חזקים בחלבונים?
נסו 10 שאלות בנושא, עם עדיפות לשאלות שלא הצלחתם. בסיום תקבלו ציון וסקירה על הטעויות שלכם. הצלחתם? גלו בחנים בדפים נוספים
תוצאות המבחן
היסטוריית מבחנים
טוען את השאלות...
אנא המתן...
מבוא לחלבונים וגנטיקה
החומר על חלבונים הוא קריטי להמשך הקורס. חלק זה יתמקד במבנה ובתפקוד חלבונים, ולאחר מכן יעבור לעקרונות יסוד בגנטיקה.
קשרים דיסולפידיים בחלבונים
קשרים דיסולפידיים (Disulfide) הם קשרים קוולנטיים הנוצרים בין שתי שרשראות צדדיות של חומצת האמינו ציסטאין. בניגוד לרוב הקשרים המייצבים את המבנה התלת־ממדי של חלבונים, שאינם קוולנטיים, קשרים דיסולפידיים הם יוצא דופן.
הקשר הדיסולפידי נוצר כאשר קבוצת SH בקצה השרשרת הצדדית של ציסטאין מגיבה עם קבוצת SH נוספת.
\[\text{R-SH + HS-R} \rightleftharpoons \text{R-S-S-R + 2H}^+ \text{ + 2e}^-\]יצירת קשר דיסולפידי תלוי במצב החמצון-חיזור של הסביבה. קשר כזה יכול להיווצר בין שתי שרשראות פוליפפטידיות שונות או בתוך אותה שרשרת, ובכך לייצב את המבנה השלישוני (הקיפול המרחבי) או הרביעוני (קומפלקס של מספר שרשראות) של החלבון.
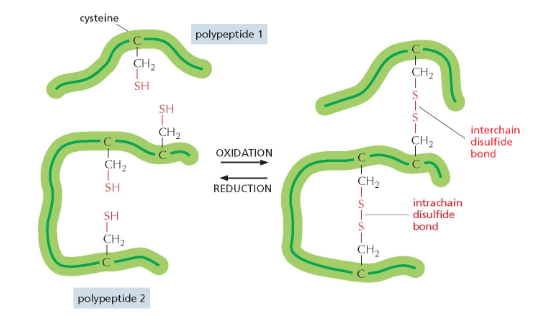
סביבת חמצון-חיזור והשפעתה על קשרים דיסולפידיים
המעבר בין שני מצבי הקשר הדיסולפידי הוא תהליך חמצון-חיזור. בסביבה מחזרת, כמו הציטופלזמה של תאים, לא ייווצרו קשרים דיסולפידיים.
לעומת זאת, חלבונים המופרשים החוצה מהתא, כמו הורמון האינסולין, או חלבונים הממוקמים בממברנה החיצונית ופונים החוצה, נמצאים בסביבה מחמצנת. בסביבה מחמצנת, הקשרים הדיסולפידיים תורמים רבות ליציבות המבנה. דוגמה נוספת הם נוגדנים, שהמבנה שלהם מיוצב על ידי קשרים דיסולפידיים.
דוגמת האינסולין
אינסולין הוא דוגמה מצוינת לחלבון קטן (51 חומצות אמינו באדם) המיוצב על ידי קשרים דיסולפידיים. הוא מיוצר תחילה כפוליפפטיד ארוך, וחלק ממנו מסולק בתהליך ההבשלה לצורה הפעילה של ההורמון.
במבנה הסופי של אינסולין יש שלושה קשרים דיסולפידיים: אחד בתוך שרשרת ושניים בין השרשראות. כדי להגיע למבנה הסופי נדרשת עזרה של גורמי הרכבה (assembly factors) – פרוטאזות, חלבונים החותכים ברצף ספציפי של חומצות אמינו. זהו חלק מתהליך רב־שלבי הדרוש להבשלת אינסולין לצורתו הפעילה.
