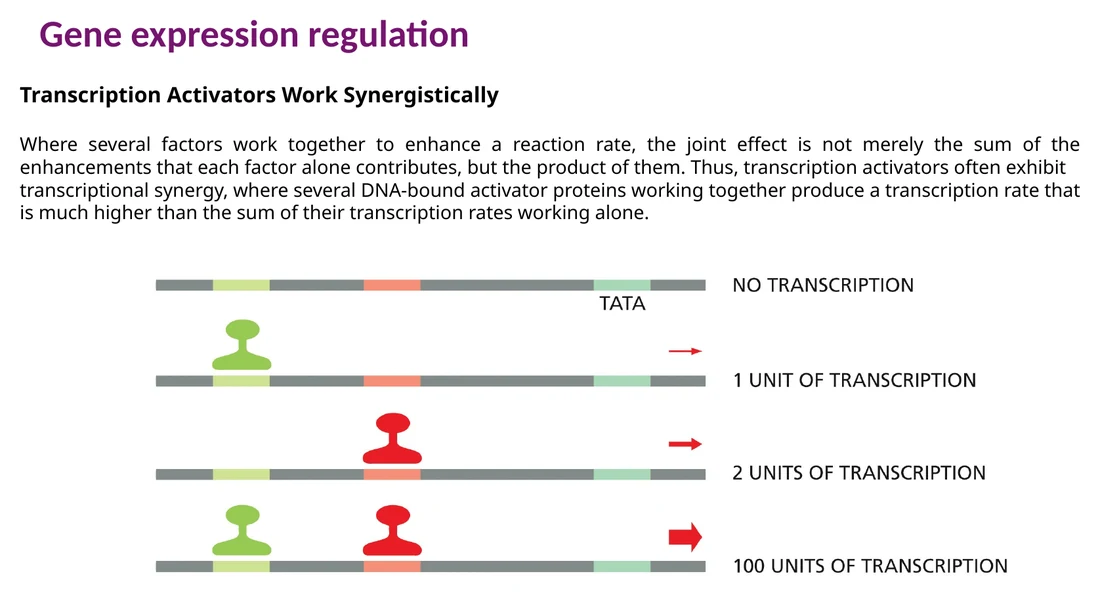
סינרגיה בין activators
כאשר שני activators נקשרים לאותו אזור רגולטורי, השפעתם יכולה להיות חזקה בהרבה מהתרומה של כל אחד בנפרד. זה מאפשר לתא ליצור תגובה מדורגת: מעט שעתוק עם פקטור אחד, הרבה שעתוק כאשר שני פקטורים או יותר נמצאים יחד.
איך repressors מדכאים שעתוק?
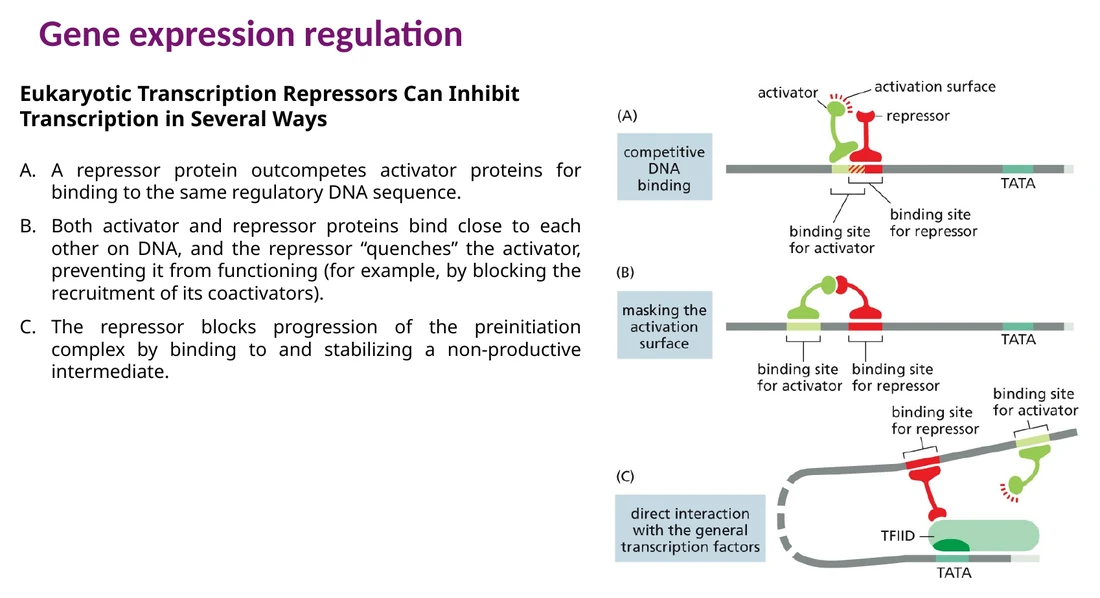
Repressor יכול לדכא שעתוק בכמה דרכים:
תחרות על אותו אתר DNA - ה־repressor תופס את הרצף שה־activator צריך.
Quenching - ה־repressor נקשר ליד ה־activator או אליו ומפריע לו לגייס coactivators.
ייצוב קומפלקס לא פעיל - ה־repressor עוצר את ההתקדמות של pre-initiation complex.
Insulators, CTCF וארגון תלת־ממדי של הכרומטין
Enhancer יכול לפעול ממרחק גדול. כדי שה־enhancer יגיע לגן הנכון, הכרומטין מאורגן בלופים ובשכונות רגולטוריות. Insulators מגבילים את תחום הפעולה של enhancer, וביונקים החלבון המרכזי בהקשר הזה הוא CTCF.
CTCF נקשר לאתרים ייעודיים, מייצב לופים של כרומטין ומסייע להפרדה בין אזורים רגולטוריים. כך enhancer שנמצא בשכונה אחת משפיע על הפרומוטר המתאים בתוך אותה שכונה, ופחות על גנים שנמצאים מעבר לגבול הכרומטיני.
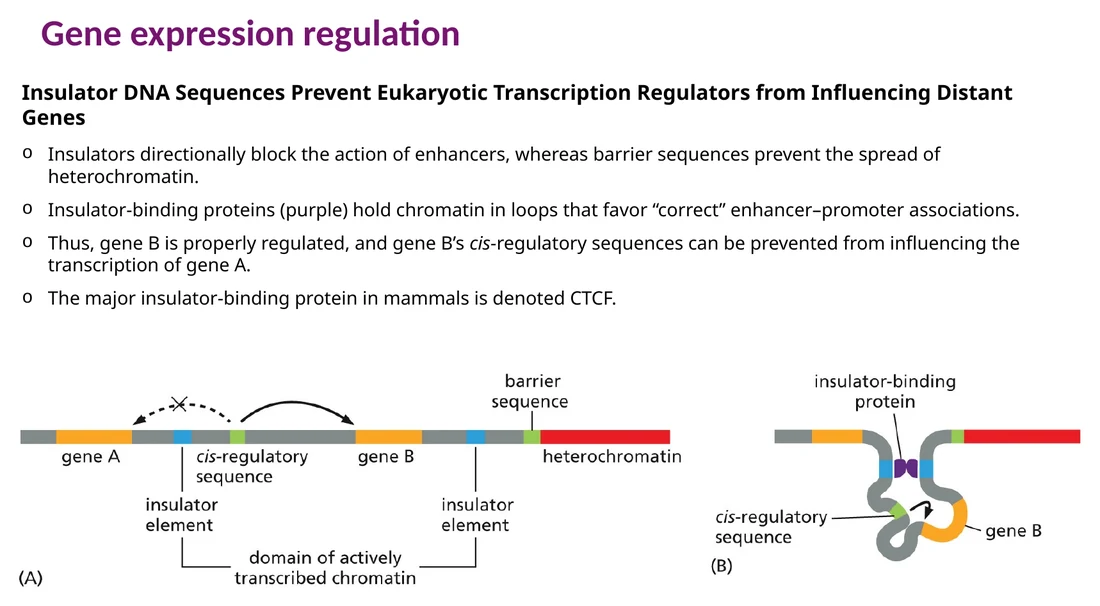
למבחן: Insulator הוא חוצץ רגולטורי. הוא מארגן את הכרומטין כך ש־enhancer יפעל על הגן המתאים ולא על גן סמוך במקרה.
איך פקטור שעתוק מזהה רצף DNA?
פקטור שעתוק לא צריך לפתוח את ה־double helix כדי לזהות רצף. הוא קורא את פני השטח של ה־DNA דרך ה־major groove וה־minor groove. לכל זוג בסיסים יש דגם מרחבי של תורמי קשרי מימן, מקבלי קשרי מימן, קבוצות מתיל ואזורים הידרופוביים. הדגם הזה משתנה לפי הזהות והכיוון של זוג הבסיסים.
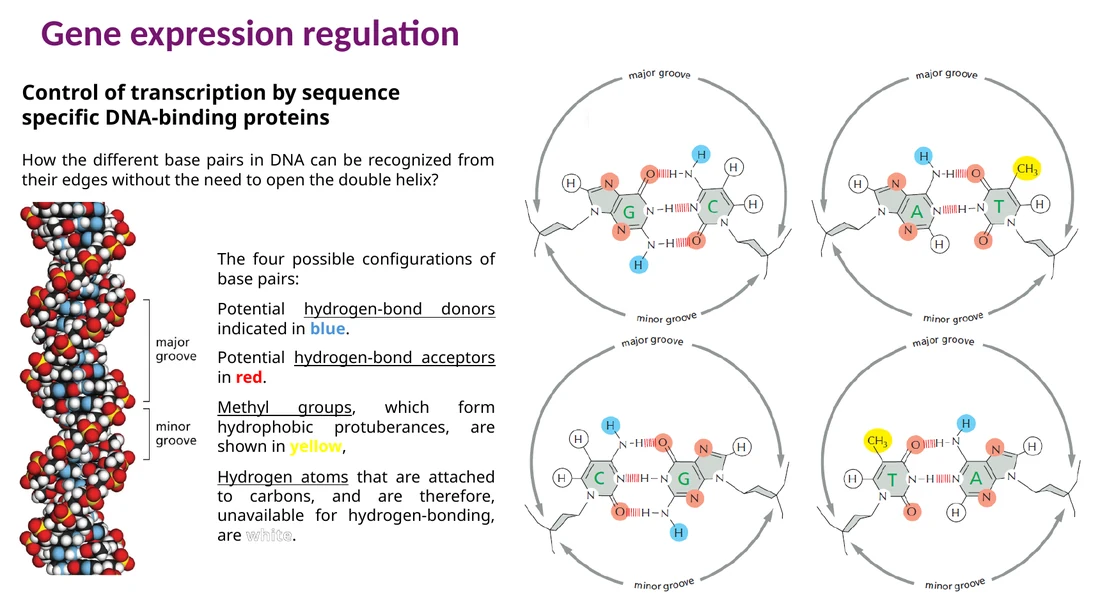
פקטור השעתוק יוצר סדרה של קשרים עם ה־DNA: קשרי מימן עם בסיסים, קשרים יוניים עם שלד הסוכר-פוספט ואינטראקציות הידרופוביות עם קבוצות מתיל. שינוי של בסיס יחיד יכול לשנות את הדגם המרחבי ולפגוע בקישור.
