פתיחה: מה מחפשים בשיעור הזה?
בשיעור הקודם עקבנו אחרי שבירת סימטריה בשלבים המוקדמים: זיגוטה, שני תאים, ארבעה תאים, שמונה תאים, רמות שונות של Cdx2, ובהמשך יצירת ICM ו־Trophectoderm.
שאלת הפתיחה היא איך זיגוטה שנראית לכאורה הומוגנית, יכולה לתת (לאחר החלוקה הראשונה) שני בלסטומרים עם פוטנציאל שונה.
ההסבר מתחיל בקומפלקס אימהי שנמצא בביצית עוד לפני שהגנום העוברי פעיל: Subcortical Maternal Complex, או בקיצור SCMC.
Subcortical Maternal Complex - SCMC
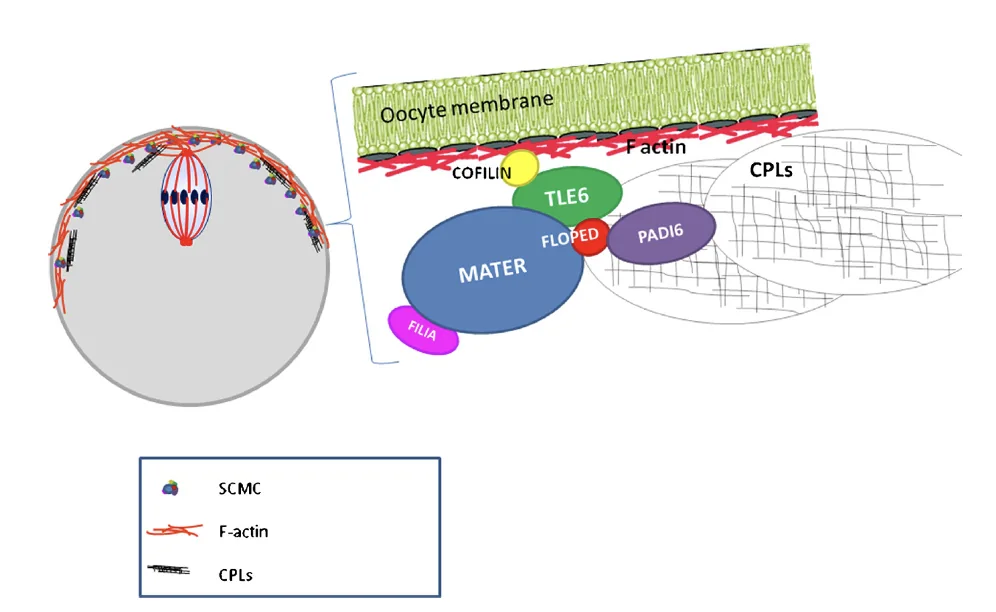
מתחת לממברנה של הביצית נמצאים פילמנטים של F-actin. האקטין הוא חלק מרכזי מהציטוסקלטון, ועוזר לשמור על צורת התא ועל ארגון הממברנה.
צמוד לאזור התת־ממברנלי הזה נמצא קומפלקס חלבוני בשם Subcortical Maternal Complex. הקומפלקסים מופיעים לאורך הממברנה ולא רק בנקודה אחת, ולכן נכון לחשוב עליהם כרשת תת־קורטיקלית שמלווה את קליפת הביצית.
הוא כולל מספר חלבונים, בהם:
| חלבון | תפקיד כללי |
|---|---|
| FLOPED | אחד מחלבוני SCMC; משמש גם כסמן למיקום הקומפלקס |
| TLE6 | חלבון חשוב לשמירת הקומפלקס מאורגן |
| MATER | חלבון נוסף בקומפלקס |
| FILIA | חלבון נוסף בקומפלקס |
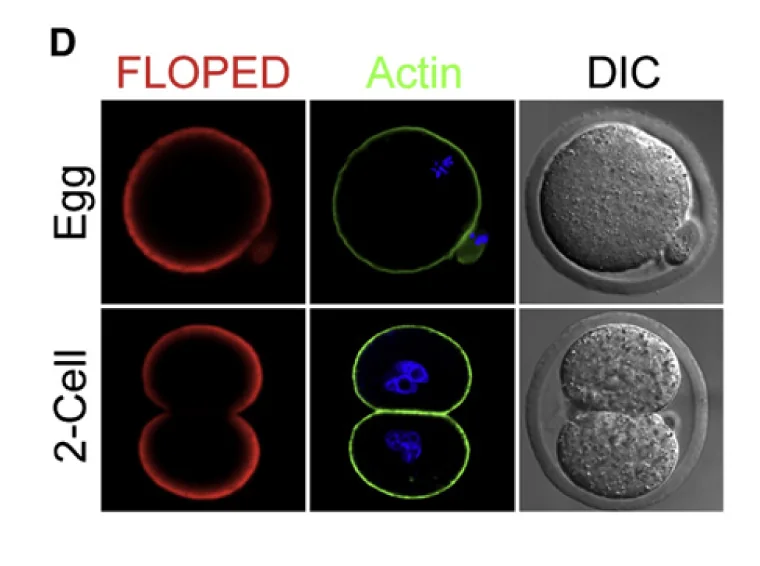
בצביעה ל־FLOPED, רואים שהחלבון נמצא באזור התת־קורטיקלי של הביצית או הזיגוטה, סמוך לממברנה. לפני ההפריה, הביצית נמצאת ב־Metaphase II arrest: יש Polar body אחד, והכרומוזומים של הביצית מסודרים קרוב לממברנה.
לאחר ההפריה והחלוקה הראשונה מתקבלים שני תאים. גם אז FLOPED נמצא מתחת לממברנה, וכמעט לא מופיע באזור המגע בין שני הבלסטומרים.
FLOPED ו־Floped null
מהו null?
Null mutation היא מוטציה שגורמת לחלבון להיות חסר או לא פעיל. לכן Floped null הוא עובר או בעל חיים בלי פעילות תקינה של FLOPED.

בעוברי wild type, החלוקה הראשונה מתרחשת לאורך ציר ה־animal-vegetal, ומתקבלים שני בלסטומרים דומים בגודלם. בהמשך, העוברים מגיעים לשלב המורולה.
במוטנט Floped null החלוקה הראשונה עדיין מתרחשת, אבל שני הבלסטומרים שמתקבלים שונים בגודלם. בהמשך, ההתפתחות נעצרת - בעיקר סביב שלב שני התאים.
המסקנה הישירה היא ש־FLOPED חיוני לשלבים הראשונים של ההתפתחות העוברית, במיוחד סביב החלוקה הראשונה והמעבר שאחריה.
FLOPED הוא אחד מחלבוני ה־SCMC. כשהוא חסר, החלוקה הראשונה יוצרת לרוב שני בלסטומרים בגדלים שונים, וההתפתחות נעצרת מוקדם.
Maternal Effect Genes
Maternal effect gene הוא גן שהתוצר שלו מגיע מהאם דרך הביצית, והוא דרוש להתפתחות העוברית המוקדמת.
בשלבים הראשונים, הגנום של העובר עדיין כבוי. העובר תלוי ב־RNA ובחלבונים שהביצית הכינה ואגרה מראש.
Maternal effect gene הוא גן שהתוצר שלו (RNA או חלבון) מיוצר על ידי האם ומאוחסן בביצית עוד לפני ההפריה, והוא חיוני לשלבי ההתפתחות המוקדמים כשהגנום העוברי כבוי.
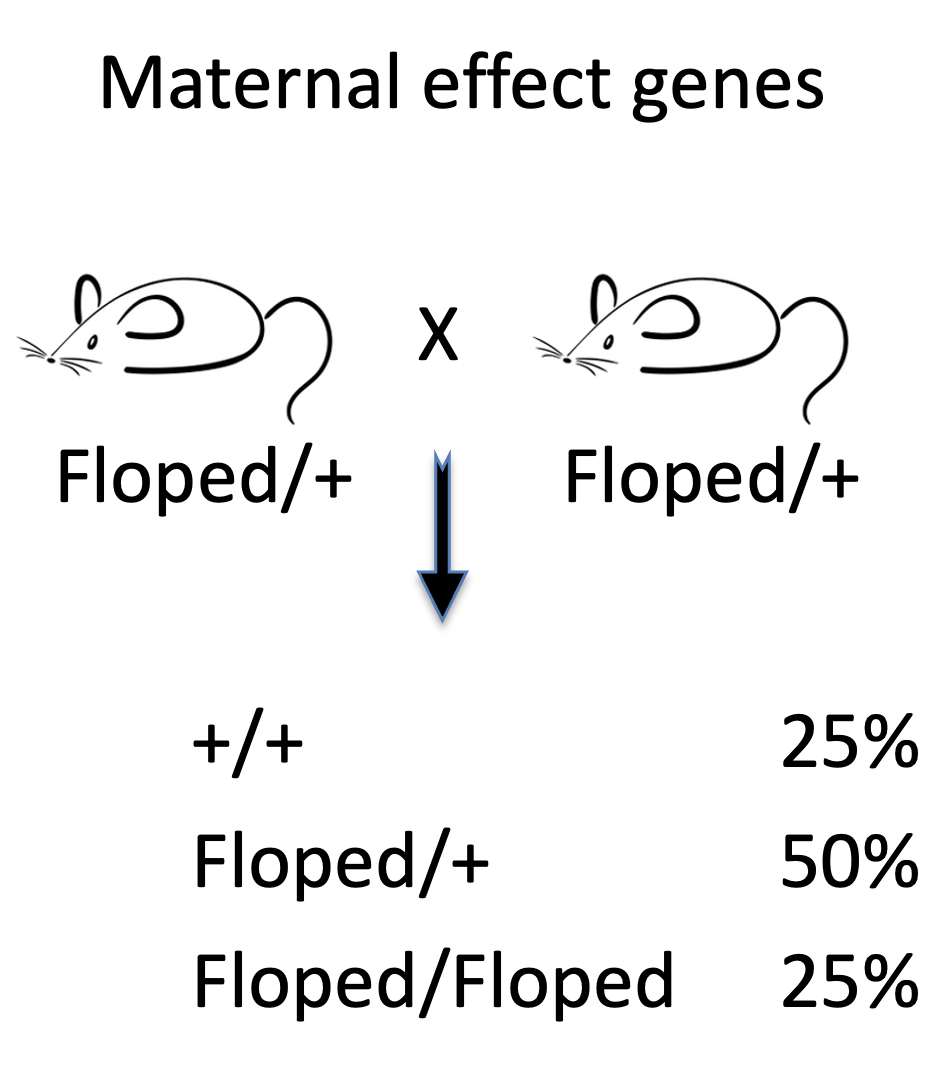
אם זכר ונקבה הטרוזיגוטיים ל־Floped מזדווגים, יש סיכוי של 25% לקבל צאצא הומוזיגוט למוטציה. למרות זאת, צאצא כזה עדיין יכול להיוולד ולהתפתח, כי הביצית של האם ההטרוזיגוטית יכולה להכיל מספיק תוצר אימהי תקין. לכן גם צאצא שמבחינה גנטית מוטנט ל־Floped (Floped/Floped) יכול להיוולד, כל עוד הביצית שקיבלה אותו הכילה תוצר אימהי תקין.
הבעיה מופיעה בדור הבא: נקבה הומוזיגוטית למוטציה יכולה להיוולד (Floped/Floped), אבל הביציות שלה לא יספקו את החלבון הדרוש. העוברים שיתפתחו מביציות כאלה ייעצרו בשלבים הראשונים.
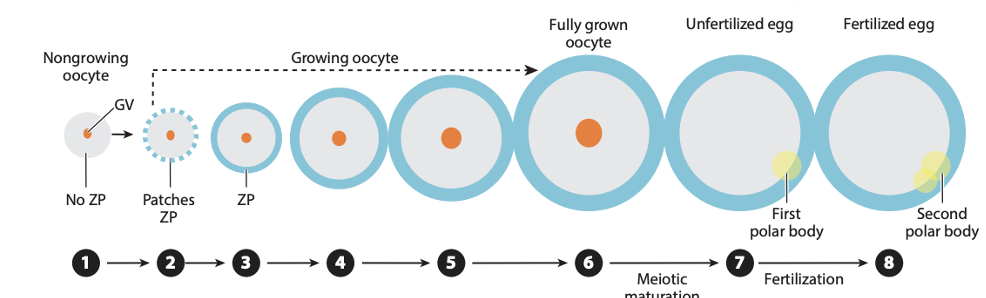
העיקרון:
- הביצית גדלה עוד לפני ההפריה.
- בזמן הגדילה היא מייצרת RNA וחלבונים, כולל חלבוני SCMC.