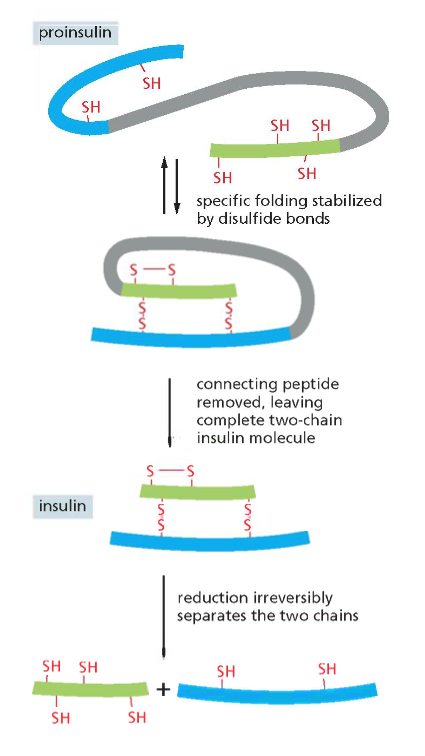
היכן נוצרים הקשרים הדיסולפידיים?
כפי שצוין, בציטופלזמה הסביבה היא מחזרת, ולכן קשרים דיסולפידיים אינם יכולים להיווצר שם. קשרים אלה נוצרים בתוך הרשתית האנדופלזמטית (ER), בחלל המימי (הלומן) הכלוא בממברנות ה־ER ומופרד מהציטופלזמה.
זוהי דוגמה ראשונה לעיקרון חשוב: חלבונים מגיעים לצורתם הבשלה והפעילה בתאים איקריוטים דרך מדורים שונים. אם נחשוף אינסולין בשל לסביבה מחזרת, הקשרים הדיסולפידיים יתפרקו, והחלבון לא ידע ליצור מחדש את הקשרים בצורה הנכונה. כלומר, המבנה התלת־ממדי הסופי שלו תלוי ביצירה נכונה של הקשרים הדיסולפידיים ובפעילות חיצונית של גורמי ההרכבה.
אתרי קישור ומבנה תלת־ממדי
הפעילות הביולוגית של חלבונים תלויה בהיכרות מולקולרית ספציפית, והיכרות זו היא תלת־ממדית. עיקרון מרכזי בביולוגיה הוא שהמבנה התלת־ממדי של חלבונים קובע את פעולתם.
התאמה מדויקת במרחב
העיקרון המרכזי הוא של “Precise Fit” – התאמה תלת־ממדית מושלמת, כמו יד לכפפה או מפתח למנעול. אתרי קישור הם מקומות ספציפיים בחלבון המאפשרים אינטראקציה עם מולקולות אחרות. לעיתים, כל המידע לבניית המבנה הנכון כבר קיים ברצף החלבון, והקיפול נוצר בזכות התאמה תלת־ממדית.
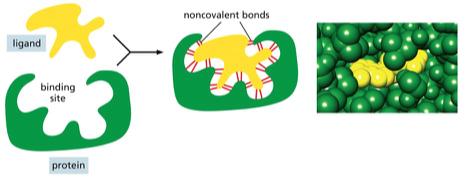
ליגנדים ורצפטורים
באינטראקציות בין חלבונים למולקולות אחרות, משתמשים במונחים “ליגנד” (Ligand וגם ״קשיר״) ו”רצפטור”. ליגנד הוא מולקולה קטנה יחסית הנקשרת לחלבון. לדוגמה, אינסולין יכול להיחשב כליגנד כאשר הוא נקשר לרצפטור שלו. אתר הקישור בחלבון בנוי תלת־ממדית על ידי שיירים של חומצות אמינו הנמצאים במקומות שונים במבנה.
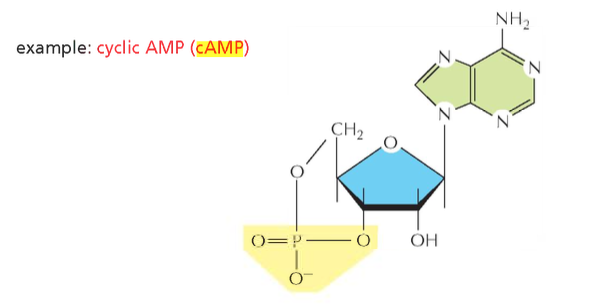
דוגמה: Cyclic AMP והחלבון הקושר אותו
הדוגמה במצגת מראה חלבון המכיר את המולקולה Cyclic AMP (cAMP), שליח משני חשוב בתא. החלבון יוצר כיס תלת־ממדי שבו הליגנד נקשר. חומצות האמינו היוצרות את הקשר יכולות להיות במקומות שונים לחלוטין ברצף הראשוני, אך הקיפול התלת־ממדי מקרב אותן ומאפשר את הקישור.