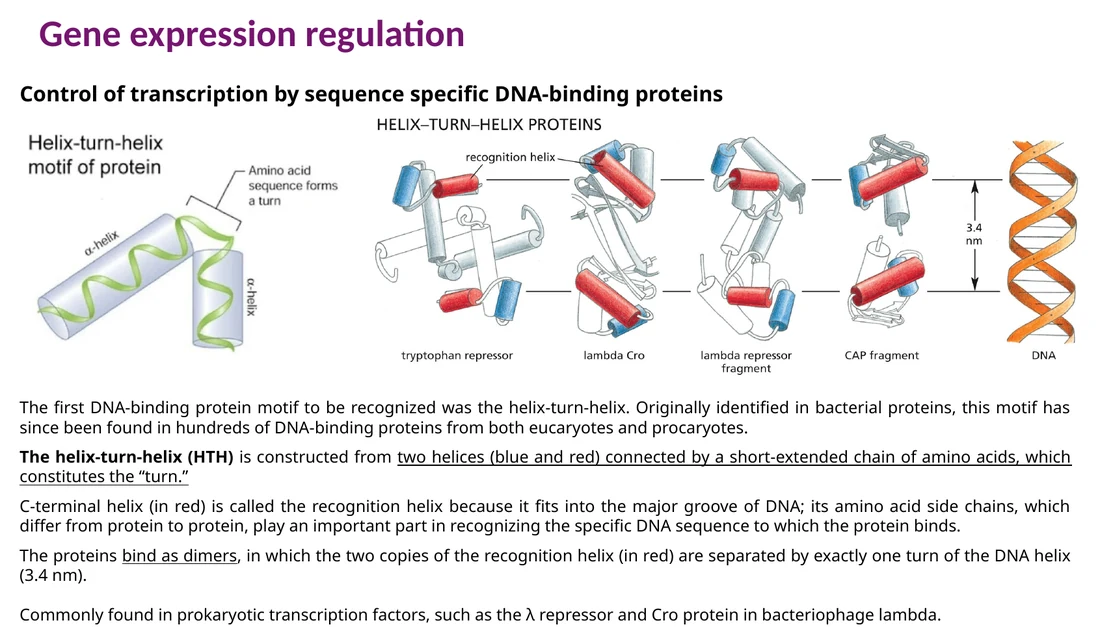
מוטיבים קושרי DNA בפקטורי שעתוק
מוטיב קושר DNA הוא אזור מבני בחלבון שמזהה את ה־DNA. רוב המוטיבים נקשרים ל־major groove, כי שם ההבדלים בין רצפים חשופים בצורה ברורה יותר.
Helix-turn-helix
Helix-turn-helix מורכב משני alpha helices המחוברים ב־turn קצר. ה־recognition helix נכנס ל־major groove ומבצע את הזיהוי הרצפי. בחלבונים רבים המוטיב עובד כדימר, ושני ה־recognition helices מופרדים בערך בסיבוב אחד של DNA, כ־3.4 ננומטר.
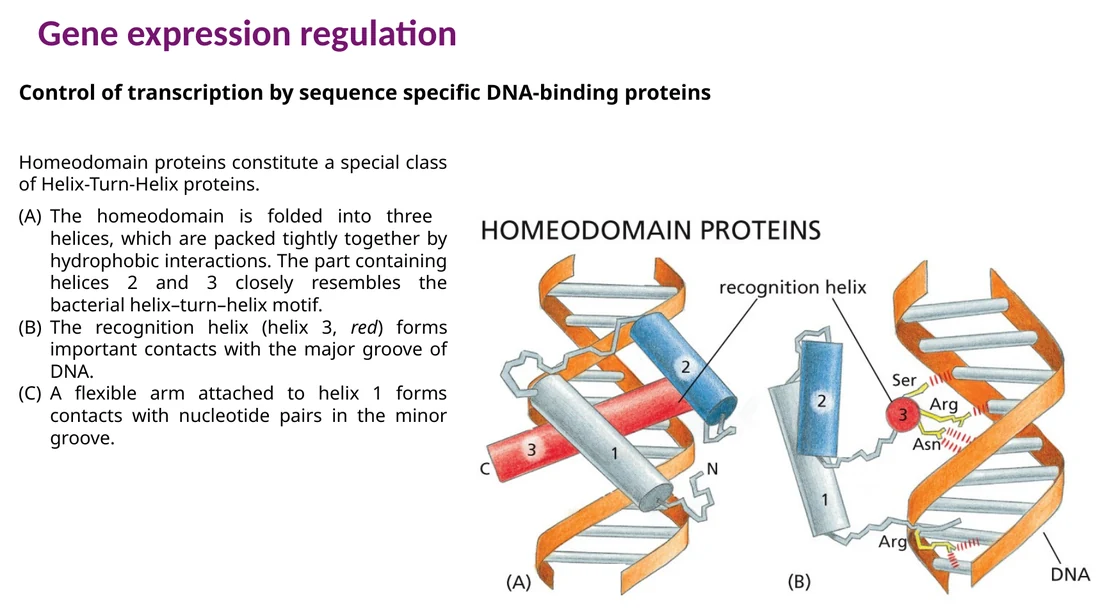
Homeodomain וחלבוני Hox
Homeodomain הוא גרסה אאוקריוטית מפותחת של helix-turn-helix. יש בו שלושה helices. ה־Helix 3 יוצר קשרים עם ה־major groove, וזרוע גמישה נוספת יכולה ליצור קשרים עם ה־minor groove.
חלבוני Hox הם דוגמה חשובה ל־homeodomain transcription factors. הם מבקרים דפוסי התפתחות לאורך הציר anterior-posterior, ולכן שינוי במיקום הביטוי שלהם יכול לשנות זהות של אזור בגוף.
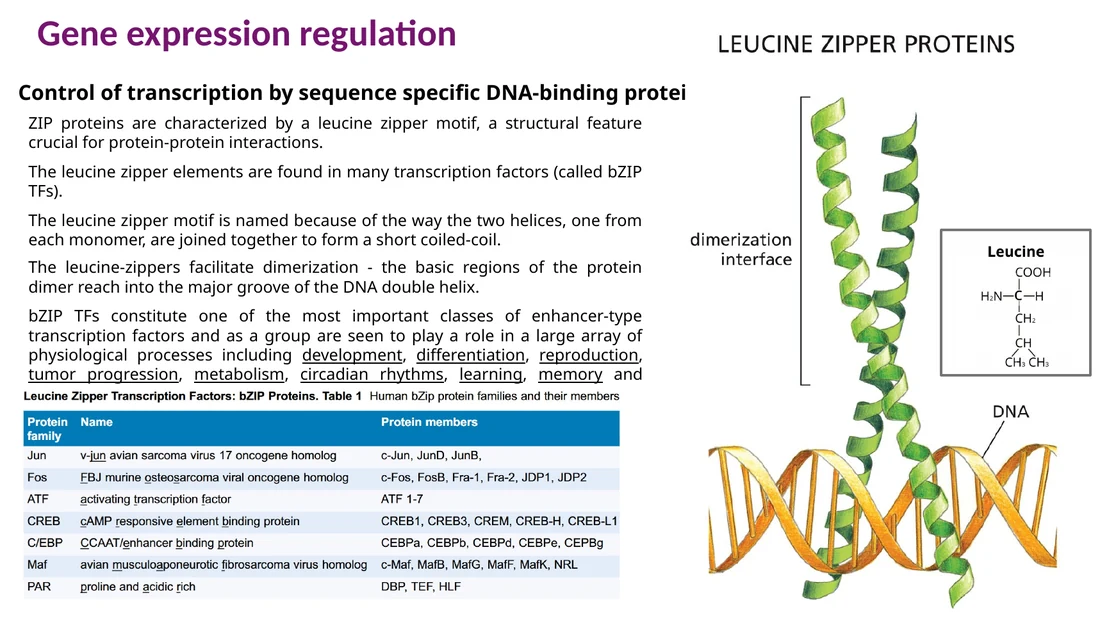
Leucine zipper
ב־leucine zipper שני alpha helices יוצרים coiled-coil. ה־Leucine מופיע בערך כל שבע חומצות אמינו באותו צד של ההליקס, וכך נוצרת דימריזציה דרך צדדים הידרופוביים. אזור basic סמוך נקשר ל־DNA.
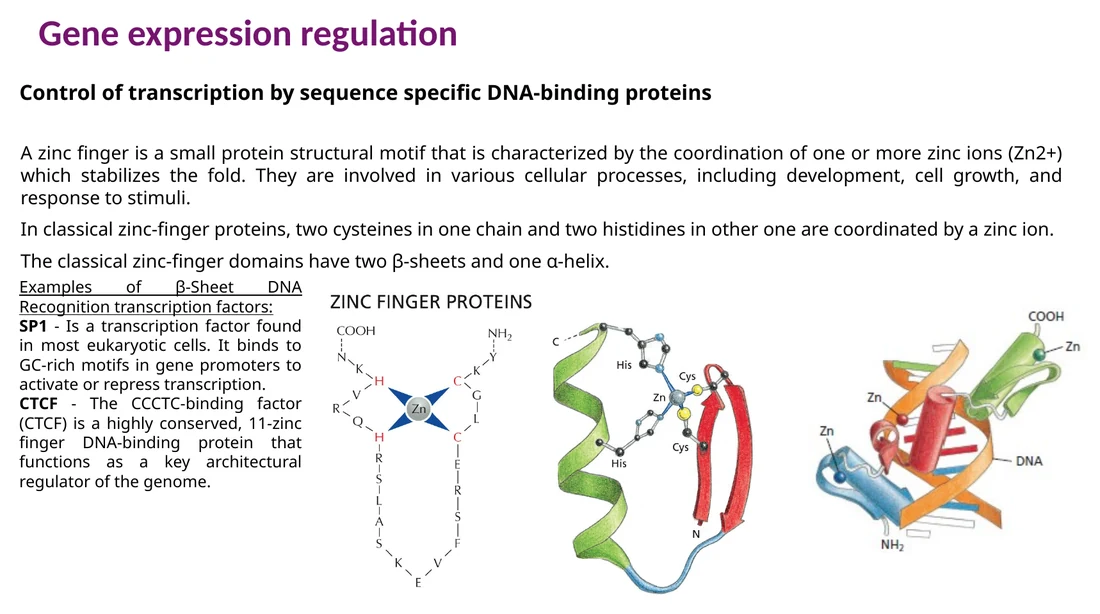
Zinc finger
ב־zinc finger, יון אבץ מייצב קיפול קטן של החלבון בעזרת Cys ו־His. חלבונים רבים מכילים כמה zinc fingers ברצף, וכל finger תורם לזיהוי חלק מהרצף. SP1 הוא דוגמה לפקטור שעתוק עם zinc fingers שמזהה GC-rich motifs.
β-sheet recognition ו־TBP
TBP נקשר ל־TATA box דרך ה־minor groove. הקישור מכופף את ה־DNA בזווית גדולה ומסייע לפתיחת האזור לקראת הרכבת קומפלקס השעתוק.