- לאחר ההבשלה המיוטית, פעילות הגנום בביצית מושתקת.
- עד Zygotic genome activation (ZGA), ההתפתחות נשענת על התוצרים שהביצית הביאה.
בעכבר, ZGA מתרחש סביב שלב שני התאים. לכן חלבונים שפועלים לפני כן חייבים להגיע מהביצית.
TLE6 ותפקיד הקומפלקס
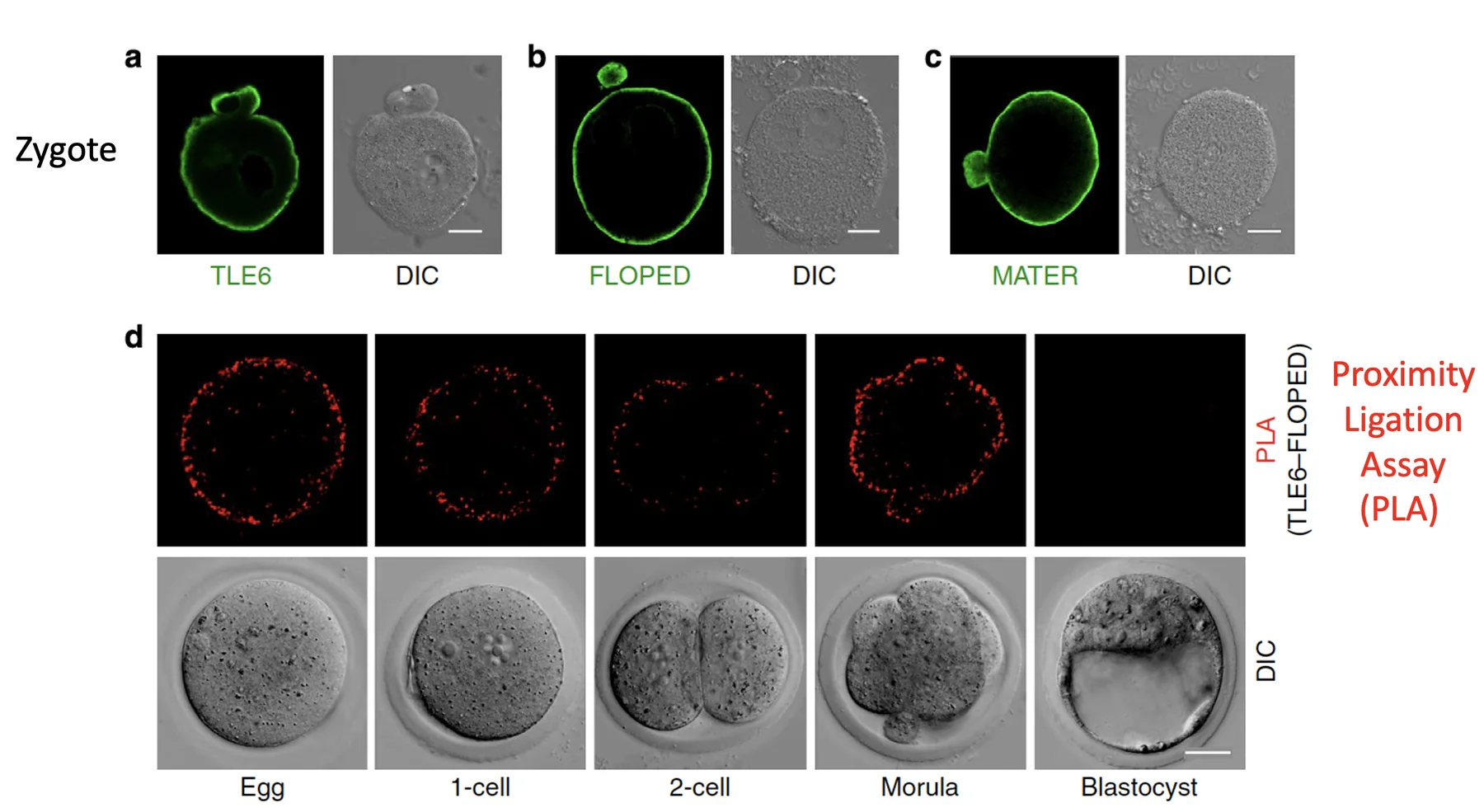
צביעות לחלבוני SCMC מראות שהם נמצאים סמוך לממברנה בשלבים המוקדמים. בניסוי שבו בודקים קרבה בין TLE6 לבין FLOPED, מתקבלת פלואורסצנציה רק כששני החלבונים צמודים זה לזה. כך אפשר להראות שהם נמצאים יחד בקומפלקס.
הקומפלקס קיים בביצית, בזיגוטה ובשלבי cleavage מוקדמים. אחרי המורולה הוא כבר כמעט לא נראה, ולכן תפקידו רלוונטי בעיקר לשלבים הראשונים.
Tle6 null

כשפוגעים ב־TLE6 מתקבל פנוטיפ דומה לזה של Floped null: החלוקה הראשונה מתרחשת, אבל שני הבלסטומרים שונים בגודלם, והמשך ההתפתחות נפגע. העוברים לא מצליחים להתקדם מעבר לשלב שני התאים, בעוד שעוברי wild type כבר ממשיכים למורולה ובהמשך לבלסטוציסט.
בניסוי נוסף שבו פגעו גם ב־MATER התקבלה תוצאה דומה. לכן המסקנה אינה רק ש־TLE6 חשוב בפני עצמו, אלא שה־SCMC כקומפלקס שלם דרוש לחלוקה ראשונה סימטרית.
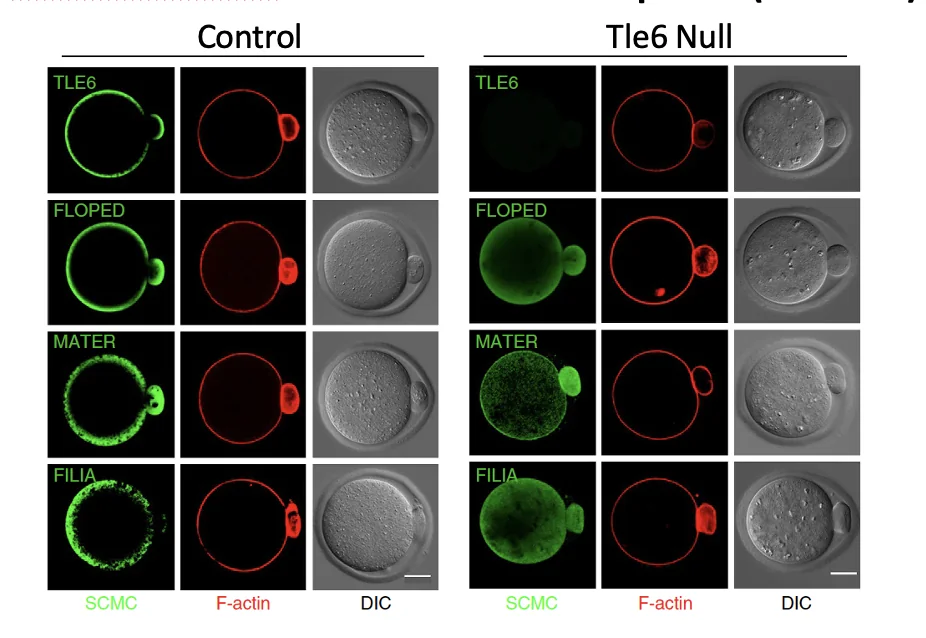
ניתן לראות שהאקטין יושב יפה מתחת לממברנה.
בצביעה לחלבוני הקומפלקס רואים שב־Tle6 null החלבונים האחרים עדיין קיימים, אבל הם מפוזרים בתא במקום להתרכז יחד כקומפלקס מסודר.
לכן TLE6 חשוב לארגון הקומפלקס. בלי TLE6, החלבונים נמצאים, אבל ה־SCMC מאבד את הארגון שלו כיחידה פעילה.
מיקום הכרומוזומים וחלוקה לשני תאים שווים
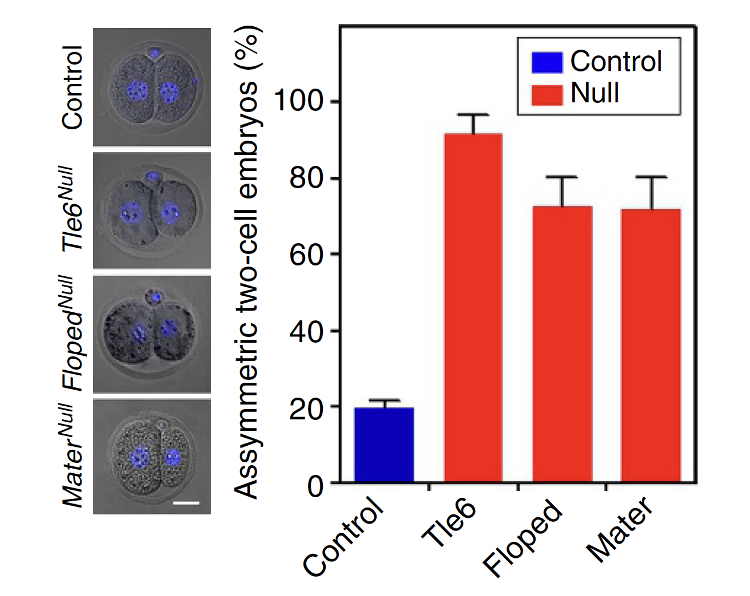
כדי למדוד אם החלוקה הראשונה סימטרית, משווים בין גודל שני הבלסטומרים שנוצרו. יחס קרוב ל־1 אומר ששני התאים דומים בגודלם.
בניסוי הוגדרה חלוקה א-סימטרית כשההבדל בגודל הבלסטומרים גדול מ־10%. זה סף שרירותי, אבל הוא מאפשר להשוות בין wild type לבין מוטנטים.
במוטנטים של חלבוני SCMC יש שיעור גבוה של חלוקות א-סימטריות. לכן תפקיד מרכזי של הקומפלקס הוא לייצר חלוקה ראשונה לשני תאים דומים בגודלם.
SCMC מבטיח חלוקה ראשונה סימטרית בגודלה. כפי שיוסבר בהמשך, הוא מסייע למקם את הכרומוזומים קרוב למרכז התא לאחר איחוי שני הגרעינים בזיגוטה. פגיעה בקומפלקס גורמת לסטייה של הכרומוזומים מהמרכז.
האם זה חשוב? כנראה שכן, מפני שראינו שאם הם לא זהים בגודלם התהליך לא ממשיך.
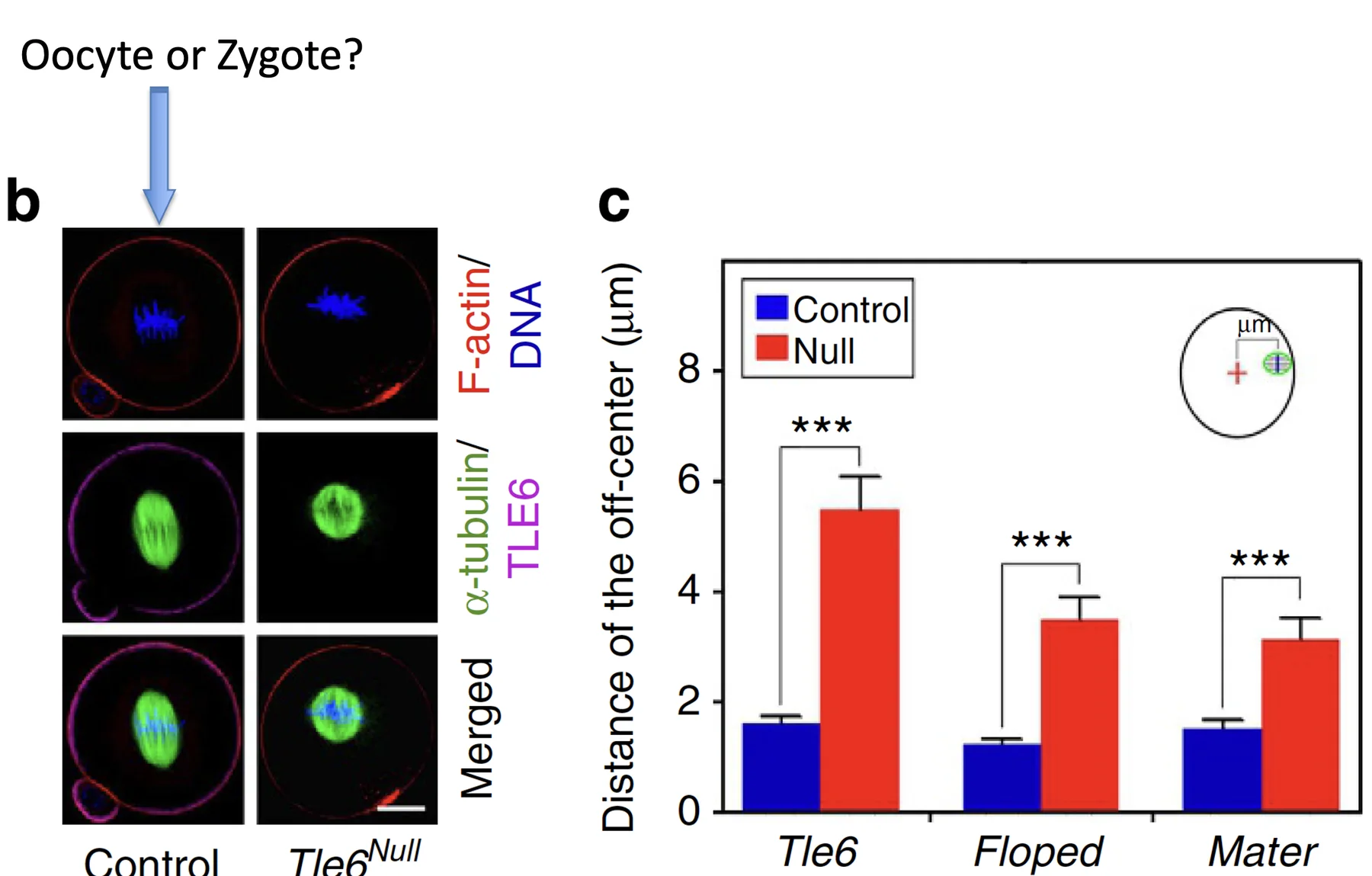
בתמונה למעלה נראית זיגוטה.