מבנה מולקולת ה־cAMP כולל בסיס חנקני (אדנין), סוכר ריבוז, וקבוצת פוספט הקשורה לפחמנים 5’ ו־3’ (יוצרת מבנה מעגלי).
השפעת מוטציות על פעילות החלבון
מוטציה באחת מחומצות האמינו באתר הקישור עלולה לפגוע משמעותית בפעילות החלבון. לדוגמה, החלפת חומצת אמינו חיובית בשלילית עלולה ליצור דחייה אלקטרוסטטית שתפריע לאינטראקציה. לא תמיד ניתן לחזות במדויק את השפעת השינוי, אך השלב הראשון בניתוח הוא סימולציה תלת־ממדית, ולאחר מכן ניסויים לאימות התחזית.
אנזימים ופעילותם
ליזוזים - דוגמה לאנזים
ליזוזים הוא האנזים הראשון שהמבנה התלת־ממדי שלו פוענח (בשנות ה־60). זהו אנזים קטן, המהווה חלק ממערכת ההגנה מפני חיידקים. הוא חותך שרשראות של פפטידוגליקן, מרכיב עיקרי בדופן של חיידקים רבים. הליזוזים נמצא בחלבון ביצה, ברוק, בדמעות ובמוקוזה, שם הוא מופרש על ידי תאי אפיתל וגורם לליזיס (התפוצצות) של חיידקים.
מנגנון פעולה אנזימטי
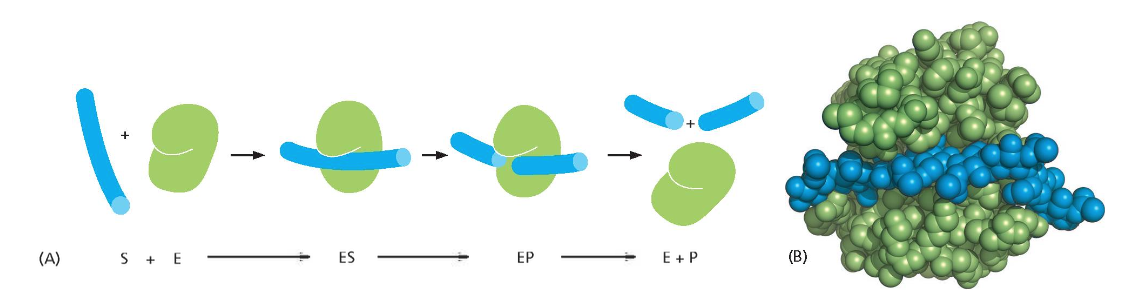
האנזים ליזוזים פועל באופן הבא:
- הסובסטרט (שרשרת הרב־סוכר) נכנס לשקע (cleft) בחלבון.
- האנזים יוצר מתח (strain) על הסוכר במקום החיתוך, ובכך מתגבר על המחסום האנרגטי של הריאקציה.
- במהלך הריאקציה נוצר קשר קוולנטי זמני (transient) בין האנזים לסובסטרט.
- חומצת אמינו ספציפית באנזים קריטית לפעילות זו.
הריאקציה היא הידרוליזה (פירוק כימי בעזרת מים), המועדפת אנרגטית אך אינה מתרחשת ספונטנית עקב המחסום האנרגטי. האנזים מזרז את הריאקציה בכ־6 סדרי גודל (פי מיליון).
מנגנון פעולה אנזימטי - המשך
חומצות אמינו מסוימות בליזוזים הן קריטיות לא רק לקישור הסובסטרט אלא גם לריאקציה עצמה. במהלך הריאקציה נוצר מצב ביניים זמני וקצר-חיים. בשלב הבא, חומצת אמינו נוספת נכנסת לפעולה ומארגנת מולקולת מים בגיאומטריה מדויקת המאפשרת התקפה נוקלאופילית.
מצב הביניים נשבר, והאנזים חוזר למצבו המקורי. מולקולת המים מתחלקת בין שני צדי הקשר שנחתך, ונוצרים שני תוצרים המשתחררים מהאנזים. האנזים יכול לחזור על התהליך פעמים רבות, לבצע חיתוכים רבים במולקולות הפפטידוגליקן, עד שהחיידק אינו יכול לעמוד בנזק ועובר ליזיס. מכאן שמו של האנזים – ליזוזים.