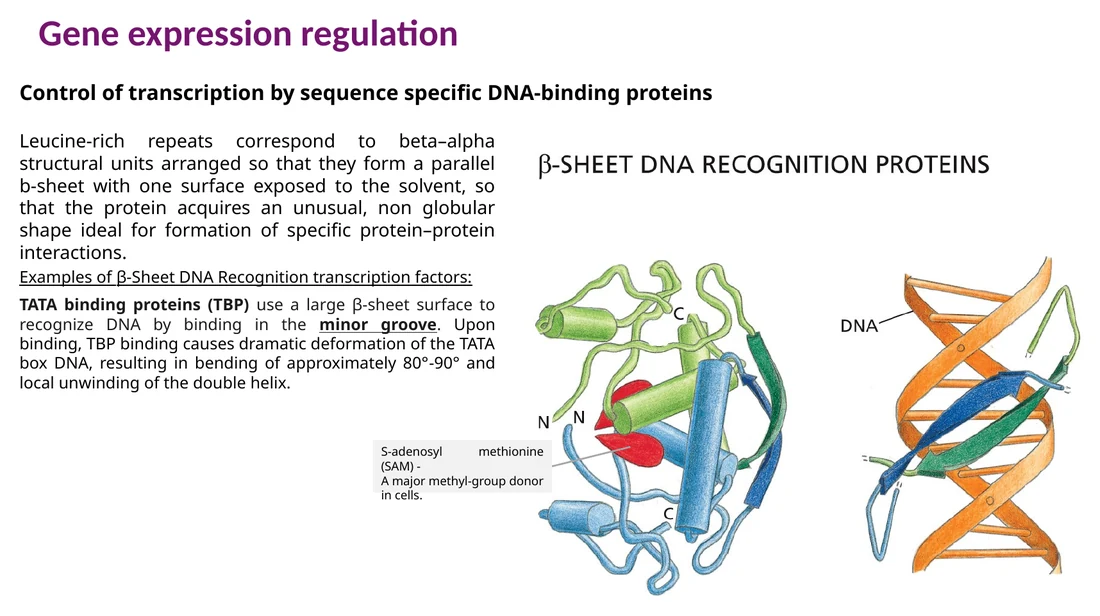
למבחן: רוב המוטיבים נקשרים ל־major groove. TBP הוא דוגמה בולטת לקישור ל־minor groove, והקישור שלו מכופף את ה־DNA באזור ה־TATA box.
בקרת כרומטין: nucleosomes, remodelers ומודיפיקציות היסטונים
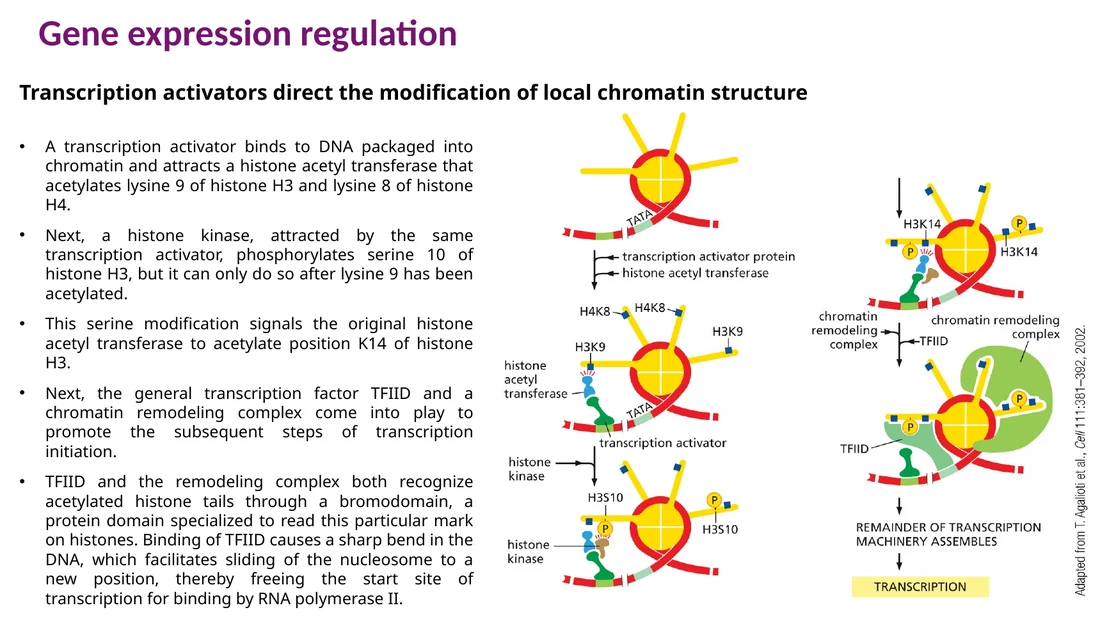
ה־DNA ארוז סביב נוקלאוזומים. האריזה הזאת מקשה על פקטורי שעתוק ו־RNA polymerase להגיע לרצפים רגולטוריים. פתיחת כרומטין וסגירת כרומטין הן חלק מרכזי מבקרת שעתוק.
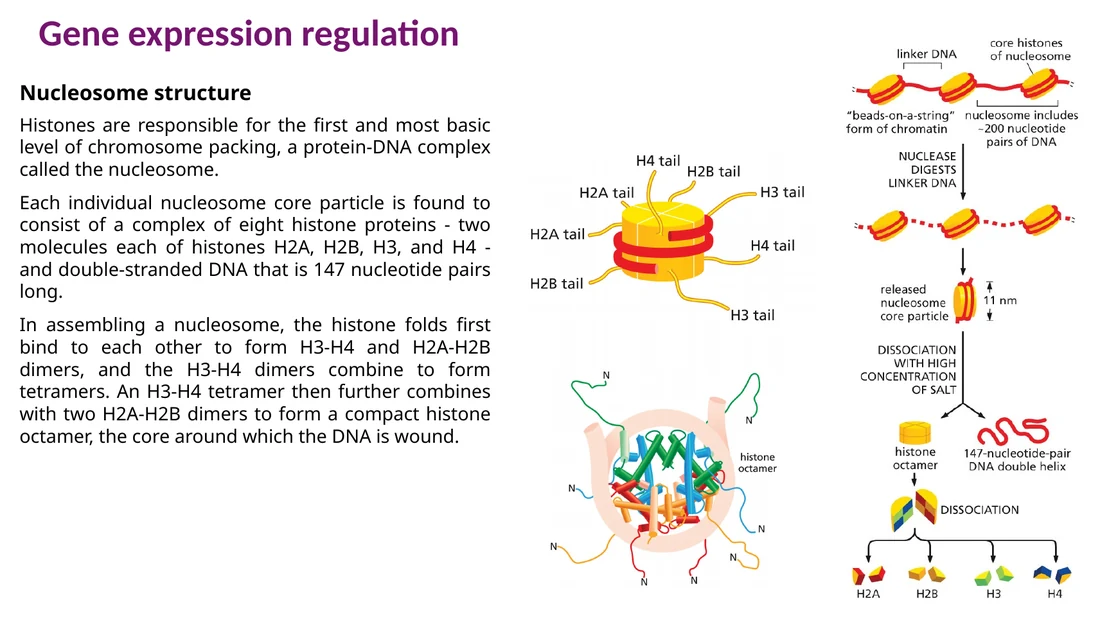
Chromatin remodelers
Chromatin remodelers משתמשים ב־ATP כדי להזיז נוקלאוזומים, לפנות אותם מה־DNA או להחליף תתי-יחידות היסטוניות. כך הם חושפים פרומוטור, enhancer או TATA box ומאפשרים קישור של פקטורי שעתוק כלליים.
Writers, erasers, readers
חלבונים שמבקרים היסטונים מתחלקים לשלוש קבוצות שימושיות:
Writers מוסיפים מודיפיקציות, למשל acetyltransferases או methyltransferases.
Erasers מסירים מודיפיקציות, למשל deacetylases או demethylases.
Readers מזהים מודיפיקציה קיימת ומגייסים חלבונים נוספים.
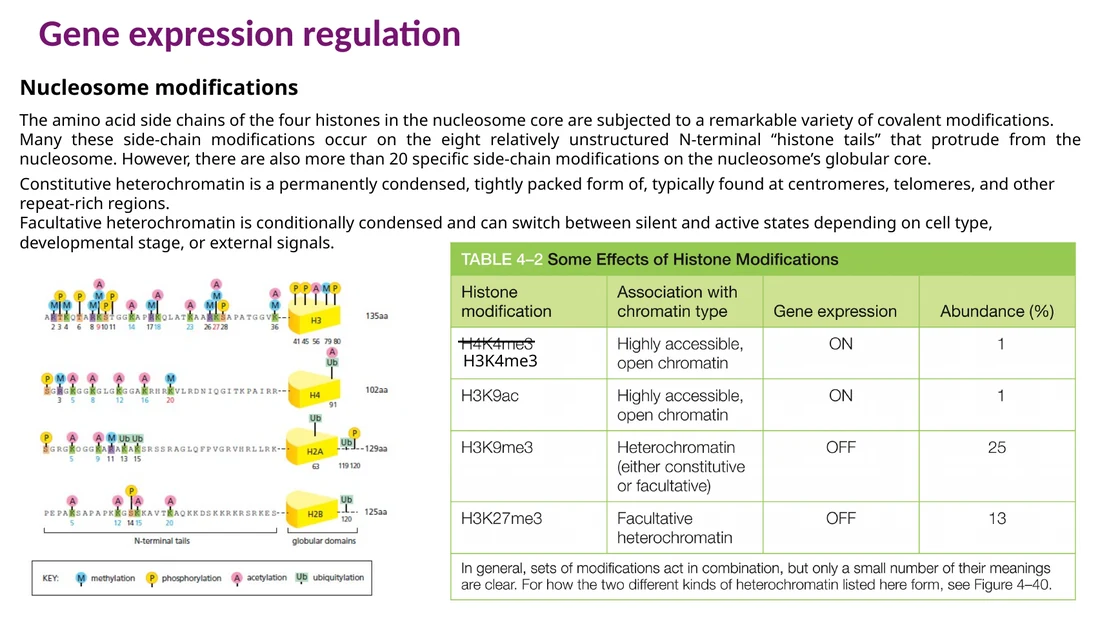
Acetylation של lysine בזנבות היסטונים מחלישה את האינטראקציה בין ההיסטונים ל־DNA ומקלה על פתיחת הכרומטין. Methylation תלויה בהקשר: למשל H3K4me3 מאפיין פרומוטורים פעילים, בעוד H3K9me3 ו־H3K27me3 מאפיינים כרומטין מושתק יותר.
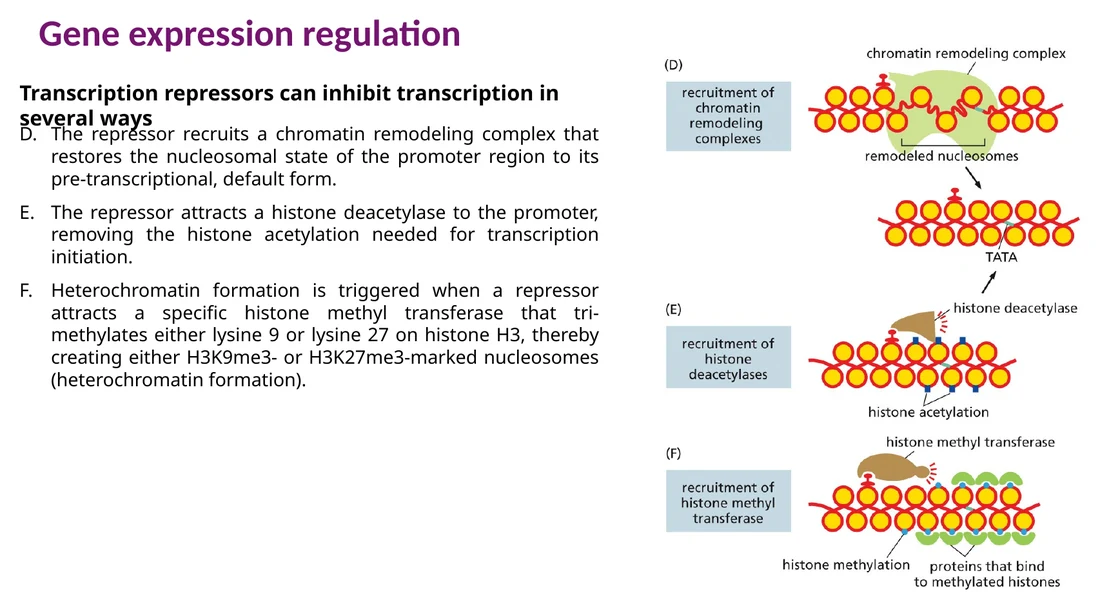
Repressor יכול לגייס remodelers שסוגרים את האזור, HDACs שמסירים acetylation, או methyltransferases שמייצרות סימנים רפרסיביים כמו H3K9me3 ו־H3K27me3.