כדי לקבל שני תאים שווים, הכרומוזומים צריכים להימצא קרוב למרכז התא לפני החלוקה. בזיגוטה תקינה, אחרי איחוי שני הגרעינים, הכרומוזומים מתארגנים במרכז. במוטנטים ל־SCMC רואים סטייה: הכרומוזומים רחוקים יותר ממרכז הזיגוטה.
המדידה נעשית כך:
- מגדירים את נקודת האמצע של הזיגוטה
- מודדים את המרחק של הכרומוזומים מנקודת האמצע
- משווים בין wild type לבין מוטנטים
ב־wild type הכרומוזומים קרובים יותר למרכז. במוטנטים של TLE6, FLOPED או MATER יש סטייה גדולה יותר. הסטייה הזאת מסבירה איך מתקבלים שני בלסטומרים בגדלים שונים.
מודל חלוקת ה־RNA בביצית
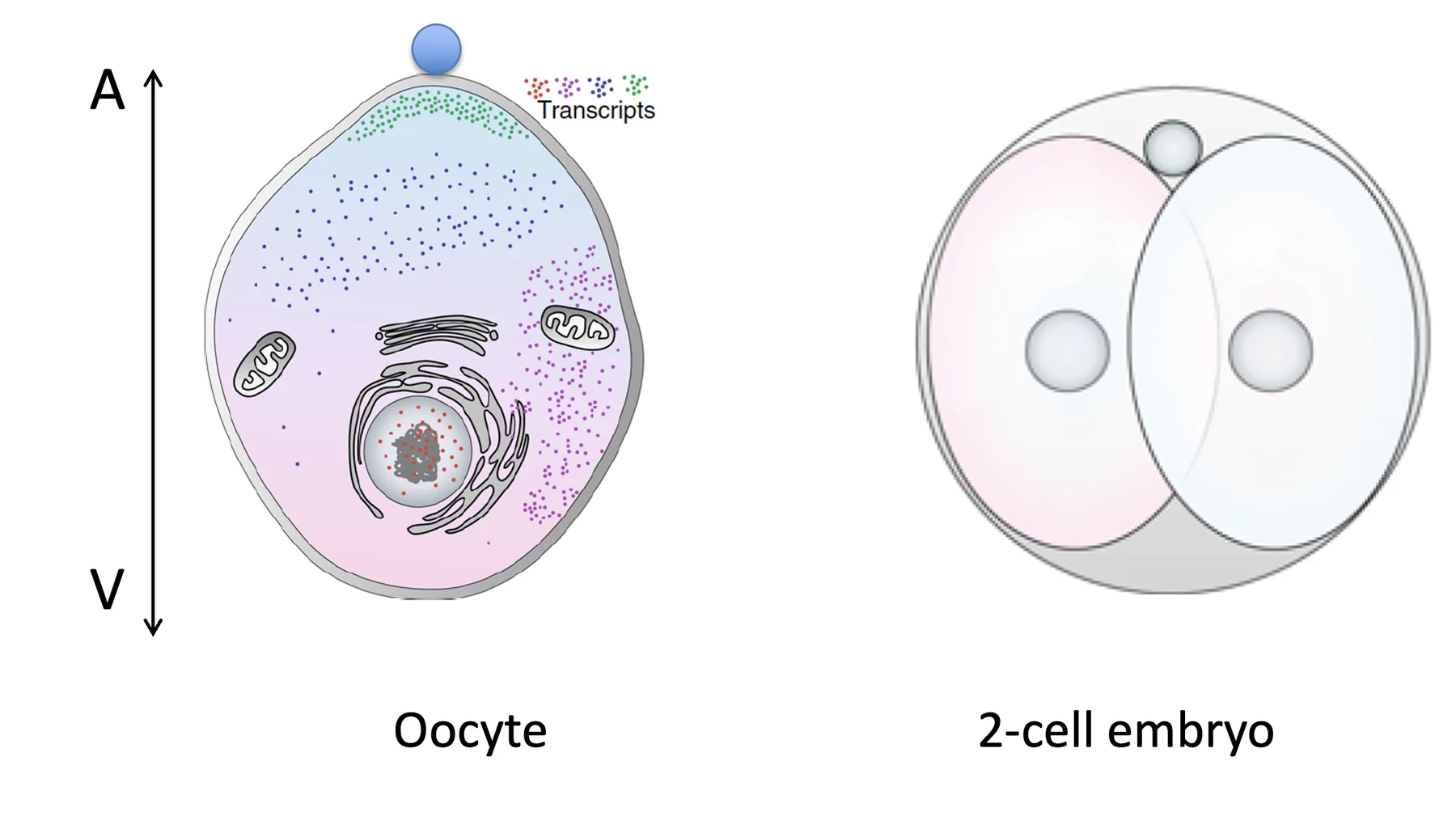
הטרנסקריפט האדום נמצא רק בגרעין, ולכן במודל הוא מייצג RNA גרעיני, כנראה non-coding RNA. הטרנסקריפט הסגול נמצא בצד אחד של התא, והטרנסקריפט הירוק נמצא בצד ה־animal, סמוך לממברנה.
המודל שמוצג כאן מנסה להסביר איך ביצית שנראית הומוגנית יכולה ליצור שני תאים שונים לאחר החלוקה הראשונה. זה מודל הסברי של השיעור, ולא הוכחה מלאה למנגנון אחד ויחיד.
הנקודה הבטוחה במודל היא זו: בביצית יכולים להיות טרנסקריפטים שממוקמים באזורים שונים. כלומר, ה־RNA בביצית מפוזר באופן לא אחיד.
באיור מופיעים כמה טרנסקריפטים לדוגמה:
| טרנסקריפט באיור | מיקום אפשרי | משמעות במודל |
|---|---|---|
| אדום | בגרעין | RNA גרעיני; במודל הוא מייצג non-coding RNA |
| סגול | בצד אחד של התא | יכול ליצור הטיה בין שני הבלסטומרים |
| ירוק | בצד animal, קרוב לממברנה | צריך להתחלק בצורה מאוזנת בין שני התאים |
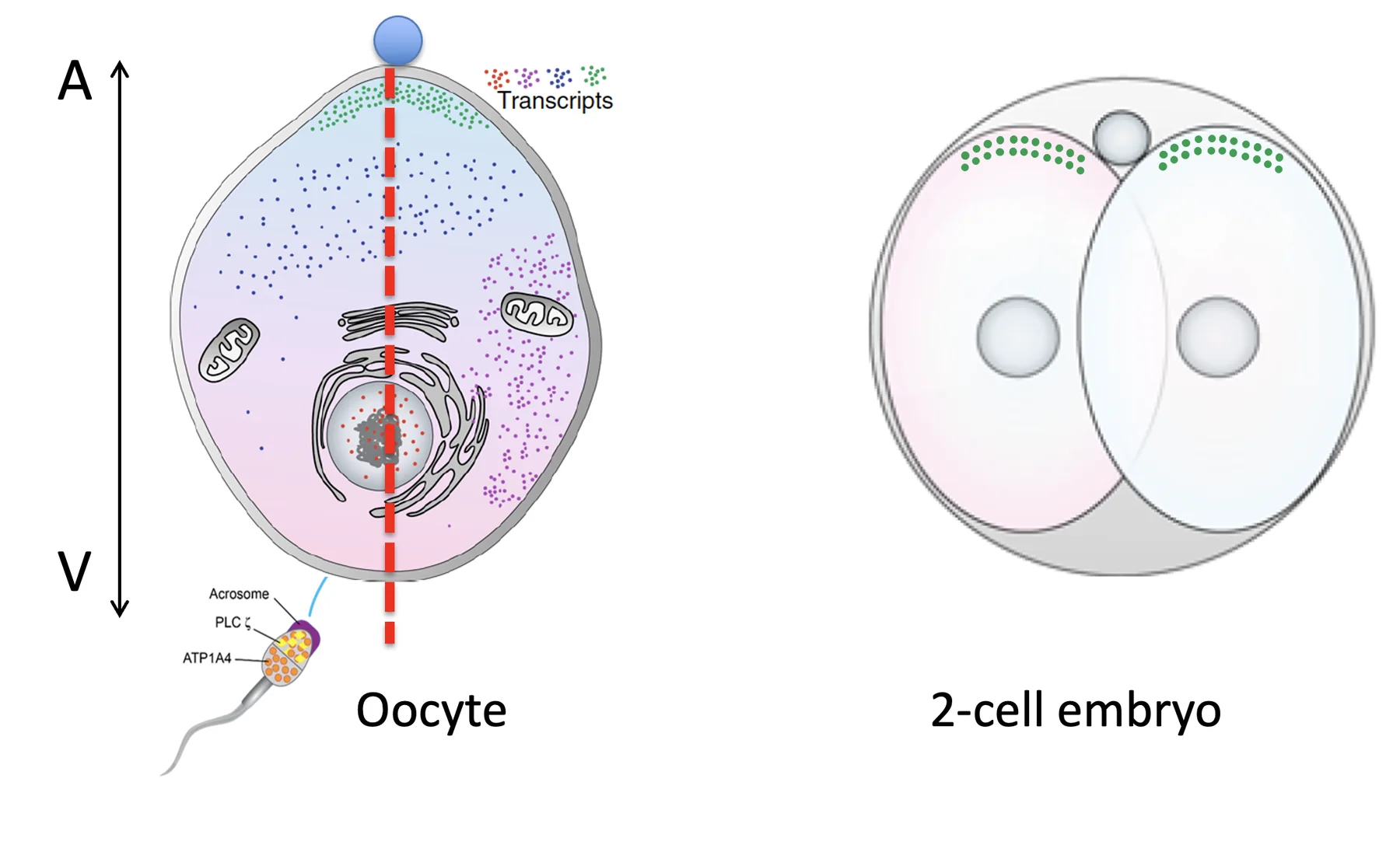
במצב תקין, ה־SCMC מסייע למקם את הכרומוזומים במרכז ולכוון את מישור החלוקה לאורך הציר animal-vegetal. כך מתקבלים שני תאים דומים בגודלם.
אם יש טרנסקריפט חיוני שצריך להגיע לשני התאים בכמות דומה (למשל הנקודות הירוקות בחלק השמאלי של האיור), חלוקה סימטרית מאפשרת לפזר אותו בצורה מאוזנת. כל תא מקבל מינון מתאים ויכול להמשיך להתחלק.