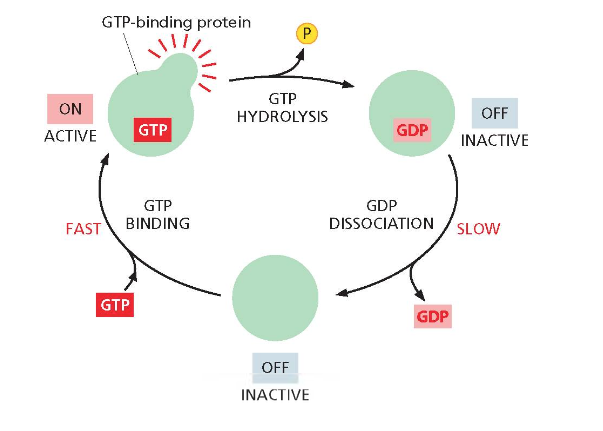
חלבוני G - מתגים מולקולריים
חלבוני G הם משפחה של חלבונים הקושרים GTP (גואנוזין תלת־פוספט) ופועלים כאנזימים המפרקים אותו ל־GDP ופוספט. החלבון עובר בין שני מצבים: מצב “פעיל” (on) כאשר הוא קושר GTP, ומצב “כבוי” (off) כאשר הוא קושר GDP.
חלבונים אלו משמשים כמתגים מולקולריים (molecular switches). במצב ״פעיל״, החלבון מפעיל שרשרת של חלבונים אחרים. המעבר בין המצבים אינו ספונטני ודורש חלבוני עזר.
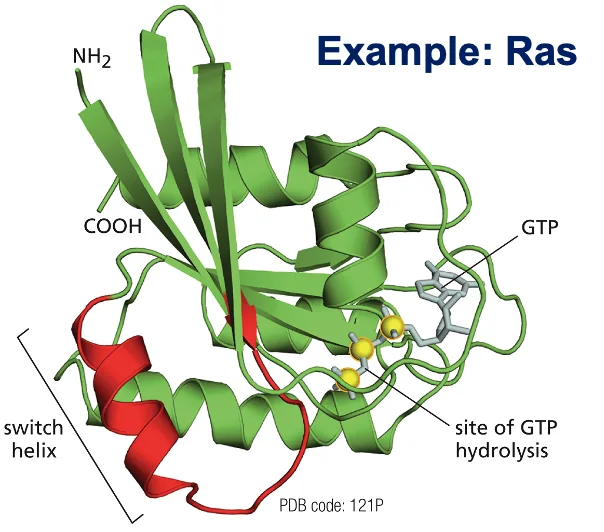
דוגמה: חלבון RAS
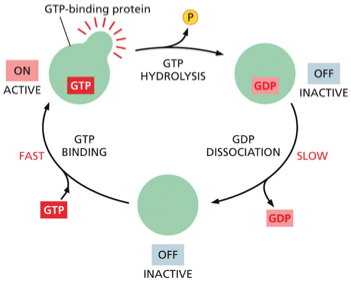
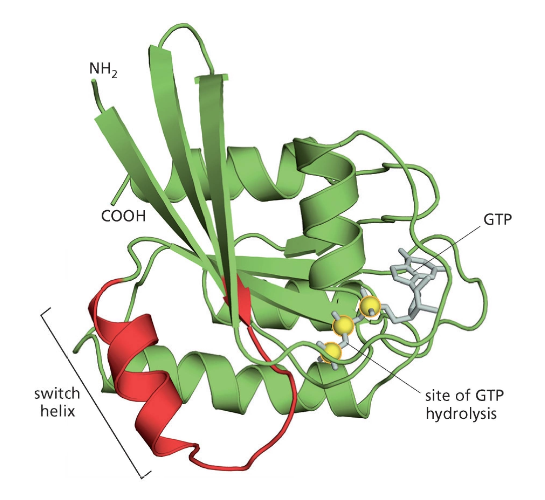
RAS הוא חלבון מפורסם ממשפחה זו. מוטציות בגן המקודד ל־RAS נמצאות בכ־50% מהגידולים הסרטניים באדם. במבנה של RAS יש אתר קישור ל־GTP. ההידרוליזה של GTP ל־GDP גורמת לשינוי במבנה התלת־ממדי של החלבון, בעיקר באזורים הנקראים “switch helix”.