Alternative splicing: איך אותו pre-mRNA יוצר כמה תוצרים?
ב־splicing קנוני כל intron יוצא וה־exons מתחברים ברצף אחד. ב־alternative splicing, התא בוחר קומבינציות שונות של exons ו־splice sites. כך גן אחד יכול ליצור כמה mRNAs וכמה איזופורמים של חלבון.
חוזק splice site
ה־spliceosome מזהה שלושה אזורים חשובים: 5′ splice site, branch point ו־3′ splice site. ככל שהרצף דומה יותר לקונצנזוס, ה־splice site חזק יותר. רצף חלש דורש סיוע של חלבונים רגולטוריים כדי שה־spliceosome יזהה אותו היטב.
SR proteins ו־hnRNPs
SR proteins נקשרים ל־splicing enhancers ומחזקים שימוש ב־splice site סמוך.
hnRNPs נקשרים ל־splicing silencers ומפחיתים הכרה של splice site סמוך.
האזורים הרגולטוריים יכולים לשבת בתוך exon או בתוך intron:
ESE - Exonic Splicing Enhancer ESS - Exonic Splicing Silencer ISE - Intronic Splicing Enhancer ISS - Intronic Splicing Silencer
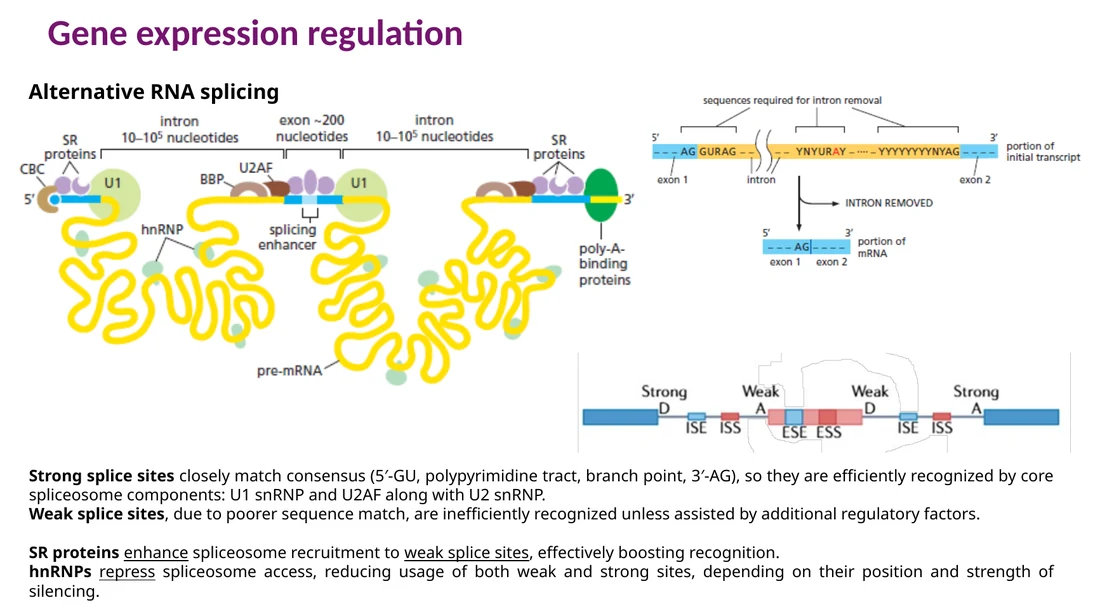
למבחן: Alternative splicing נשען על שני דברים יחד: חוזק הרצף של ה־splice sites וחלבונים רגולטוריים שנקשרים ל־enhancers או silencers.
סוגים נפוצים של alternative splicing

| סוג | מה קורה בפועל |
|---|---|
| Exon skipping | exon מסוים יוצא יחד עם האינטרונים שסביבו. |
| Intron retention | intron נשאר ב־mRNA. לעיתים ה־RNA נשאר בגרעין, ולעיתים התוצר משתתף ברגולציה. |
| Alternative 5′ splice site | התחלת האינטרון נבחרת ממיקום אחר. |
| Alternative 3′ splice site | סוף האינטרון נבחר ממיקום אחר. |
| Mutually exclusive exons | exon אחד נכלל, והאחר יוצא. |
| Alternative promoter / poly-A site | התחלה או סיום שונים של ה־transcript. |
| Back-splicing / circular RNA | exon מתחבר לעצמו או לאזור upstream ונוצר RNA מעגלי. |
בקרת מיקום mRNA בתא
mRNA יכול לצאת מהגרעין ולעבור למיקום מוגדר בציטופלזמה לפני תרגום יעיל. הסיגנלים לכך נמצאים לרוב ב־3′ UTR ונקראים לעיתים zipcode elements. RNA-binding proteins נקשרים אליהם, יוצרים mRNP, מחברים אותו למנועים של cytoskeleton ומביאים את ה־mRNA לאזור שבו החלבון נדרש.
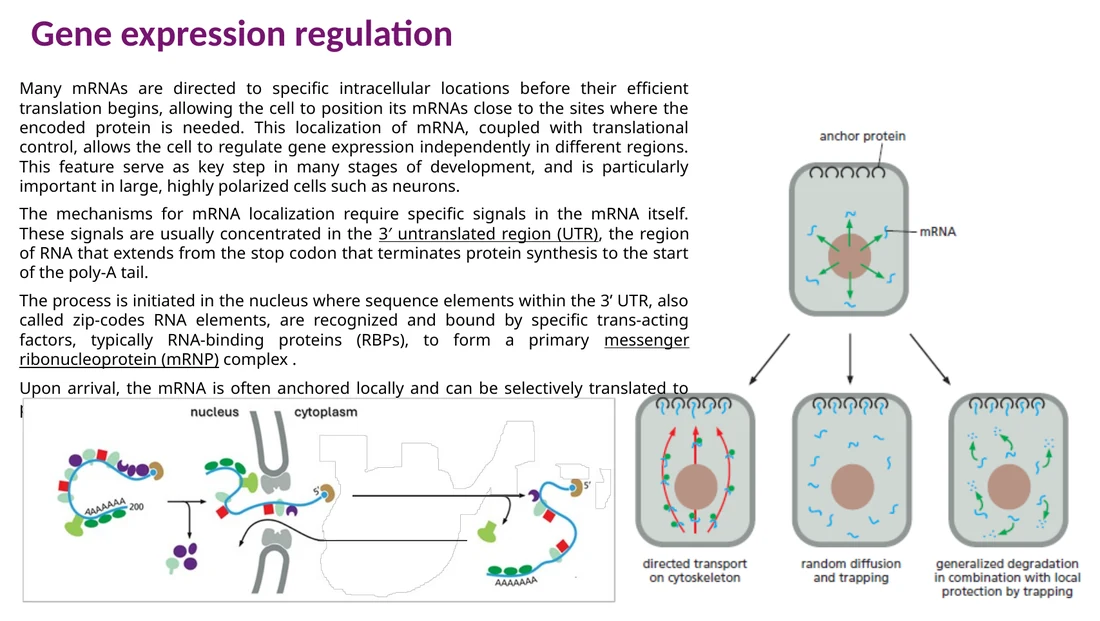
הבקרה הזאת חשובה במיוחד בתאים גדולים או מקוטבים, כמו נוירונים, שבהם ייצור חלבון מקומי בקצה התא מהיר ומדויק יותר מאשר ייצור בגרעין והובלת החלבון למרחק גדול.
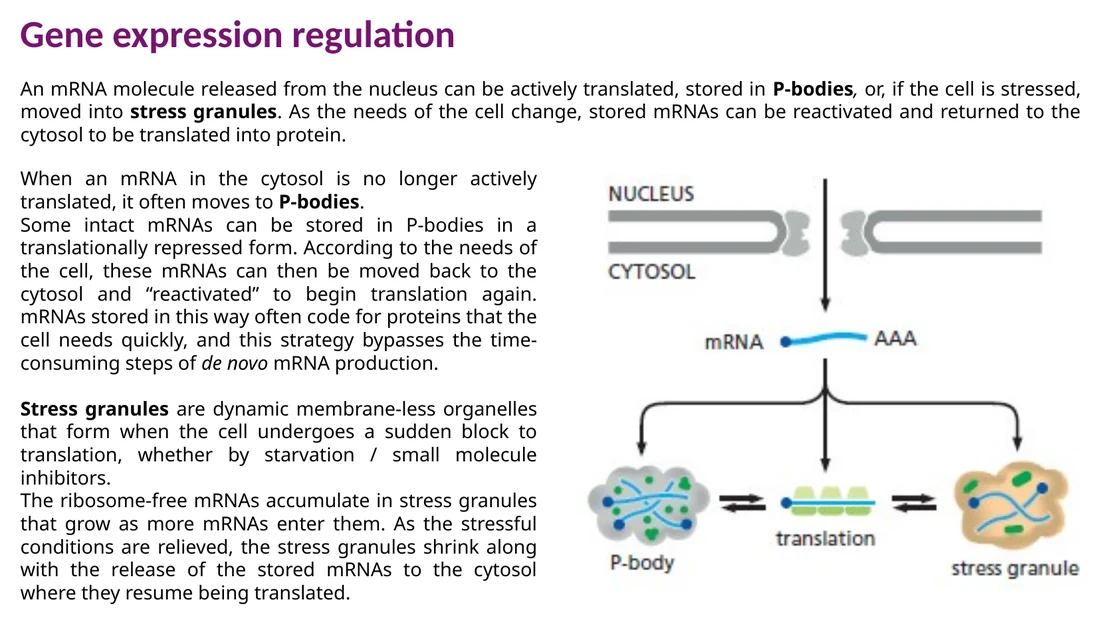
P-bodies ו־stress granules
mRNA שיצא לציטופלזמה יכול לעבור תרגום, להישמר זמנית או להתפרק. שני מבנים חסרי ממברנה חשובים כאן:
P-bodies עשירים באנזימי decapping, exonucleases וחלבונים שמעכבים תרגום. חלק מה־mRNAs נשמרים בהם וחלק עוברים פירוק.
Stress granules נוצרים בזמן עקה כאשר תרגום גלובלי נעצר. mRNAs שמשוחררים מריבוזומים נאספים בהם ויכולים לחזור לתרגום כאשר העקה חולפת.