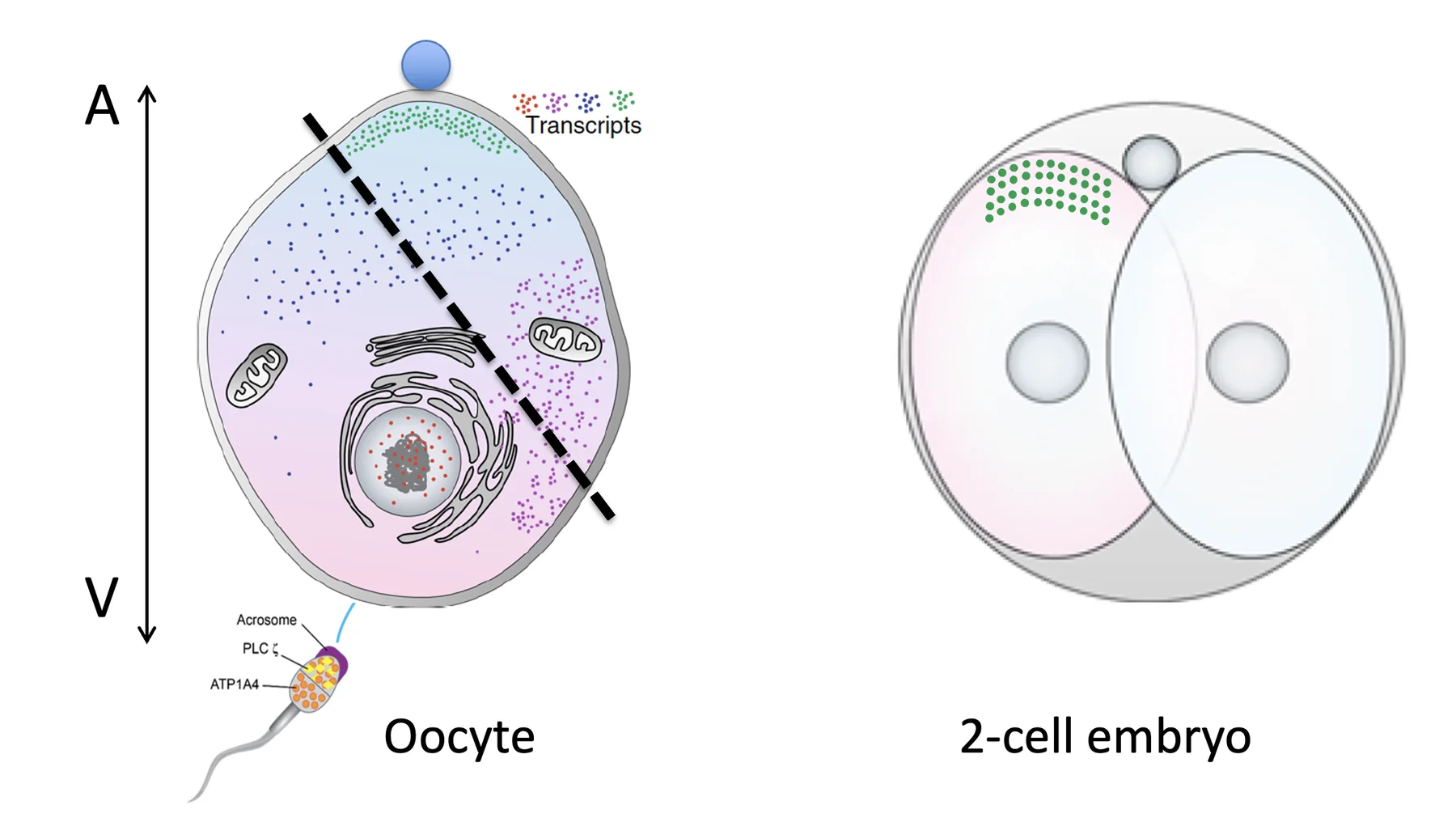
כש־SCMC פגוע, מישור החלוקה זז מהמרכז, ומתקבלים שני תאים בגדלים שונים. במצב כזה גם טרנסקריפטים מסוימים יכולים להתחלק בצורה לא מאוזנת (למשל הנקודות הירוקות, שכעת נמצאות רק בצאצא השמאלי).
המודל מפריד בין שני סוגי טרנסקריפטים:
| סוג טרנסקריפט | תוצאה אפשרית של חלוקה לא מאוזנת |
|---|---|
| טרנסקריפט חיוני במינון מדויק | פגיעה בהמשך החלוקות, עד עצירה מוקדמת |
| טרנסקריפט שמטה תוכנית התפתחותית | יצירת הבדל פרובביליסטי בין שני הבלסטומרים |
כך אפשר לחבר בין שני דברים שנראו בשיעור הקודם:
פיזור לא הומוגני של RNA בביצית
↓
חלוקה ראשונה בכיוון מסוים
↓
שני בלסטומרים עם רמות שונות של טרנסקריפטים מסוימים
↓
הבדלים מוקדמים, למשל ברמות LincGET
↓
Carm1 / H3R26me / Sox2 / Nanog
↓
רמות שונות של Cdx2
↓
First cell fate decision
זה מודל של הטיה פרובביליסטית. הוא לא מתאר נעילה מוחלטת של גורל התא, אלא דרך להתחיל הבדל קטן שמערכת רגולטיבית יכולה בהמשך להגביר או לתקן.
עכבר לעומת אדם: התפתחות מוקדמת
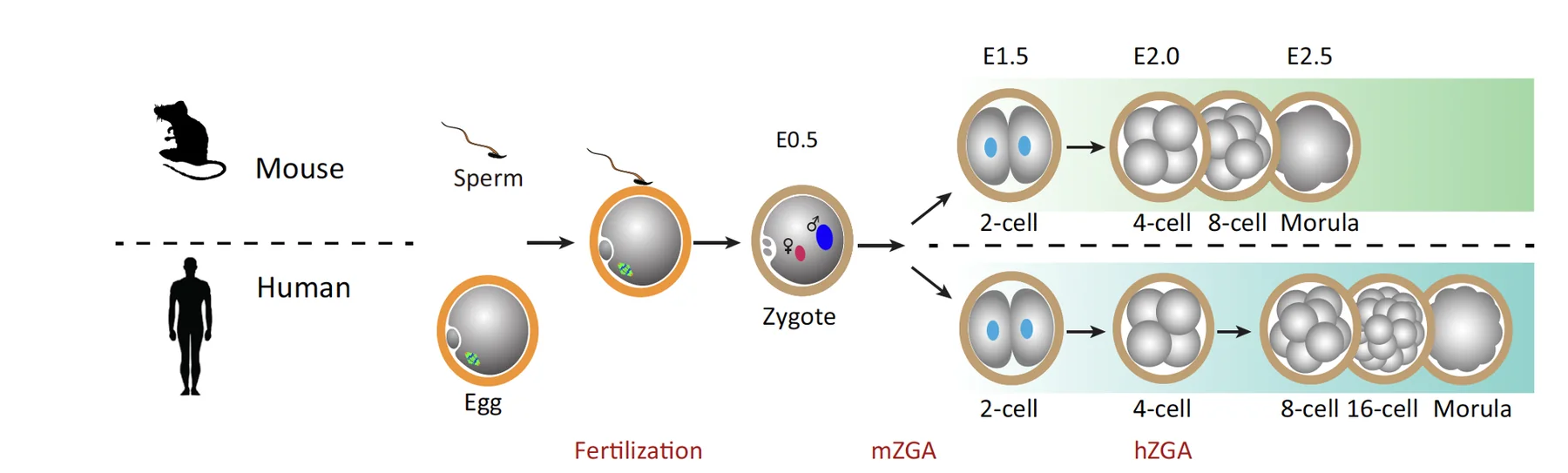
העקרונות דומים בעכבר ובאדם, אבל יש הבדלים בזמנים ובאנטומיה.
| תהליך | עכבר | אדם |
|---|---|---|
| הפריה | ביצית ב־Metaphase II, כניסת זרע, איחוי גרעינים | אותו עיקרון |
| Zygotic genome activation | סביב שלב שני התאים | סביב שלב ארבעת התאים |
| תלות ב־maternal RNA | קצרה יותר | ארוכה יותר, כי ZGA מאוחר יותר |
| Compaction / polarization | סביב המורולה המוקדמת | במורולה מתקדמת יותר, סביב 16 תאים |
הבדלים כאלה חשובים כשמשווים ניסויים בעכברים להתפתחות אדם. הם משנים את הזמנים ואת המבנה, אבל העיקרון הכללי של התפתחות רגולטיבית נשמר.
In Vitro Fertilization - IVF
IVF הוא תהליך שבו מוציאים ביציות מהשחלה לאחר טיפול הורמונלי, מפגישים אותן עם תאי זרע מחוץ לגוף, ומאפשרים התחלה של התפתחות עוברית בתרבית.
יש שתי גישות עיקריות:
| שיטה | מה עושים? | מתי משתמשים? |
|---|---|---|
| Conventional / Standard IVF | שמים ביצית ותאי זרע בצלחת, והזרע מפרה את הביצית בעצמו | כשאין בעיה בולטת בזיהוי הביצית או בתאי הזרע |
| ICSI Intracytoplasmic sperm injection | בוחרים תא זרע אחד ומזריקים אותו ישירות לתוך הביצית | כשיש בעיה שמונעת מהזרע לזהות או לחדור לביצית, או כשיש ספירת זרע נמוכה מאוד / תנועתיות נמוכה |
ב־ICSI מחקים באופן מכני את השלב שבו הזרע נכנס לתוך הביצית. אם יש מעט מאוד תאי זרע תקינים, אפשר לבחור תא שנראה חי ותנועתי ולהזריק אותו.
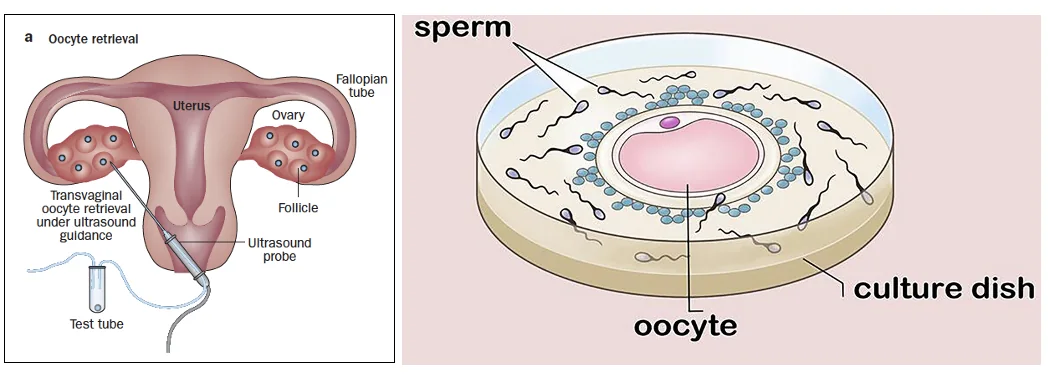
בתמונה למעלה מימין רואים IVF סטנדרטי, ומשמאל ICSI.
PGD ו־PGS
PGD - Preimplantation Genetic Diagnosis הוא אבחון גנטי של עוברים לפני השרשה. הוא מתבצע כחלק מתהליך IVF, משום שצריך לקבל עוברים מחוץ לגוף כדי לבדוק אותם.
PGD מתאים כשידועה מוטציה מסוימת במשפחה. לדוגמה, אם ידוע שההורים נשאים או שיש במשפחה מחלה עם מוטציה מוכרת, אפשר לבדוק כל עובר לפני ההחזרה לרחם. ב־PGD לא מחפשים באופן פתוח את כל הגנום, אלא בודקים מראש את המוטציה או המחלה הידועה. את העוברים שנמצאו תקינים אפשר להחזיר לרחם או להקפיא.
PGS - Preimplantation Genetic Screening הוא סקר רחב יותר שמכוון לבעיות כרומוזומליות, למשל aneuploidy. זה רלוונטי במיוחד כשיש הפלות חוזרות, אובדני היריון בשלבים מאוחרים יותר, או חשד להפרעות במספר הכרומוזומים בלי מוטציה ספציפית ידועה.