כאשר RAS במצב “פעיל”, הוא מעודד חלוקות תא. מוטציה הגורמת לו להישאר “תקוע” במצב הפעיל מובילה לחלוקות תאים בלתי מבוקרות – מאפיין מרכזי של סרטן. טעינה מחדש של החלבון ב־GTP דורשת אנרגיה, מה שמדגים את העיקרון שתאים משקיעים אנרגיה רבה בשמירה על סדר.
מערכת האוביקוויטין ופירוק חלבונים
בסיום פרק החלבונים, יש להזכיר את אברהם הרשקו ואהרן צ’חנובר, שקיבלו פרס נובל בשנת 2004 על גילוי מערכת האוביקוויטין.
אוביקוויטין הוא חלבון קטן (76 חומצות אמינו). מערכת שלמה בתא מחברת אוביקוויטין לחלבונים אחרים. האוביקוויטין נקשר לחלבונים דרך קבוצת צד של חומצת האמינו ליזין, בקשר הנקרא “קשר איזופפטידי” (isopeptide bond).
חיבור של מולקולות אוביקוויטין לחלבון מהווה מעין “קוד” נוסף, המשפיע על פעילותו. אחת הפעילויות החשובות של קוד זה היא סימון חלבונים לפירוק (degradation). לכל חלבון יש אורך חיים אופייני, והמערכת מסמנת חלבונים פגומים או כאלה שהגיע זמנם להתפרק. תגלית זו פתחה תחום שלם בביולוגיה.
מבוא לעקרונות בסיסיים בגנטיקה
חלק זה של הקורס יעסוק בעקרונות של תורשה, גנטיקה וחלוקת תא. זהו מבוא למושגי יסוד שיידרשו בהמשך.
הנזיר מנדל והתחלת הגנטיקה המודרנית
גרגור מנדל, נזיר שעסק בגנטיקה של צמחי אפון, הניח את היסודות לגנטיקה המנדלית הבסיסית. במשך מאות שנים, בני אדם ניסו להשביח זנים של בעלי חיים וצמחים מבלי להבין את עקרונות התורשה לעומק.
שלושה ענקי מדע שהניחו את היסודות לגנטיקה המודרנית
במחצית השנייה של המאה ה־19, שלושה מדענים הניחו את היסודות לגנטיקה ולביולוגיה מודרנית:
צ’רלס דרווין
דרווין פיתח את תורת האבולוציה, המהווה רקע לכל הקורסים הביולוגיים. תובנותיו התבססו על תצפיות, כמו אלו שערך באיי גלאפגוס על מיני הפרושים. עקרון הברירה הטבעית שהציע מסביר כיצד תכונות יעילות נוטות להישמר באוכלוסייה. הבעיה של דרווין הייתה חוסר ידע בגנטיקה, והוא לא הכיר את עבודתו של מנדל.
גרגור מנדל
מנדל עקב אחר שבע תכונות ברורות של צמחי אפון, תוך שימוש בזנים טהורים וגישה כמותית. הוא גילה שני חוקים חשובים:
- החוק הראשון של מנדל: שני האללים של כל גן נפרדים זה מזה בעת יצירת הגמטות.
- החוק השני של מנדל: גנים שונים מתנהגים באופן בלתי תלוי זה בזה.
תגליותיו נותרו לא ידועות במשך כ־30 שנה.
וולטר פלמינג
פלמינג ערך תצפיות מיקרוסקופיות וצייר תאים מתחלקים. הוא טבע את המונח “כל תא מקורו בתא קודם” (Omnis cellula e cellula) וזיהה את הכרומטין, הכרומוזומים והמיטוזה.
ספרות מומלצת לגנטיקה
לנושא הגנטיקה, ספר מומלץ הוא “Genetics” של Brooker.