בקרת תרגום
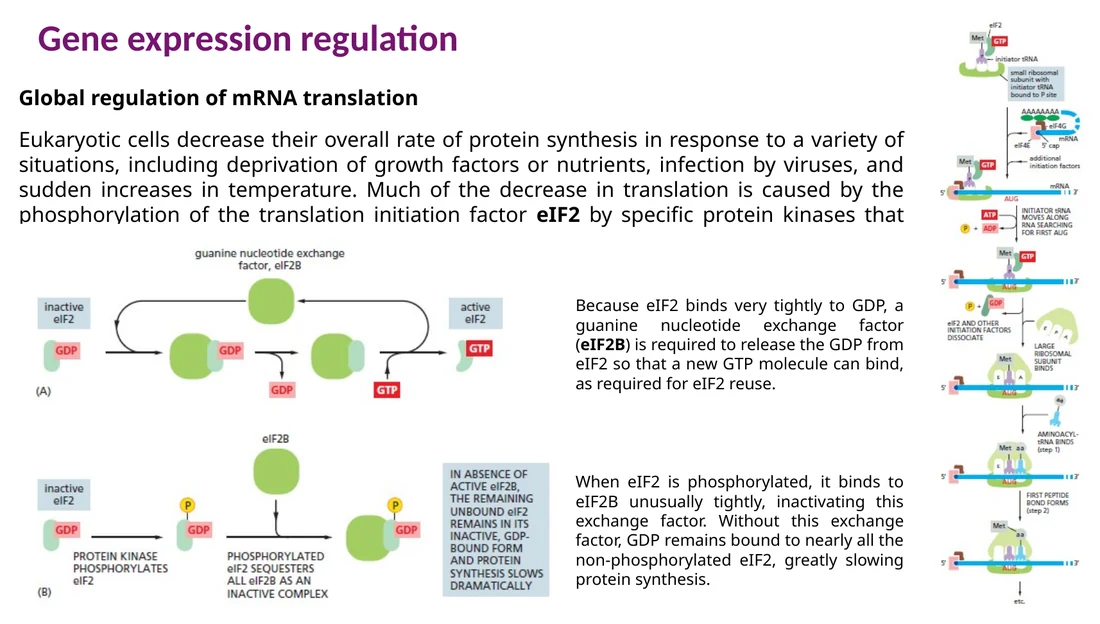
בקרה גלובלית דרך eIF2
במצבי עקה, חוסר נוטריינטים, חום או זיהום ויראלי, התא מוריד את קצב התרגום הכללי. אחד המנגנונים המרכזיים הוא פוספורילציה של eIF2.
במצב רגיל eIF2-GTP מביא את initiator Met-tRNA אל תת־היחידה הקטנה של הריבוזום. לאחר הידרוליזה ל־GDP, eIF2 צריך את eIF2B כדי להחליף GDP ב־GTP. פוספורילציה של eIF2 גורמת לו להיקשר חזק ל־eIF2B ולעכב אותו. התוצאה היא פחות eIF2-GTP ופחות initiation.
בקרה ספציפית דרך 5′ UTR
ה־5′ UTR קובע כמה טוב הריבוזום מגויס ל־mRNA וכמה טוב הוא מוצא את start codon. רצפים ומבנים באזור הזה יכולים להעלות או להוריד תרגום.
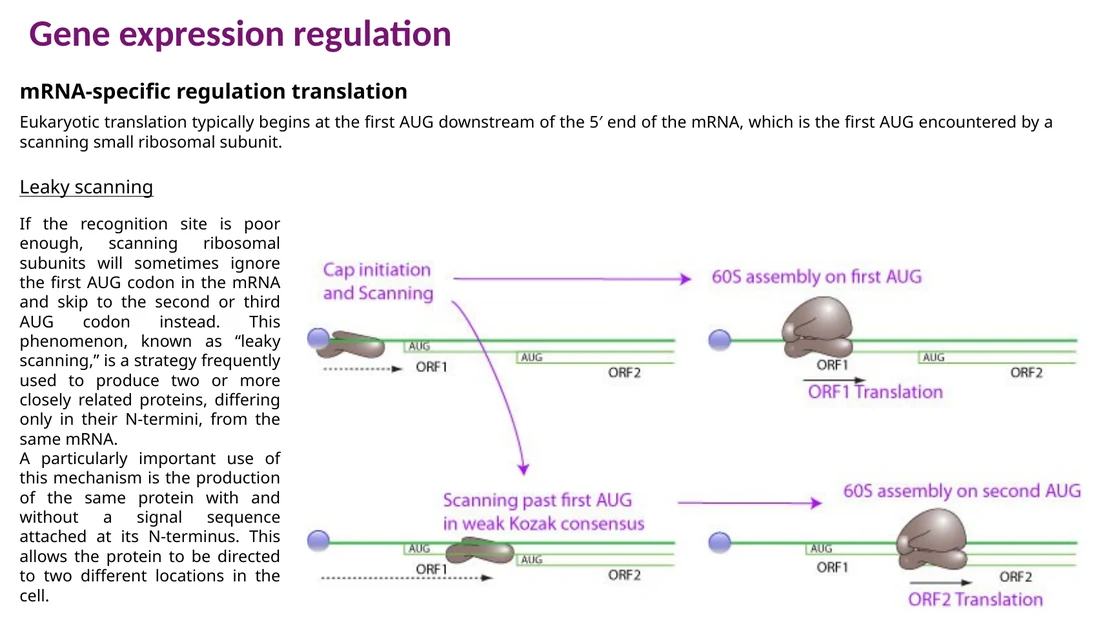
Leaky scanning
כאשר AUG ראשון נמצא בסביבת Kozak חלשה, תת־היחידה הקטנה יכולה לדלג עליו ולהתחיל ב־AUG downstream. כך מאותו mRNA יכולים להיווצר חלבונים עם N-terminus שונה.
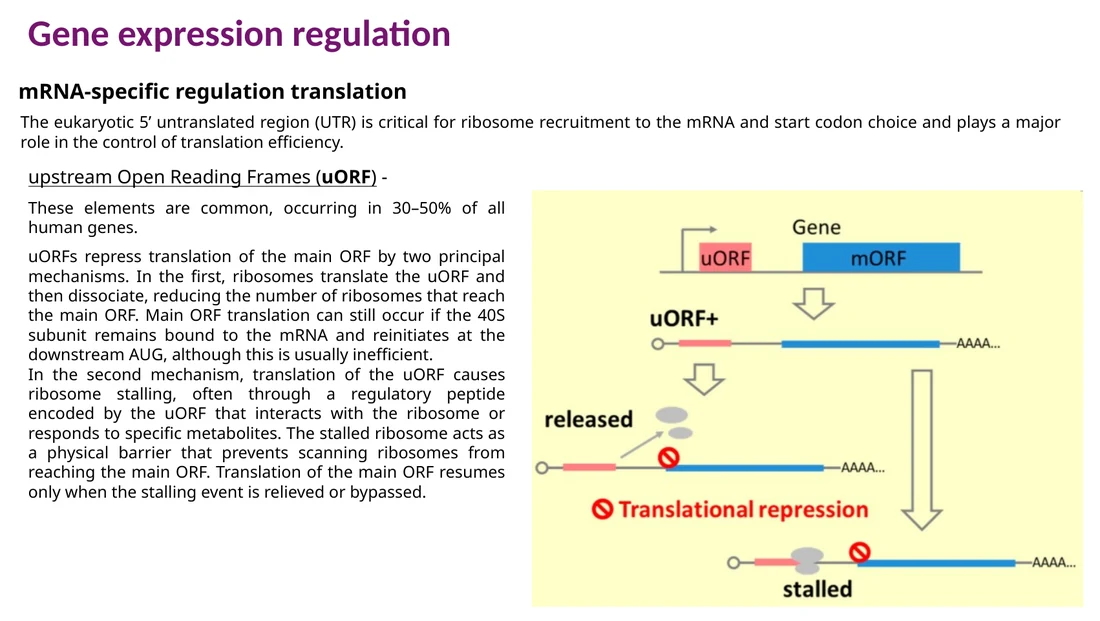
uORF
upstream ORF הוא ORF קצר שנמצא לפני ה־main ORF. תרגום של uORF יכול להוריד תרגום של החלבון העיקרי, כי הריבוזום מתנתק או נתקע לפני שהגיע ל־main AUG.
IRES
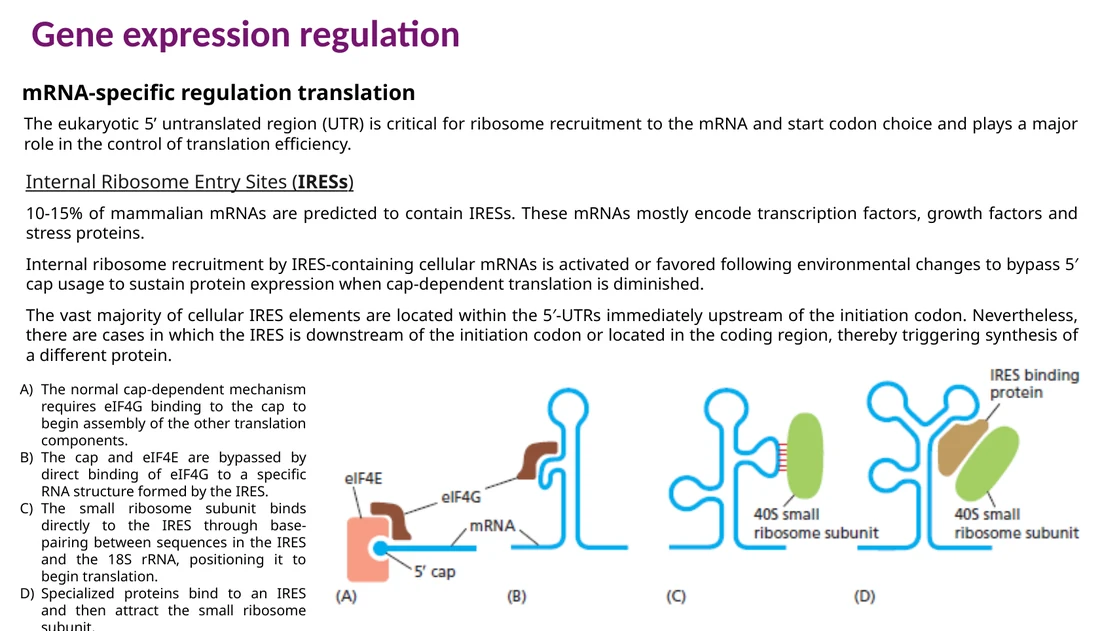
IRES מאפשר גיוס ריבוזום בצורה cap-independent. זה שימושי כאשר cap-dependent translation ירד, למשל בזמן עקה או זיהום ויראלי.
Stem-loop ו־IRE
מבנה stem-loop ליד ה־cap יכול לעכב scanning. דוגמה חשובה היא Iron Responsive Element ב־5′ UTR של ferritin mRNA. ברזל נמוך מאפשר ל־IRP להיקשר ל־IRE ולעכב תרגום של ferritin. ברזל גבוה מפחית את הקישור של IRP, ואז ferritin מתורגם ומסייע לאחסן עודפי ברזל.