ההבדל העיקרי:
| מושג | מה מחפשים? |
|---|---|
| PGD | מוטציה ידועה או מחלה גנטית ידועה |
| PGS | הפרעות כרומוזומליות רחבות, למשל טריזומיות או aneuploidy |
שיטות דגימה ב־PGD
דגימה בשלב ה־Cleavage
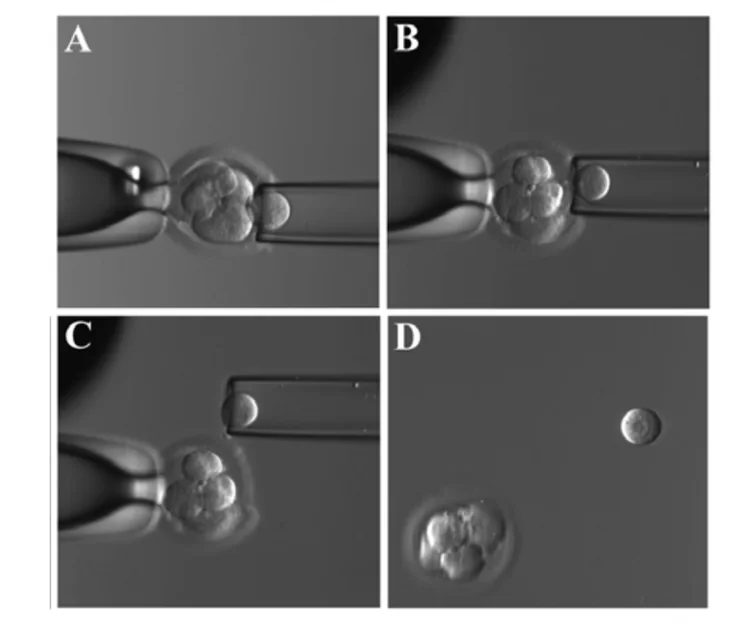
בשלב ה־cleavage, לפני ה־compaction, אפשר להחזיק את העובר בעזרת ואקום, ליצור חור קטן ב־Zona Pellucida, ולשלוף בלסטומר אחד או שניים.
העובר יכול להמשיך להתפתח משום שבשלבי ה־cleavage המערכת עדיין רגולטיבית. אם הבדיקה תקינה, נותנים לעובר להמשיך להתפתח לבלסטוציסט ואז מחזירים אותו לרחם או מקפיאים אותו. עם זאת, הוצאת תא מהעובר היא מניפולציה, ולכן חשוב לזכור שגם אם לכאורה מתקבל בלסטוציסט תקין, לא תמיד אפשר לדעת מה יהיו ההשפעות ארוכות הטווח.
דגימה בשלב הבלסטוציסט
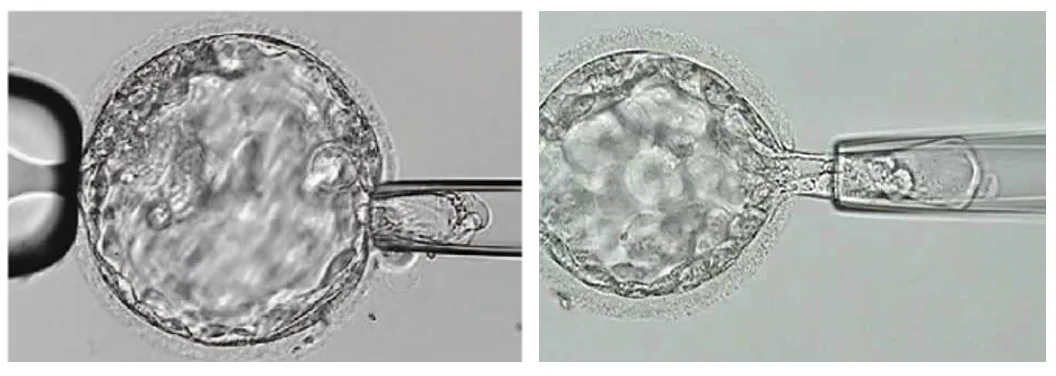
בשלב הבלסטוציסט ניתן לקחת כמה תאים מה־Trophectoderm. היתרון הוא שמקבלים יותר DNA לבדיקה, והפגיעה בעובר קטנה יותר ביחס להוצאה של חלק גדול מתאי ה־cleavage.
דגימת Polar Bodies
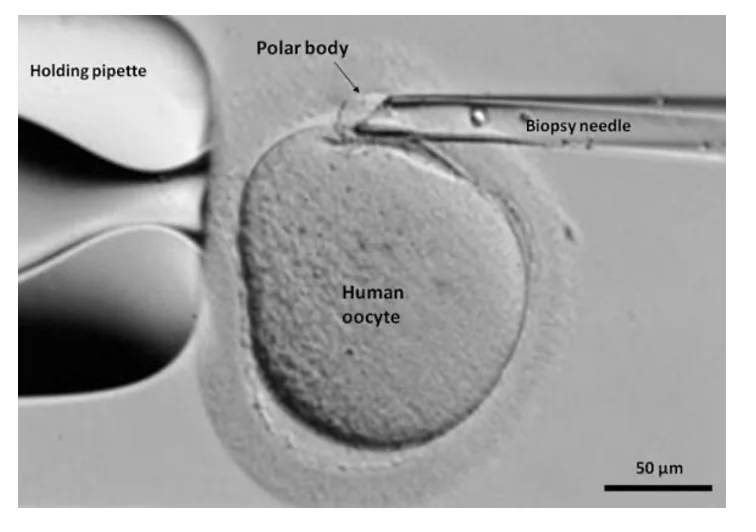
אפשר לבדוק גם את הגופיפים הפולריים של הביצית. היתרון הוא שמתחילים לבדוק עוד לפני ההפריה או ממש סמוך לה. אפשר לבדוק את ה־Polar body הראשון לפני יצירת הזיגוטה, ובשלב הזיגוטה אפשר לעיתים לבדוק גם את הגופיף הפולרי השני.
סיכום שיטות הדגימה
| שלב התפתחותי | ביצית לפני/סביב הפריה | עובר בשלב Cleavage | בלסטוציסט יום 5-7 |
|---|---|---|---|
| טכניקה | דגימת הגופיפים הפולריים 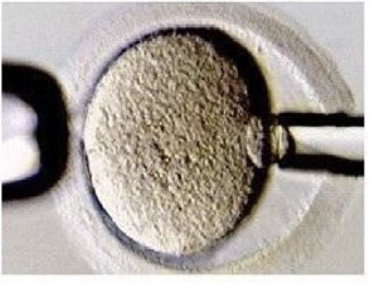 | דגימת בלסטומרים מהעובר 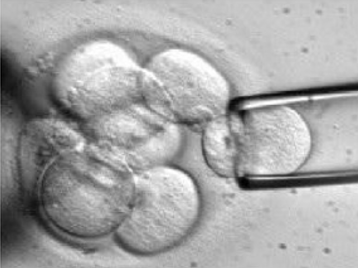 | דגימת תאי TE מהבלסטוציסט 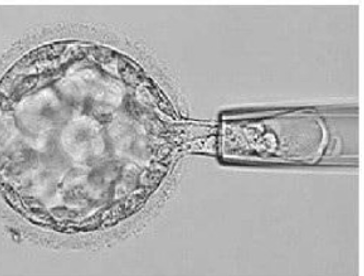 |
| דגימה | הגופיפים הפולריים הראשון והשני | 1 או 2 בלסטומרים | 5-10 תאי TE |
| יתרון מרכזי | מתחילים בדיקה מוקדם מאוד | שיטה ותיקה ומבוססת | יותר DNA לבדיקה ואפשרות טובה יותר לזהות מוזאיציזם |
שיטות בדיקה גנטית
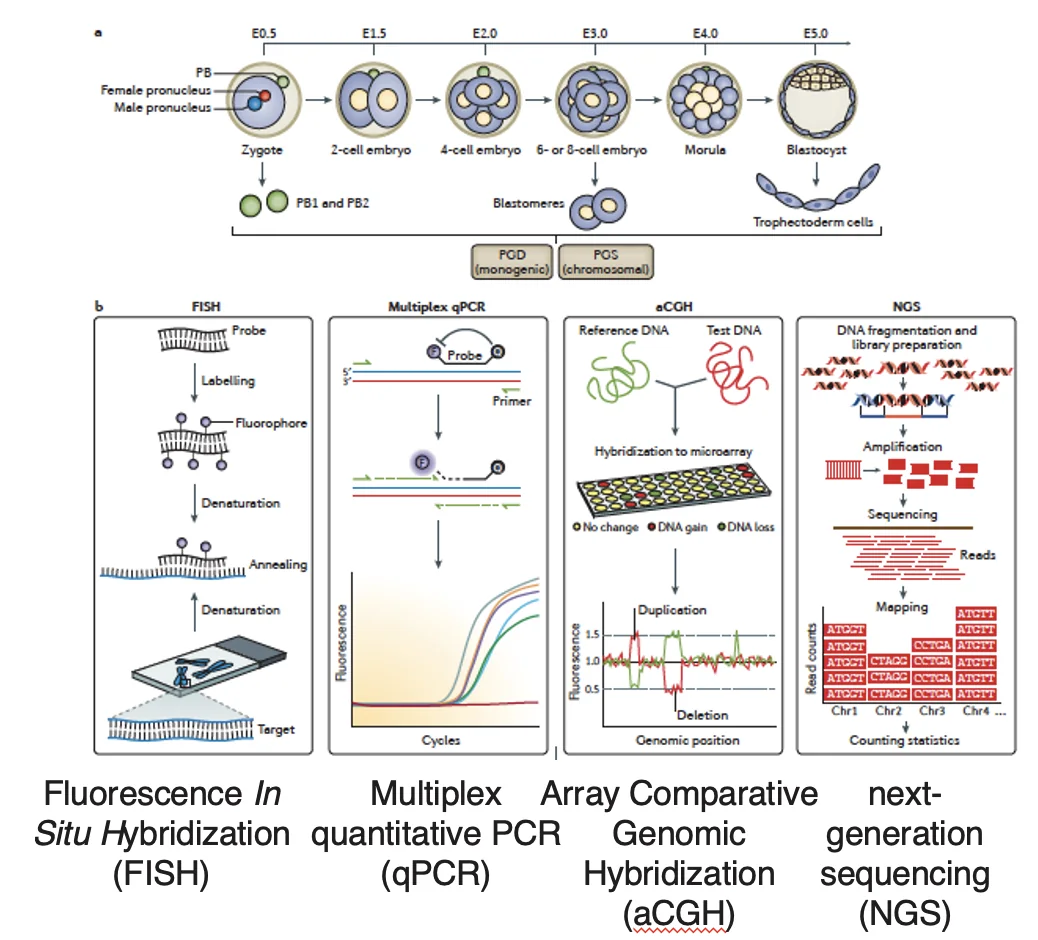
לאחר שלוקחים תא או מספר תאים, אפשר לבצע כמה סוגי בדיקות:
| שיטה | שימוש כללי |
|---|---|
| NGS | ריצוף רחב של DNA; מתאים גם לסקר רחב וגם לזיהוי מוטציות, בהתאם לתכנון הבדיקה |
| aCGH | זיהוי שינויים כרומוזומליים רחבים יותר |
| RT-PCR / qPCR | בדיקה ממוקדת של מוטציה או רצף ידוע, לפי תכנון הבדיקה |
| FISH | זיהוי כרומוזומים או אזורים כרומוזומליים בעזרת פרובים פלואורסצנטיים |
ב־FISH אפשר לזהות למשל טריזומיות, משום שרואים מספר עותקים לא תקין של כרומוזום מסוים.
הנקודה המעשית היא שהבדיקה הגנטית קובעת איזה עובר יוחזר לרחם או יוקפא להמשך שימוש.