מושגי יסוד בגנטיקה
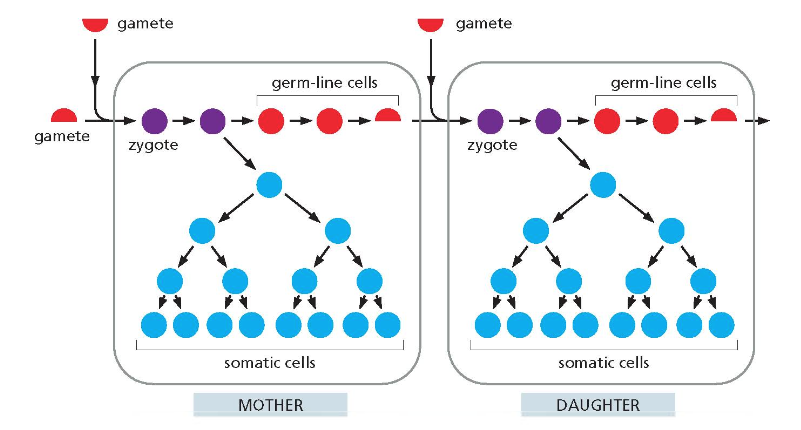
- גונדות: (Gonads) בלוטות המין (שחלות, אשכים).
- גמטות: (Gametes) תאי המין (ביציות, זרעונים).
- זיגוטה: (Zygote) הביצית המופרית, התא הראשון של אורגניזם חדש.
- תאים סומטיים: (Somatic cells) כל תאי הגוף שאינם תאי מין.
הגונדות מייצרות את הגמטות. איחוי הגמטות יוצר את הזיגוטה. לפני איחוי הגמטות מתרחש שכפול DNA, עיקרון המבוסס על מבנה הסליל הכפול. המודל המתואר הוא של רבייה מינית.
גמטות והחוקים של מנדל
יצירת הגמטות והפרדת זוגות הגנים הם תהליכים שמנדל הבין, והקישור ביניהם הוא סיפורה של הביולוגיה המודרנית. החלוקה היוצרת את הגמטות מתרחשת בגונדות והיא חלוקה מיוחדת המפחיתה את המטען הגנטי.
מושגי יסוד נוספים
תאי גוף הם תאים סומטיים. מוטציות סומטיות (כמו ב־RAS) מתרחשות בתאי גוף ואינן עוברות בתורשה.
- רבייה לא מינית: יוצרת את התאים הסומטיים.
- רבייה מינית: מתרחשת בגונדות ויוצרת את הגמטות.
המושג “סומא” מתייחס לכל מה שאינו שייך לקו הנבט (germ-line cells). בקו הנבט מתרחשת חלוקה מיוחדת לפי חוקי מנדל. מושג חשוב נוסף הוא “אלל” – אחד משני העותקים של גן.
דוגמאות לתורשה מנדלית
מנדל עבד עם צמחי אפון, שבהם איברי מין זכריים ונקביים נמצאים באותו פרח, מה שמאפשר הפריה עצמית. הוא ביצע הכלאות מבוקרות בין זן סגול לזן לבן. לאחר שזרע את האפונים שנוצרו, הוא עקב אחר שלושה דורות.
דומיננטיות ורצסיביות
התכונה של צבע סגול נמצאה כדומיננטית על פני צבע לבן, מכיוון שבדור הראשון כל הפרחים היו סגולים. האלל הלבן לא נעלם, אלא הופיע מחדש בדור השני. זו הייתה פריצת הדרך של מנדל: קיימים “פקטורים” (גנים) שאינם מתערבבים. היחס בדור השני היה 3 סגולים לכל 1 לבן.
האם דומיננטי = חזק/טבעי ורצסיבי = חלש/פגום?
התכונה הדומיננטית היא זו הנראית חיצונית (פנוטיפ). הגנוטיפ כולל את שני האללים. אין להניח שתכונה דומיננטית היא “חזקה” או “טבעית” ושתכונה רצסיבית היא “חלשה” או “פגומה”. לדוגמה, עיניים כחולות הן תכונה רצסיבית.
במקרה של צבע הפרח, ייתכן שהגן מקודד לאנזים המייצר פיגמנט, ובפרח הלבן האנזים אינו פעיל. אך זה לא תמיד המצב. למעשה, ארבע מהתכונות שמנדל בדק הן רצסיביות והן הנפוצות ביותר בטבע בצמחי אפון.