RNA surveillance: בקרת איכות בזמן תרגום
הריבוזום משמש גם חיישן איכות. כאשר הוא נתקל בבעיה ב־mRNA, התא מפעיל מסלולי surveillance שמפרקים mRNA פגום, משחררים ריבוזומים ומצמצמים ייצור של חלבונים בעייתיים.
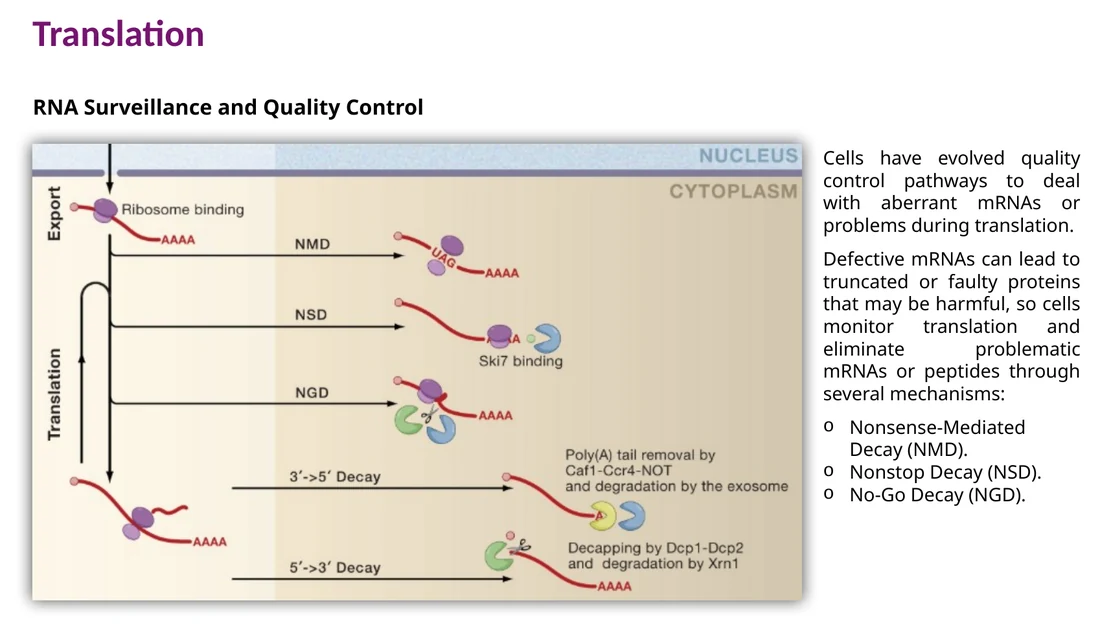
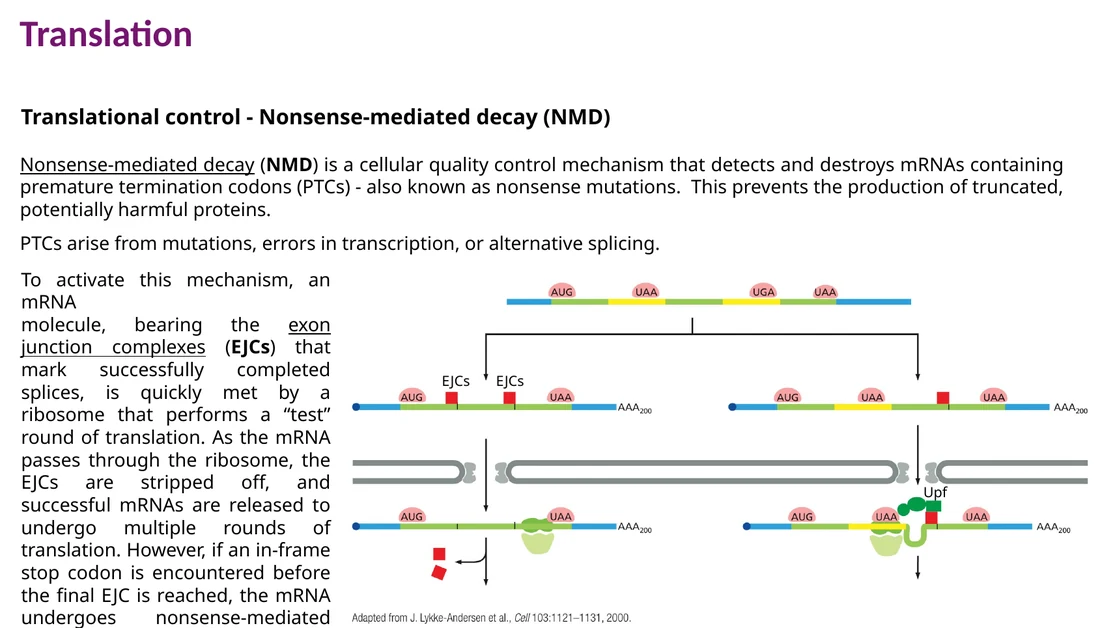
Nonsense-mediated decay - NMD
NMD מזהה mRNA עם stop codon מוקדם. לאחר splicing, אתרי exon-exon מסומנים על ידי EJC. בסיבוב התרגום הראשון, הריבוזום מסיר EJCs כאשר הוא עובר על ה־mRNA. אם הריבוזום מגיע ל־stop codon כאשר EJC עדיין נמצא downstream, המערכת מזהה premature termination codon ומגייסת חלבוני Upf שמובילים לפירוק ה־mRNA.
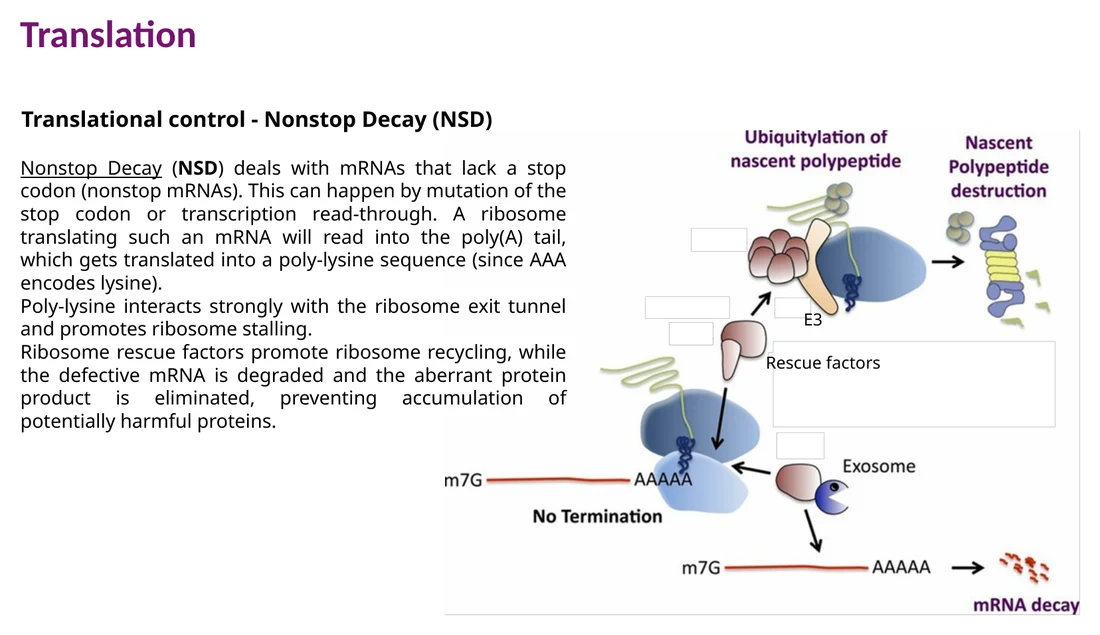
Nonstop decay - NSD
NSD מטפל ב־mRNA שאין בו stop codon תקין. הריבוזום ממשיך לתרגם אל תוך poly-A tail, נוצר רצף poly-lysine, והריבוזום נתקע. rescue factors משחררים את הריבוזום, וה־mRNA והתוצר החלבוני נשלחים לפירוק.
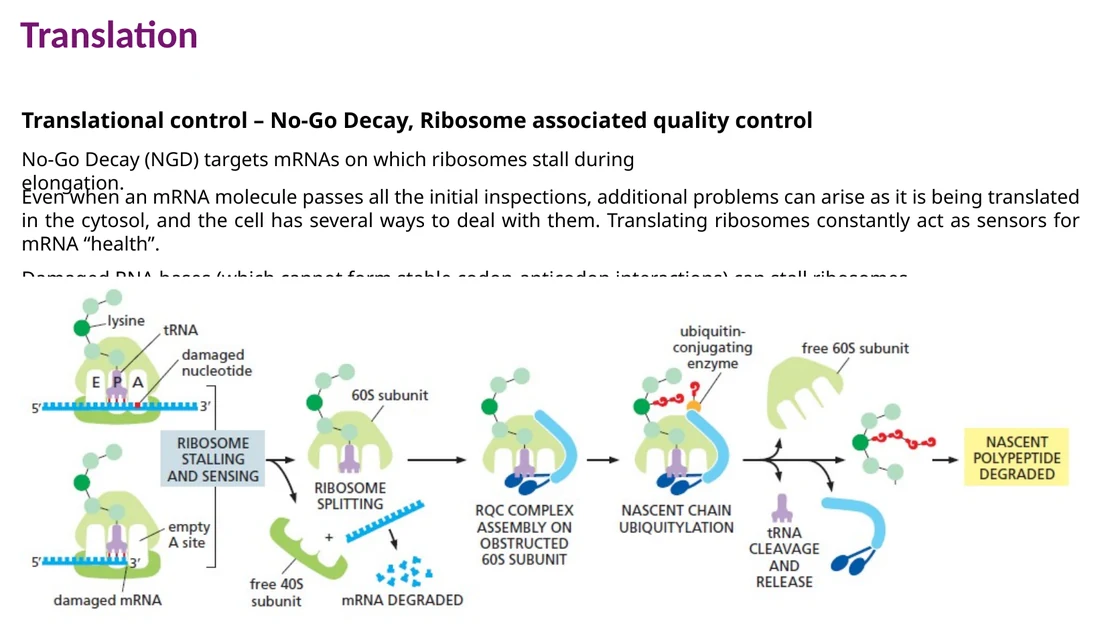
No-go decay - NGD
NGD מופעל כאשר ריבוזום נתקע בזמן elongation, למשל בגלל בסיס פגום ב־RNA, מבנה שניוני קשה או רצף שגורם לעצירה. ה־mRNA נחתך, הריבוזום ממוחזר והתוצר החלקי מטופל על ידי בקרת איכות חלבונית.