השרשה: Apposition, Adhesion, Invasion
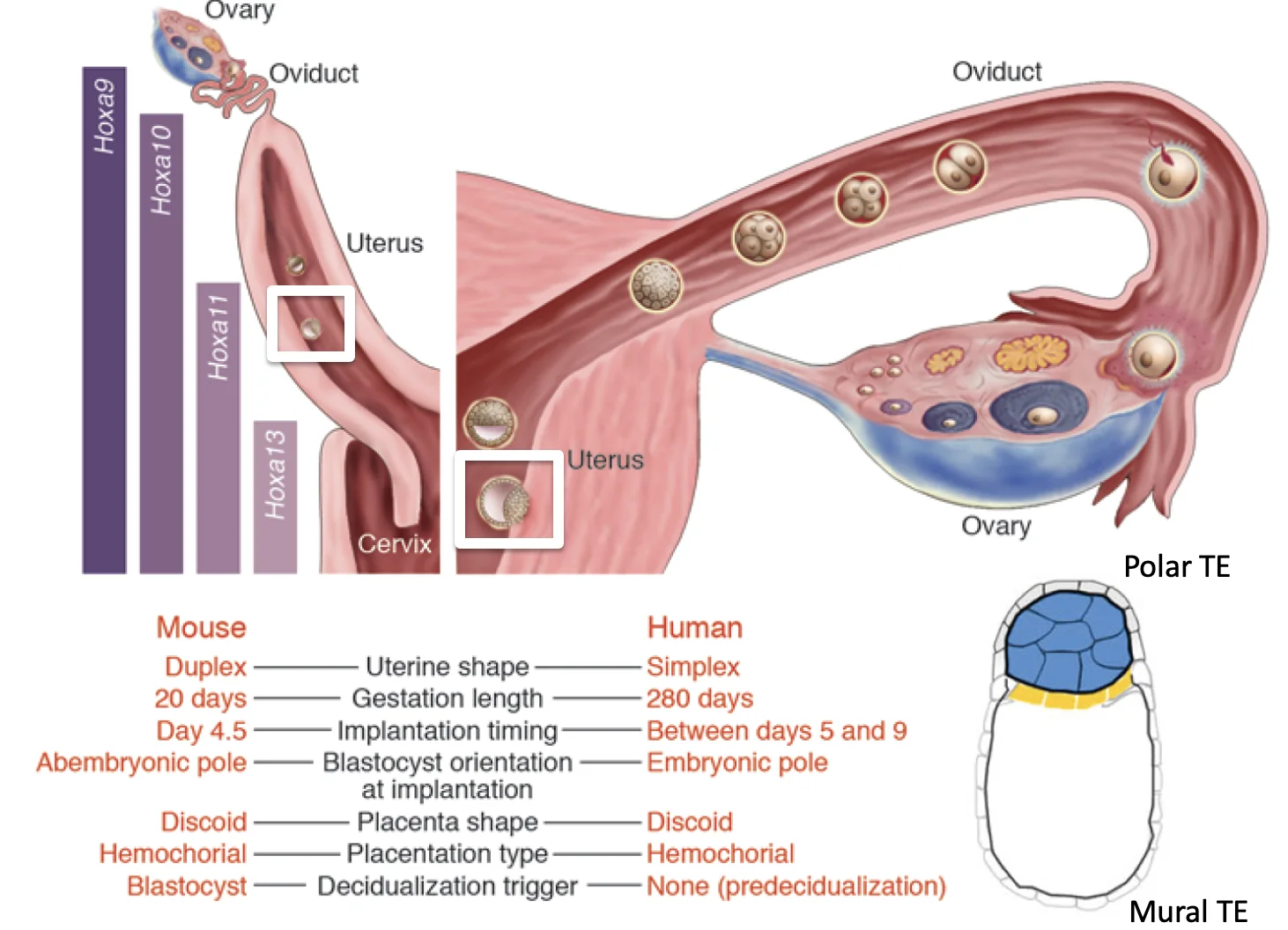
השרשה (implantation) מתחילה אחרי שהבלסטוציסט בוקע מה־Zona Pellucida ומגיע לרחם. קודם לכן, בביוץ הביצית יוצאת מהשחלה אל החצוצרה, שם מתרחשת ההפריה. בזמן הנדידה לכיוון הרחם העובר ממשיך להתחלק, ולכן הוא מגיע לרחם כבר כבלסטוציסט.
ברקע האנטומי, מערכת המין הנקבית מתפתחת מה־Müllerian ducts. באדם יש איחוי נרחב של שני הצינורות ולכן נוצר רחם עם חלל יחיד, ואילו בעכבר האיחוי חלקי יותר ולכן מתקבל רחם דו־קרני עם צוואר רחם משותף. ההבדל הזה חשוב להבנת ההשרשה בעכבר לעומת אדם.
באדם ההשרשה מתחילה דרך הצד הפולרי של הבלסטוציסט (polar trophectoderm), כלומר הצד שבו נמצא ה־ICM. בעכבר ההידבקות הראשונית מתרחשת בעיקר דרך ה־mural trophectoderm.
שלושת השלבים המרכזיים:
| שלב | משמעות |
|---|---|
| Apposition | הבלסטוציסט מתמקם מול רירית הרחם באזור מתאים |
| Adhesion | הבלסטוציסט נצמד לאפיתל של הרחם |
| Invasion | תאי העובר פולשים לרירית הרחם |
השרשה מוצלחת דורשת שני תנאים בו־זמנית:
- בלסטוציסט קומפטנטי - עובר שמסוגל להידבק ולפלוש
- רחם רצפטיבי - רירית רחם שנמצאת בחלון הזמן המתאים לקליטה
השלבים האלה מבוססים על תקשורת מולקולרית בין הבלסטוציסט לבין הרחם.
Human Endometrium
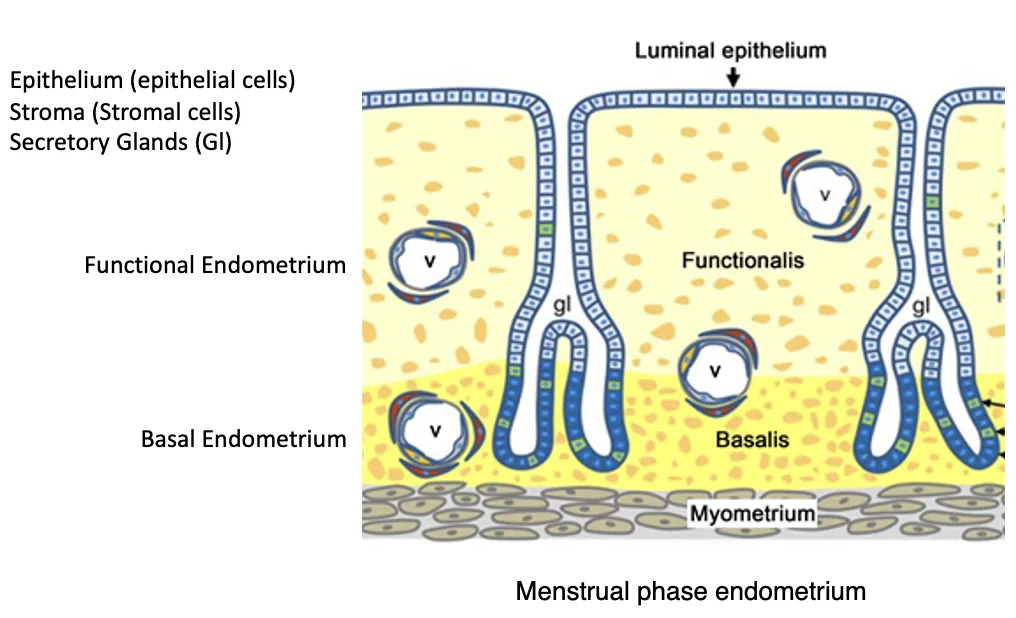
דופן הרחם באדם בנויה משלוש שכבות עיקריות: endometrium בחלק הפנימי, myometrium כשכבת השריר, ו־perimetrium כקרום חיצוני.
ה־endometrium הוא רירית הרחם. מבחוץ לו נמצאת שכבת השריר, ה־myometrium, ומחוץ לה ה־perimetrium.
ברירית הרחם מבחינים בין שני אזורים:
| אזור | תפקיד |
|---|---|
| Basal endometrium / basalis | החלק הקבוע שנשאר גם אחרי הווסת |
| Functional endometrium / functionalis | החלק שנבנה מחדש בכל מחזור ונושר בזמן הווסת |
חלל הרחם מצופה ב־luminal epithelium, שכבת אפיתל חד־שכבתית שמפרידה בין הלומן לבין הרירית. בתוך הרירית יש גם stromal cells, כלי דם ובלוטות הפרשה. בלוטות ההפרשה הן למעשה התקפלויות של האפיתל פנימה אל תוך האנדומטריום, והן נפתחות אל חלל הרחם.
במהלך הווסת ה־functionalis נושר. ה־basalis נשאר, ומתוכו הרירית נבנית מחדש במחזור הבא.
המחזור החודשי והמחזור השחלתי
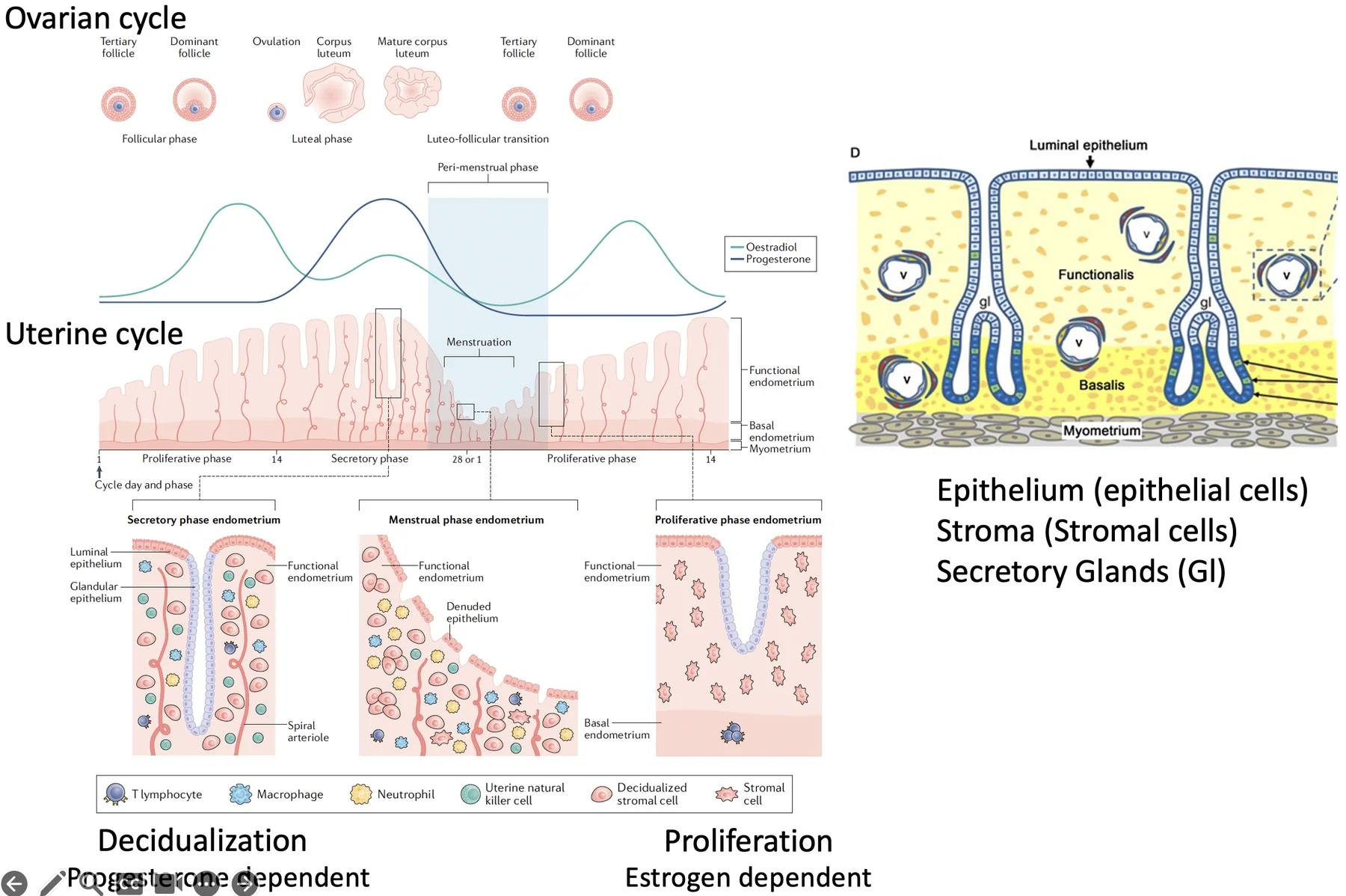
יש שני מחזורים שמתרחשים במקביל ומסונכרנים דרך הורמוני המין:
- Uterine cycle - השינויים ברירית הרחם
- Ovarian cycle - התפתחות הזקיק בשחלה, ביוץ ויצירת corpus luteum
Uterine Cycle
| שלב | מה קורה ברירית הרחם? | הורמון מרכזי |
|---|---|---|
| Menstruation | ה־functionalis נושר | ירידה בהורמונים |
| Proliferative phase | ה־basalis בונה מחדש את ה־functionalis; תאי אפיתל וסטרומה מתחלקים | אסטרוגן |
| Secretory phase | תאי הסטרומה עוברים התמיינות, והרירית נעשית מתאימה יותר לקליטת עובר | פרוגסטרון |
תאי סטרומה - תאים שמרכיבים את הרקמה התומכת ברירית הרחם. הם משתנים במהלך המחזור, במיוחד בשלב הסקרטורי, כדי להכין את הרירית לקליטת העובר. מוזמנים להעמיק בויקיפדיה על משתית.