דוגמה למלאנין והקשר בין גנטיקה וביולוגיה מולקולרית
דוגמה זו מחברת בין הדוגמה המרכזית של הביולוגיה ($\text{DNA} \rightarrow \text{RNA} \rightarrow \text{protein}$) לגנטיקה קלאסית. היא עוסקת ביצירת מלאנין, פיגמנט בעור, בעיניים ובשיער. בתא עור (מלנוציט) עם שני עותקים של האלל הדומיננטי (A):
- שני עותקי ה־DNA מתועתקים ל־RNA.
- נוצר פוליפפטיד המתקפל לאנזים פעיל (טירוזינאז).
- האנזים מייצר מלאנין מהסובסטרט טירוזין.
אם יש פגם באלל אחד בלבד (מצב הטרוזיגוטי), לרוב עדיין ייווצר מלאנין, מכיוון שהעותק השני של הגן תקין. רק כאשר שני העותקים של הגן פגומים (למשל, עקב מוטציה המשנה חומצה אמינית באתר הפעיל), לא נוצר אנזים פעיל כלל, ומלאנין לא נוצר.
זוהי הדוגמה של אלביניזם. במקרה זה, האלל המוטנטי הוא רצסיבי וגורם לחוסר פעילות של האנזים (loss of function).
חלוקת תא - מיטוזה ומיוזה
קיימת חלוקה לא מינית (סומטית) וחלוקה מינית, שהיא הבסיס לחוקי מנדל.
- הפלואידי (n): עותק אחד מכל גן בגנום.
- דיפלואידי (2n): שני עותקים מכל גן.
החלוקה המינית, מיוזה, מתרחשת בגונדות ויוצרת תאים הפלואידים (גמטות). לאחר הפריה, הזיגוטה חוזרת למצב דיפלואידי.
מיטוזה - חלוקה לא מינית
מיטוזה היא חלוקה לא מינית שבה תא אחד מתחלק לשני תאי בת זהים גנטית. לפני החלוקה, כל כרומוזום מוכפל בתהליך שכפול DNA. כל כרומוזום נכנס לחלוקה עם שתי זרועות זהות הנקראות “כרומטידות אחיות”. כל כרומטידה היא מולקולת DNA אחת. במהלך המיטוזה, הכרומטידות האחיות נפרדות. מטרת המיטוזה היא לשמר את המידע הגנטי הקיים.
מיוזה - חלוקה מינית
מיוזה כוללת שתי חלוקות עוקבות ללא שכפול DNA נוסף ביניהן. במיוזה מתרחשים שני תהליכים חשובים:
- הפחתת המטען הגנטי: מדיפלואידי להפלואידי.
- יצירת שונות גנטית.
במהלך המיוזה, כרומוזומים הומולוגיים (אחד מהאם ואחד מהאב) נצמדים זה לזה. בשלב זה יכול להתרחש תהליך של “שחלוף” (crossing over), שבו קטעים של כרומוזומים מוחלפים ביניהם, מה שיוצר ערבוב גנטי.
כך, אללים שונים של אותו גן (למשל, A ו־a) נפרדים זה מזה (החוק הראשון של מנדל). בנוסף, גנים שונים (למשל, A/a ו־B/b) יכולים להתנהג באופן בלתי תלוי (החוק השני של מנדל). השחלוף הוא מנגנון מורכב העומד בבסיס השונות הגנטית.
סיכום
לחלוקה המינית (מיוזה) שתי מטרות עיקריות: הפחתת המטען הגנטי ויצירת שונות גנטית רבה. זו הסיבה ששילובים של תכונות בצאצאים יכולים להיות שונים מהצפוי, במיוחד כאשר עוקבים אחר גנים רבים בו־זמנית.
| נושא | מונחים חשובים | תהליך עיקרי | דוגמה |
|---|---|---|---|
| קשר דיסולפידי | Cys, SH, S-S | חיזור/חמצון | אינסולין |
| חוקי מנדל | אלל, גנוטיפ | הפרדת אללים | אפונים: סגול/לבן |
| חלוקת תא | מיטוזה/מיוזה | שכפול כרומוזומים | זיגוטה ← הפלואידי |
| חלבון-ליגנד | אתר קישור, התאמה תלת ממדית | אינטראקציה ספציפית | cAMP |
| אנזימים | סובסטרט, אתר פעיל | הידרוליזה | ליזוזים |
| RAS | GTP ↔ GDP | מתג מולקולרי | סרטן |
| אוביקוויטין | ליזין, פירוק | isopeptide | הרשקו & צ’חנובר |