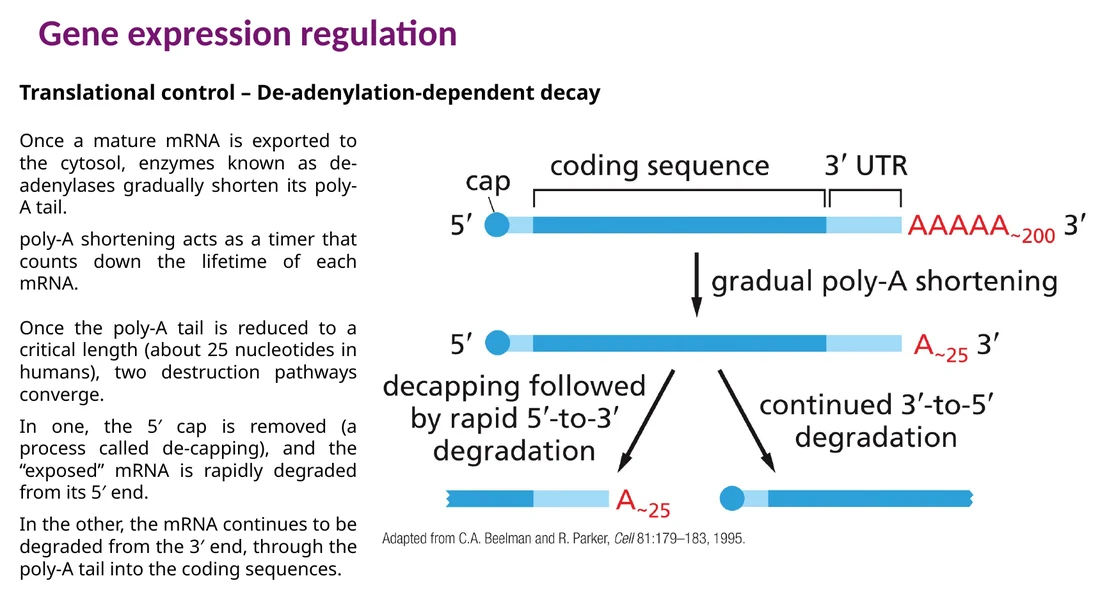
פירוק mRNA לפי אורך poly-A
ברוב mRNAs, poly-A tail מתקצר בהדרגה אחרי יציאה לציטופלזמה. האורך של הזנב מתפקד כמו שעון זמן חיים. כאשר הזנב מגיע לאורך קצר מאוד, בערך 25 נוקלאוטידים באדם, מתחילים מסלולי פירוק:
Decapping ואז פירוק $5’ \to 3’$ - ה־cap מוסר, וה־mRNA מתפרק מהקצה 5′.
פירוק $3’ \to 5’$ - exosome מתקדם דרך poly-A tail אל תוך הרצף המקודד.
רצפים ב־3′ UTR וחלבונים שנקשרים אליהם יכולים להאיץ או להאט deadenylation, decapping ופירוק. לכן 3′ UTR הוא אזור רגולטורי חשוב לזמן החיים של mRNA.
בקרה על חלבונים קיימים
פירוק מבוקר של חלבונים
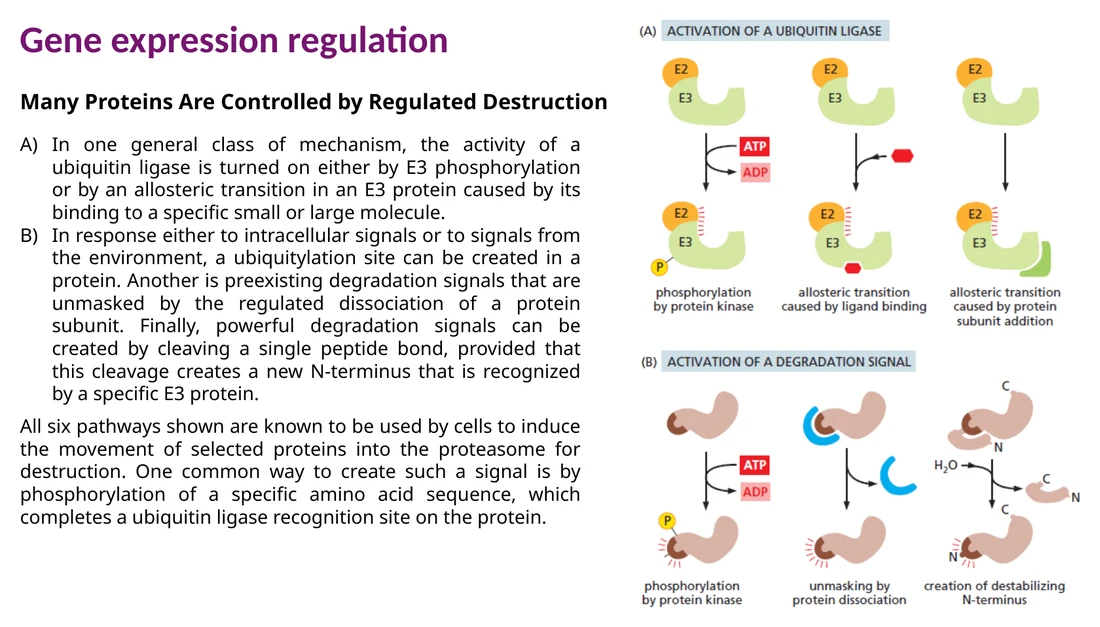
חלבון שכבר נוצר יכול להישלח לפירוק דרך ubiquitin-proteasome system. E1 מפעיל ubiquitin, E2 נושא אותו, ו־E3 ubiquitin ligase מזהה את החלבון הספציפי ומעביר אליו ubiquitin. שרשרת polyubiquitin, במיוחד דרך Lys48, מסמנת את החלבון ל־proteasome.
האות לפירוק יכול להיווצר בכמה דרכים: פוספורילציה שיוצרת אתר זיהוי ל־E3, חשיפת degron שהיה מוסתר, dissociation של תת־יחידה שמגנה על החלבון, או חיתוך פרוטאוליטי שיוצר N-terminus חדש שמזוהה על ידי E3.
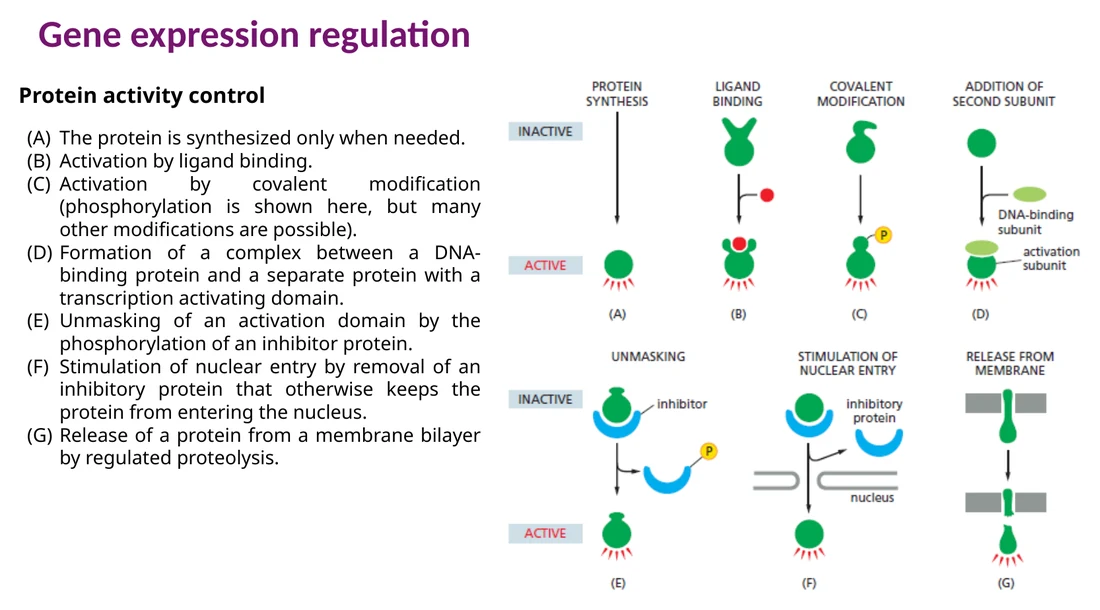
הפעלה של חלבונים
חלבון יכול להיווצר במצב פעיל או לעבור הפעלה לפי סיגנל. דרכי הפעלה נפוצות:
קישור ליגנד שמשנה קונפורמציה.
פוספורילציה או מודיפיקציה קוולנטית אחרת.
הרכבת קומפלקס עם תת־יחידה נוספת.
שחרור inhibitor שמסתיר active site, activation domain או NLS.
חיתוך פרוטאוליטי, למשל שחרור חלבון מהממברנה.
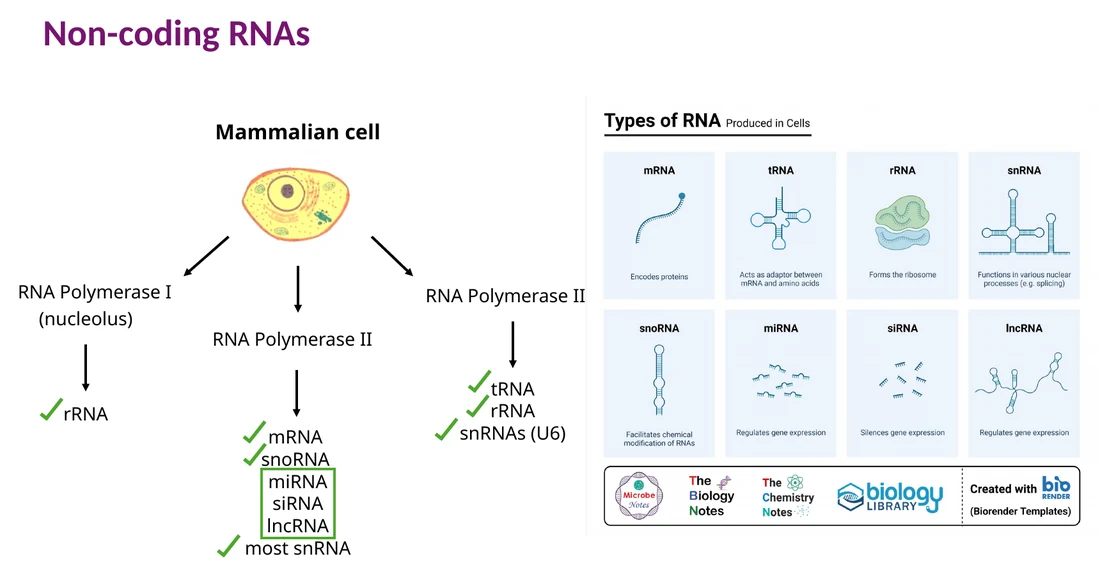
Non-coding RNAs
לא כל RNA מתורגם לחלבון. בקורס כבר הופיעו rRNA, tRNA, snRNA ו־snoRNA. כאן נוספות שלוש קבוצות רגולטוריות חשובות: lncRNA, miRNA ו־siRNA.
Long non-coding RNA - lncRNA
lncRNA מוגדר בדרך כלל כ־RNA באורך מעל 200 נוקלאוטידים ללא קידוד לחלבון. רבים מהם משועתקים על ידי RNA polymerase II, ולכן יכולים לקבל cap, poly-A ולעבור splicing. הפעולה שלהם נשארת ברמת RNA.
lncRNA יכול לפעול כ־scaffold שמקרב חלבונים שצריכים לעבוד יחד, כ־guide שמכוון קומפלקס חלבוני לרצף DNA או RNA מסוים, או כמרכיב של condensates כמו nucleolus, P-bodies ו־stress granules.
cis - ה־lncRNA פועל על אותו כרומוזום שממנו שועתק.
trans - ה־lncRNA עוזב את אתר השעתוק ופועל במקום אחר בגנום או בתא.