Ovarian Cycle
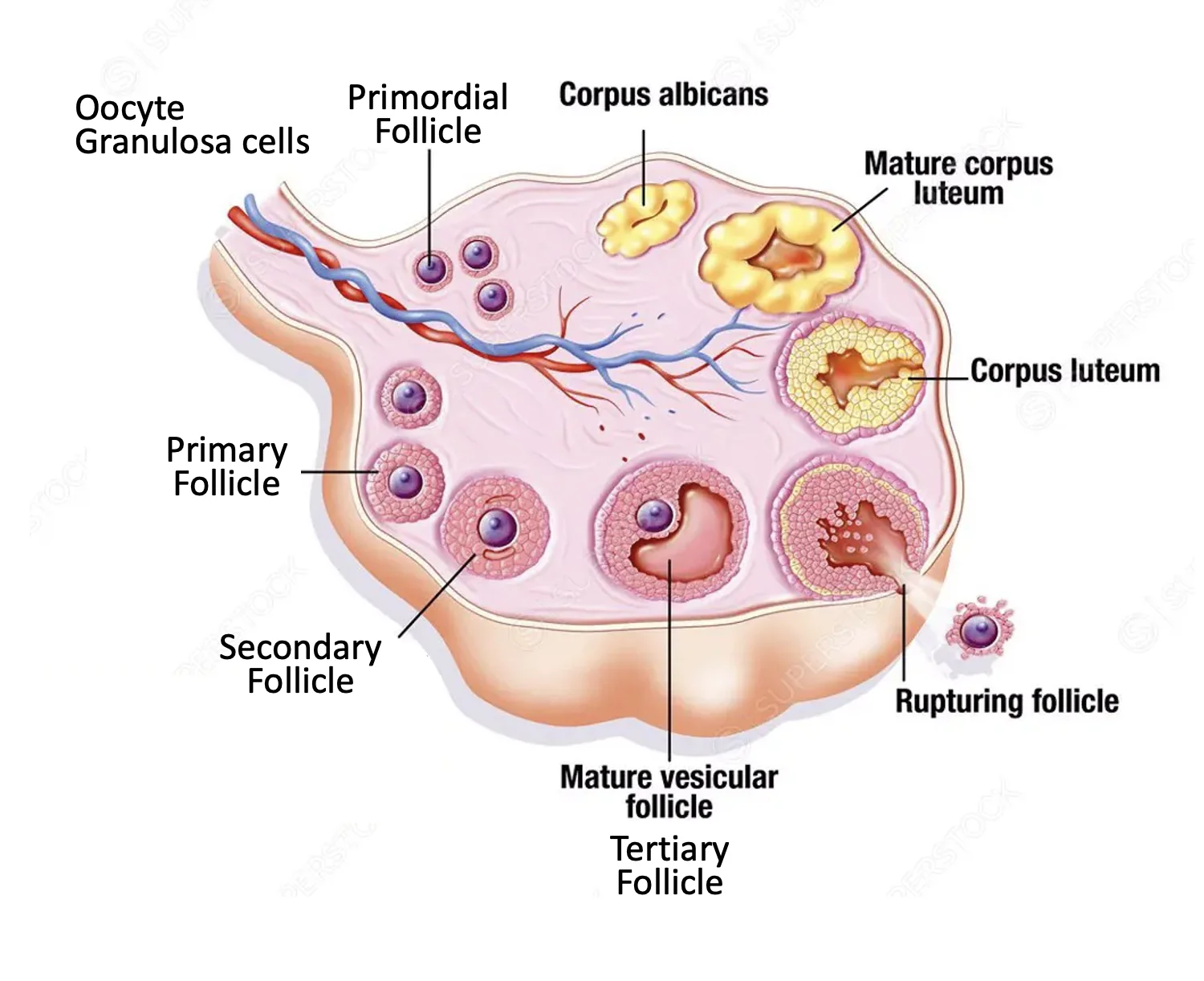
המחזור השחלתי מתאר את ההתפתחות של הזקיק והביצית בשחלה, החל מהשלב העוברי ועד הביוץ ומה שקורה אחריו.
Phase 1 - לפני הלידה: יצירת המאגר השחלתי
במהלך ההתפתחות העוברית, תאי ה־PGC נודדים אל הגונדה המתפתחת. בנקבות הם מגיעים לשחלה, עוברים חלוקות מיטוטיות רבות, ובהמשך מתפתחים לביציות ראשוניות.
בשלב העוברי מספר התאים מגיע לשיא של בערך 7 מיליון, אך לקראת הלידה חלק גדול מהם מתנוון. בזמן הלידה נשארות בערך 2 מיליון ביציות.
בשלב זה הביציות כבר נכנסו למיוזה I, אבל נעצרו ב־Prophase I arrest. סביב כל ביצית יש שכבה של תאי granulosa, וכך נוצר primordial follicle.
כלומר:
PGC
↓
oogonia / primary oocytes
↓
Prophase I arrest
↓
Primordial follicle
Phase 2 - מהלידה עד הבגרות המינית
מהלידה ועד הבגרות המינית אין התקדמות משמעותית במיוזה. הביציות נשארות ב־Prophase I arrest בתוך ה־primordial follicles.
עם זאת, המאגר ממשיך להצטמצם: עד הבגרות המינית נשארות בערך 300,000-200,000 ביציות בכל שחלה.
Phase 3 - Follicular phase: התפתחות הזקיק עד הביוץ
החל מהבגרות המינית, בהשפעת הציר ההורמונלי:
Hypothalamus
↓ GnRH
Pituitary
↓ FSH + LH
Ovary
ההיפותלמוס מפריש GnRH, שמשפיע על ההיפופיזה. ההיפופיזה מפרישה את שני ההורמונים המרכזיים למחזור השחלתי:
- FSH - Follicle Stimulating Hormone
- LH - Luteinizing Hormone
בהשפעת ההורמונים האלה זקיקים מתחילים להתפתח:
Primordial follicle
↓
Primary follicle
↓
Secondary follicle
↓
Tertiary / mature follicle
↓
Ovulation
בשלב ה־primordial follicle אין עדיין Zona Pellucida. במעבר ל־primary follicle, הביצית גדלה, מייצרת RNA וחלבונים, ונוצרת סביבה ה־Zona Pellucida.
בדרך כלל כמה זקיקים מתחילים את התהליך, אבל רק אחד מהם הופך לזקיק דומיננטי. שאר הזקיקים מתנוונים. הזקיק הדומיננטי ממשיך להתפתח עד ל־tertiary / mature follicle ולביוץ.
במהלך ה־follicular phase תאי ה־granulosa מפרישים בעיקר estrogen. האסטרוגן חשוב גם למחזור הרחמי, כי הוא מעודד פרוליפרציה של רירית הרחם.
Phase 4 - Ovulation: ביוץ והשלמת מיוזה I
הביוץ תלוי ב־LH surge, כלומר עלייה חדה ב־LH.
בעקבות העלייה ב־LH, הביצית הראשונית משלימה את מיוזה I ונוצרת secondary oocyte. הביצית השניונית מתחילה מיוזה II, אבל נעצרת שוב - הפעם ב־Metaphase II arrest.
זה השלב שבו הביצית יוצאת מהזקיק ומהשחלה:
Primary oocyte
↓ completes meiosis I
Secondary oocyte
↓ starts meiosis II
Metaphase II arrest
↓
Ovulation
לכן הביצית שיוצאת בביוץ היא secondary oocyte שנמצאת ב־Metaphase II arrest.
Phase 5 - Luteal phase: אחרי הביוץ
אחרי הביוץ, שארית הזקיק הופכת ל־corpus luteum.
ה־corpus luteum מפריש בעיקר progesterone, וגם מעט estrogen. הפרוגסטרון חשוב להכנת רירית הרחם לקליטת העובר, בעיקר דרך תהליך של decidualization.
אם לא מתרחשת הפריה ולא מתחיל היריון, ה־corpus luteum מתנוון בהדרגה והופך ל־corpus albicans. בעקבות ההתנוונות שלו רמות הפרוגסטרון יורדות, וה־functionalis של רירית הרחם נושר בזמן הווסת.
סיכום קצר
לפני הלידה:
PGC → primary oocyte → Prophase I arrest → primordial follicle
מהלידה עד הבגרות:
אין המשך מיוזה, אבל יש ירידה במספר הביציות
Follicular phase:
FSH + LH → follicle growth → estrogen secretion
Ovulation:
LH surge → completion of meiosis I → secondary oocyte in Metaphase II arrest
Luteal phase:
Corpus luteum → progesterone → decidualization
If no pregnancy → corpus albicans → progesterone drops → menstruation
המחזור השחלתי והמחזור הרחמי מסונכרנים דרך ההורמונים השחלתיים: estrogen בעיקר לפני הביוץ (לפרולפרציה, מאפשר לרירית הרחם לגדול), ו־progesterone בעיקר אחרי הביוץ (בעיקר ל־deciudalization, הכנת הרירית לקליטת העובר).
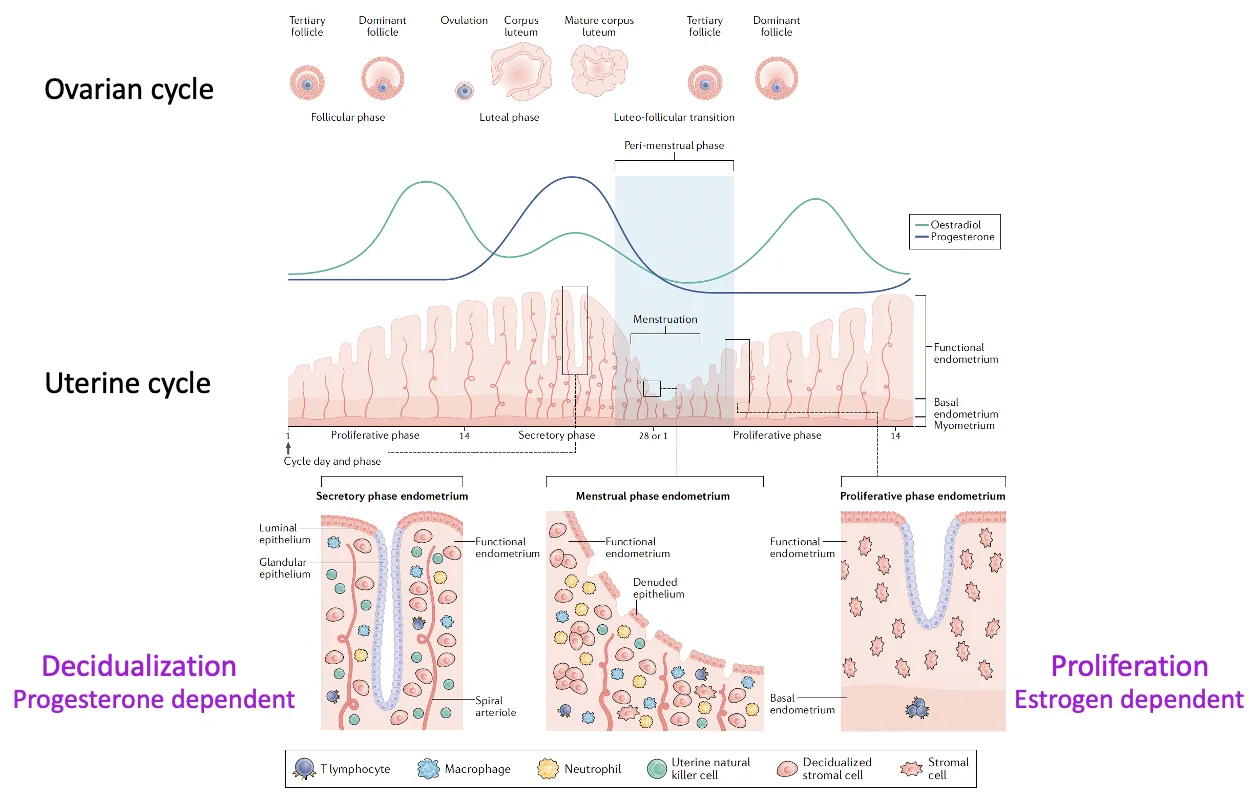
האסטרוגן תומך בבנייה מחדש של רירית הרחם. הפרוגסטרון תומך ב־decidualization ובהכנת הרירית לקליטת העובר.
Decidualization
Decidualization (מילולית, כנראה קשור למילה הלטינית deciduus = נושר / נופל / עתיד לנשור) הוא תהליך תלוי פרוגסטרון שבו תאי הסטרומה של רירית הרחם משתנים ומתאימים את הרירית לתמיכה בהיריון.
במהלך התהליך תאי הסטרומה עוברים שינוי צורה: מתאים מאורכים דמויי פיברובלסטים לתאים עגולים יותר, עם תכונות שמתאימות לקליטה ולתמיכה בעובר.
באדם, decidualization מתרחשת באופן ספונטני במחזור החודשי, גם בלי נוכחות עובר. אם אין השרשה, רמות הפרוגסטרון יורדות, והרירית שהוכנה נושרת בווסת. לכן אפשר לקשור בין decidualization לבין עצם היכולת לקבל וסת: בבעלי חיים ללא וסת, התהליך בדרך כלל לא מתרחש באופן ספונטני.