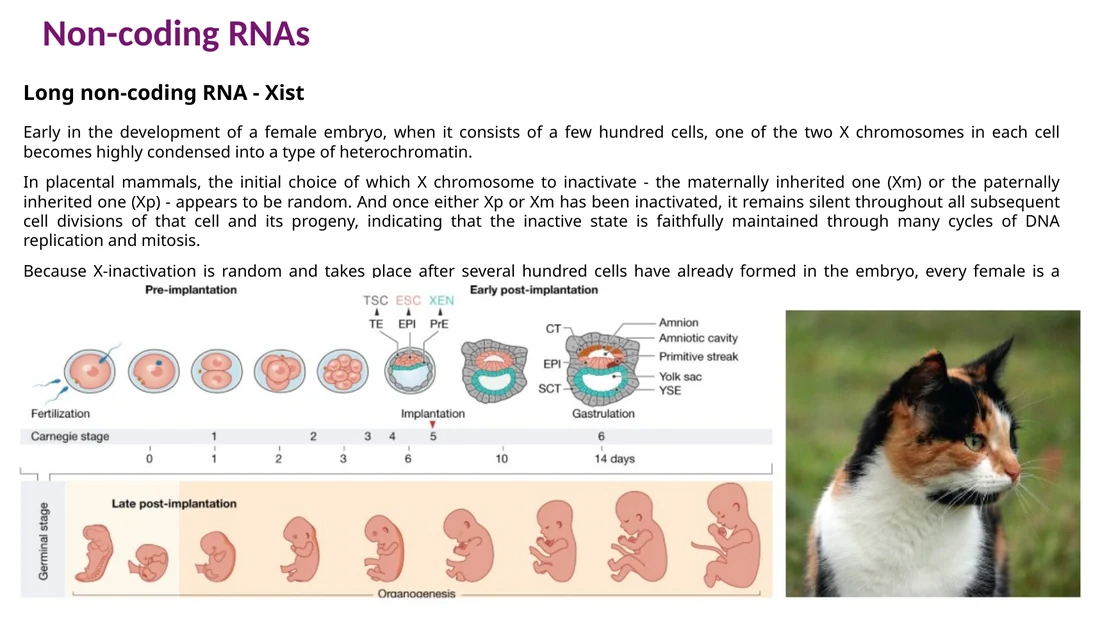
Xist ו־X chromosome inactivation
Xist הוא lncRNA חשוב ב־X chromosome inactivation. בתאי נקבה יש שני כרומוזומי X, ובזמן ההתפתחות אחד מהם מושתק כדי לאזן את מינון הגנים ביחס לתאים זכריים. Xist מתבטא מאחד מכרומוזומי X, מצפה אותו ומגייס מנגנוני השתקה כרומטינית.
הבחירה איזה X יושתק היא אקראית בשלבים מוקדמים של ההתפתחות, ולאחר מכן נשמרת בצאצאי התא. לכן נקבות הן mosaic: אזורים שונים בגוף יכולים לבטא X אימהי או X אבהי. בחתולות טריקולור רואים את זה היטב כי אחד הגנים לצבע פרווה נמצא על כרומוזום X.
RNA interference: miRNA ו־siRNA
RNA interference הוא מנגנון שבו RNA קצר מכוון קומפלקס חלבוני אל RNA מטרה. התוצאה יכולה להיות חיתוך ה־RNA, עיכוב תרגום או העברה ל־P-bodies.
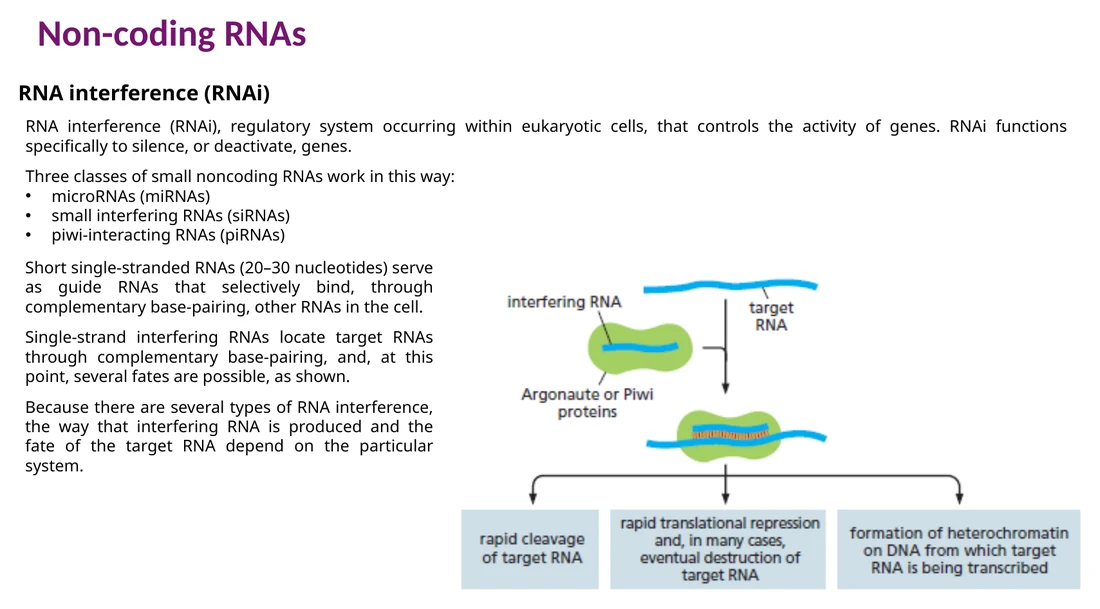
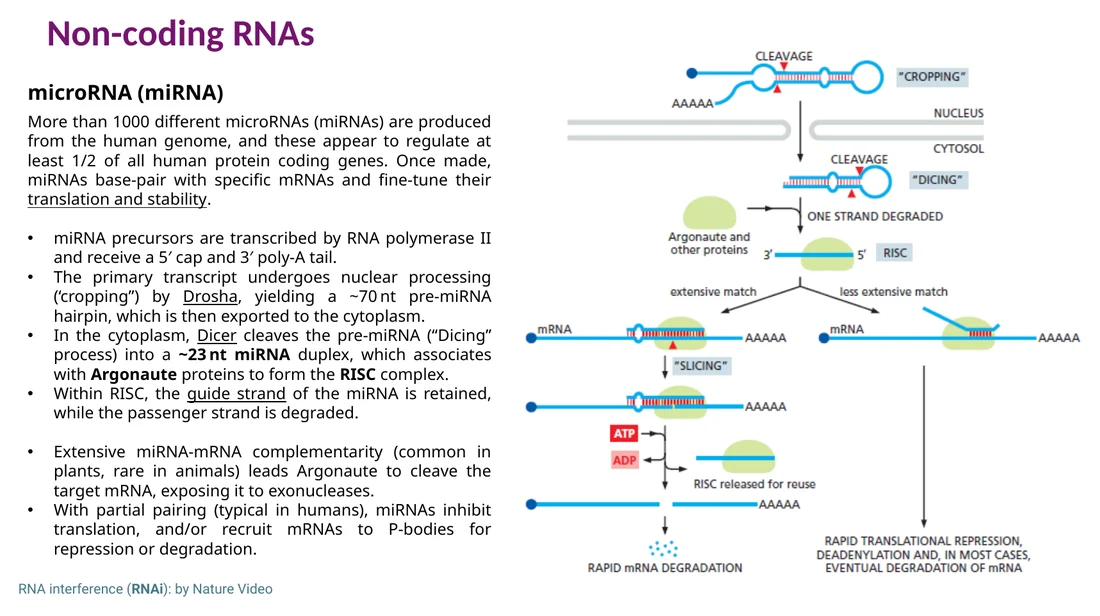
miRNA
miRNA הוא RNA קצר, בערך 23-20 נוקלאוטידים, שמקורו בגנום של התא. הוא משועתק על ידי RNA polymerase II כ־primary miRNA. בתוך הגרעין Drosha חותך אותו ל־pre-miRNA בצורת hairpin. ה־pre-miRNA יוצא לציטופלזמה, שם Dicer חותך אותו לדופלקס קצר. הדופלקס נטען על Argonaute ונוצר RISC. גדיל אחד נשאר כ־guide strand, והגדיל השני מתפרק.
בצמחים, התאמה כמעט מושלמת בין miRNA ל־mRNA מובילה לרוב לחיתוך ה־mRNA. בתאי בעלי חיים ההתאמה בדרך כלל חלקית, ולכן התוצאה הנפוצה היא עיכוב תרגום, deadenylation, מעבר ל־P-bodies ובהמשך פירוק.
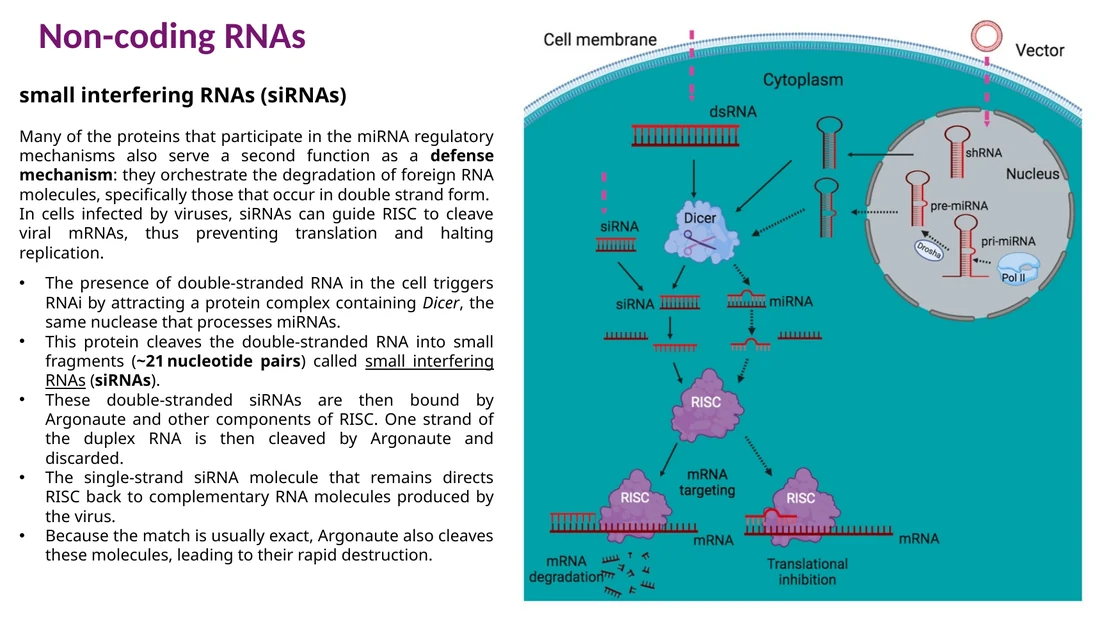
siRNA
siRNA נוצר לרוב מ־double-stranded RNA, למשל RNA ויראלי או RNA שהוכנס במעבדה. Dicer חותך את ה־dsRNA למקטעים קצרים. Argonaute/RISC שומר גדיל אחד, והגדיל הזה מזהה mRNA משלים. כאשר ההתאמה חזקה, Argonaute חותך את ה־mRNA והוא מתפרק.
במעבדה משתמשים במנגנון הזה ל־knockdown: מכניסים siRNA או shRNA שמתאימים ל־mRNA של גן מסוים, מורידים את רמת החלבון, ובודקים איך התא משתנה.
למבחן: miRNA מגיע לרוב מגנים של התא ומווסת mRNAs של התא. siRNA מגיע לרוב מ־dsRNA חיצוני או מניסוי, ובדרך כלל מכוון לחיתוך מדויק של RNA משלים.