בעכבר, decidualization תלויה גם בהידבקות של העובר לרחם. adhesion של העובר הוא חלק מהאות שמפעיל את התהליך. בתהליך הזה יכולה להופיע גם polyploidization, בעיקר בעכברים: התא מכיל יותר משתי ערכות כרומוזומים מלאות, משום שאין ציטוקינזיס רגיל.
הערת הקשר: Endometriosis
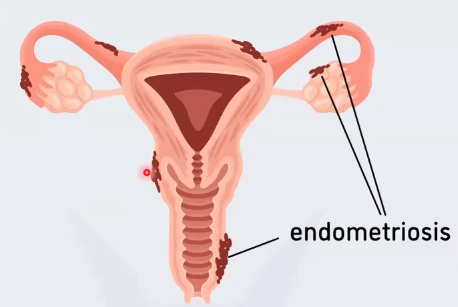
Endometriosis הוא מצב שבו רקמת endometrium נמצאת מחוץ לחלל הרחם, למשל על פני אזורים בחלל הבטן או סביב הרחם. הרקמה הזאת יכולה להתנהג כמו רירית הרחם: לעבור שינויים מחזוריים, לדמם, וליצור תסמינים כמו כאב, דלקת והידבקויות.
אחת ההיפותזות שהוזכרה היא שבזמן נשירת רירית הרחם, חלק מהרקמה נע בכיוון הפוך דרך החצוצרות אל חלל הבטן, ושם הוא יכול להיצמד ולהמשיך להגיב למחזור ההורמונלי.
חלון ההשרשה ואקטיבציית הבלסטוציסט
בעכבר ובאדם יש הבדלים אנטומיים והורמונליים, אבל העיקרון דומה: השרשה מתרחשת רק כשהבלסטוציסט מוכן והרחם רצפטיבי.
בעכברים לא קיימת ההשלה של ה־functionalis. מכל מקום, מבחינת ההשרשה ההבדלים בעכבר ובאדם אינם מהותיים.
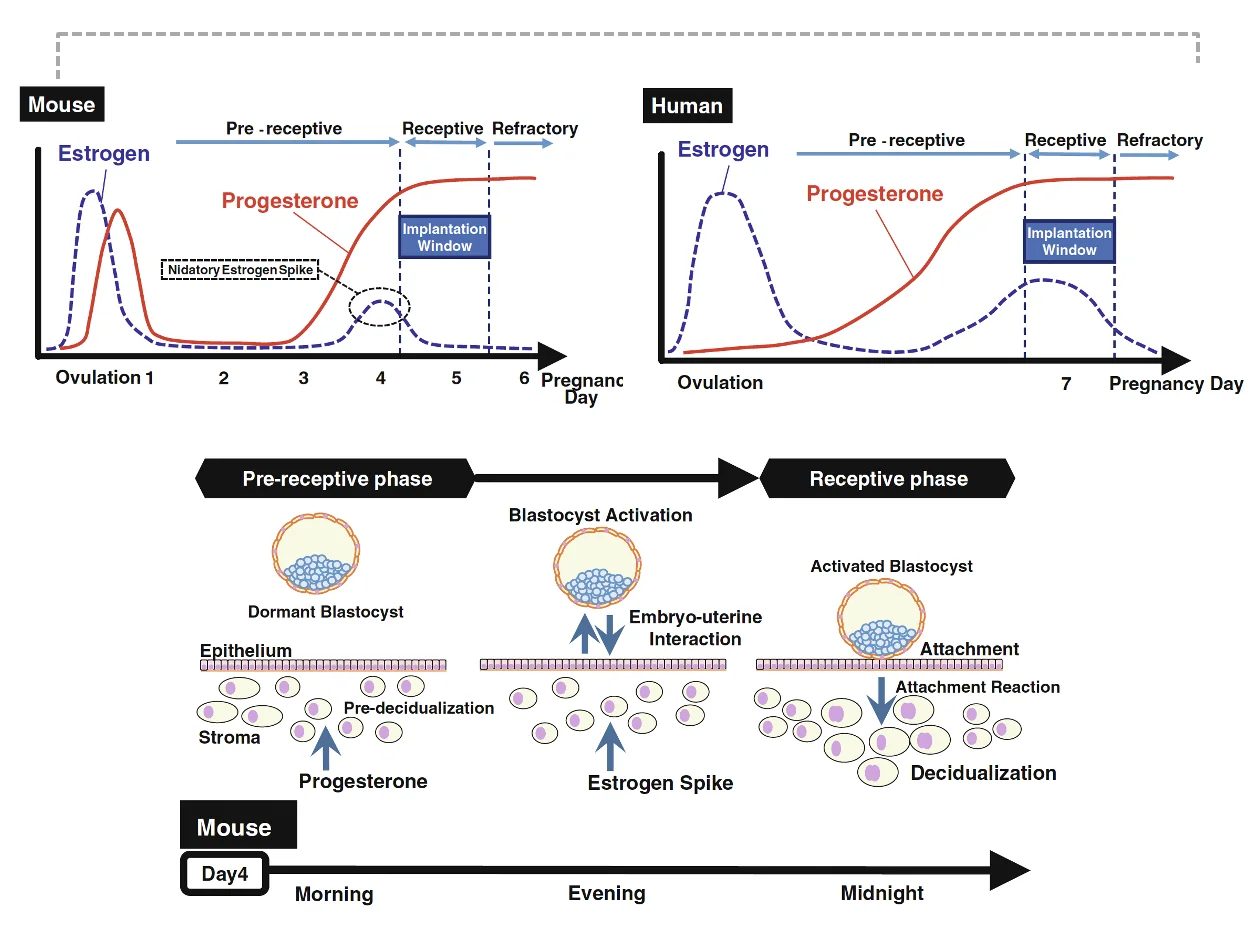
יש שלב pre-receptive, שבו נוכחות עובר לבדה לא מספיקה להשרשה. בהמשך מופיע חלון receptive, שבו רמות פרוגסטרון גבוהות יחד עם פיק נוסף של אסטרוגן מאפשרים השרשה.
הפיק של האסטרוגן מפעיל את הבלסטוציסט. האקטיבציה הזאת מאפשרת לבלסטוציסט להתחיל אינטראקציה פעילה עם האפיתל של הרחם.
השרשה דורשת שילוב:
Progesterone → decidualization / הכנת הרירית
Estrogen peak → blastocyst activation
Blastocyst activation + receptive uterus → implantation window
היצמדות הבלסטוציסט (Blastocyst Adhesion)
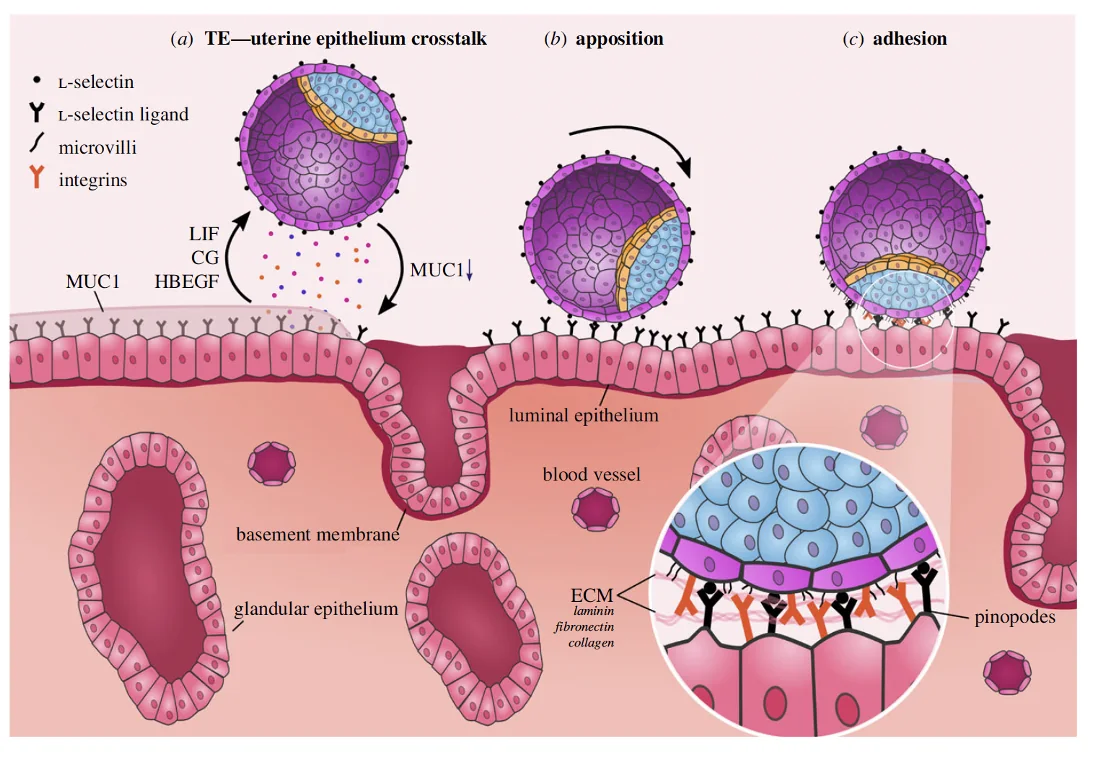
לפני ההיצמדות, בלוטות ברירית הרחם מפרישות מוקוס (ריר). המוקוס מגן על הרירית, אבל גם מקשה על הבלסטוציסט להיצמד ישירות לאפיתל.
בזמן האקטיבציה יש ירידה ב־Mucin-1, מרכיב חשוב במוקוס. כך הבלסטוציסט יכול ליצור מגע קרוב יותר עם תאי האפיתל.
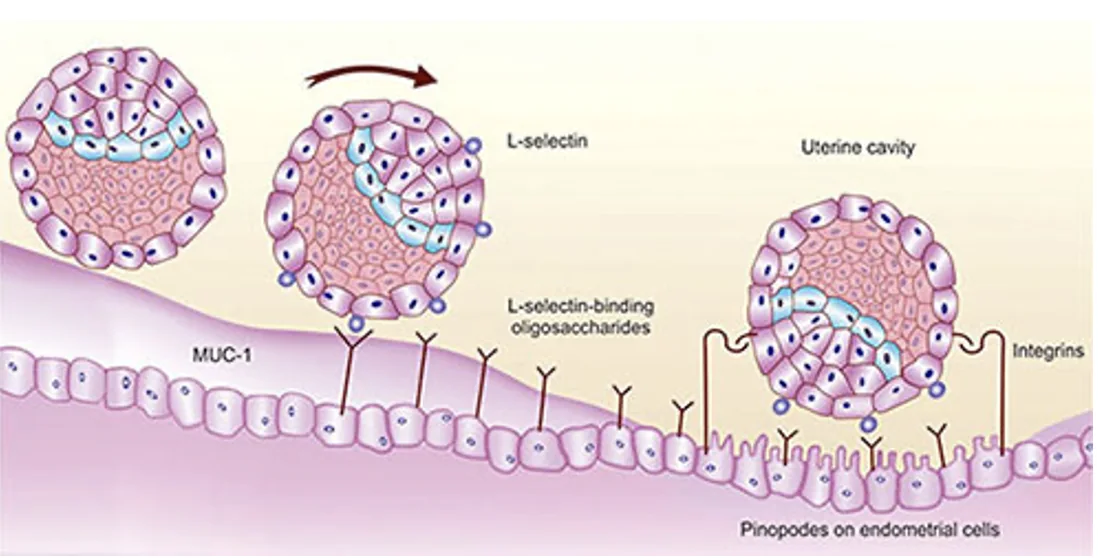
בשלב ה־apposition וה־adhesion משתתפות מולקולות הידבקות. באדם, שלב ה־rolling עוזר לבלסטוציסט להתמקם כך שהצד הפולרי שלו פונה לעבר רירית הרחם, ורק אז ההידבקות מתחזקת.
| מולקולה | תפקיד כללי |
|---|---|
| L-selectin | חלבון טראנס־ממברנלי שנמצא על ה־Trophectoderm ומשתתף בשלב rolling ראשוני מול ליגנדים סוכריים ברירית הרחם |
| סוכרים על תאי הרחם | משמשים כליגנדים לאינטראקציות ראשוניות (ליגנדים ל־L-selectin) |
| Integrins | חלבונים טראנס־ממברנלים שנמצאים על תאי האפיתל ויש להם ליגנד על ה־Trophectoderm. משתתפים בהידבקות יציבה יותר בין הבלסטוציסט לרירית |
השלב הזה מוביל מהתמקמות רופפת יחסית להידבקות חזקה יותר של הבלסטוציסט לרירית הרחם.
Blastocyst Invasion באדם
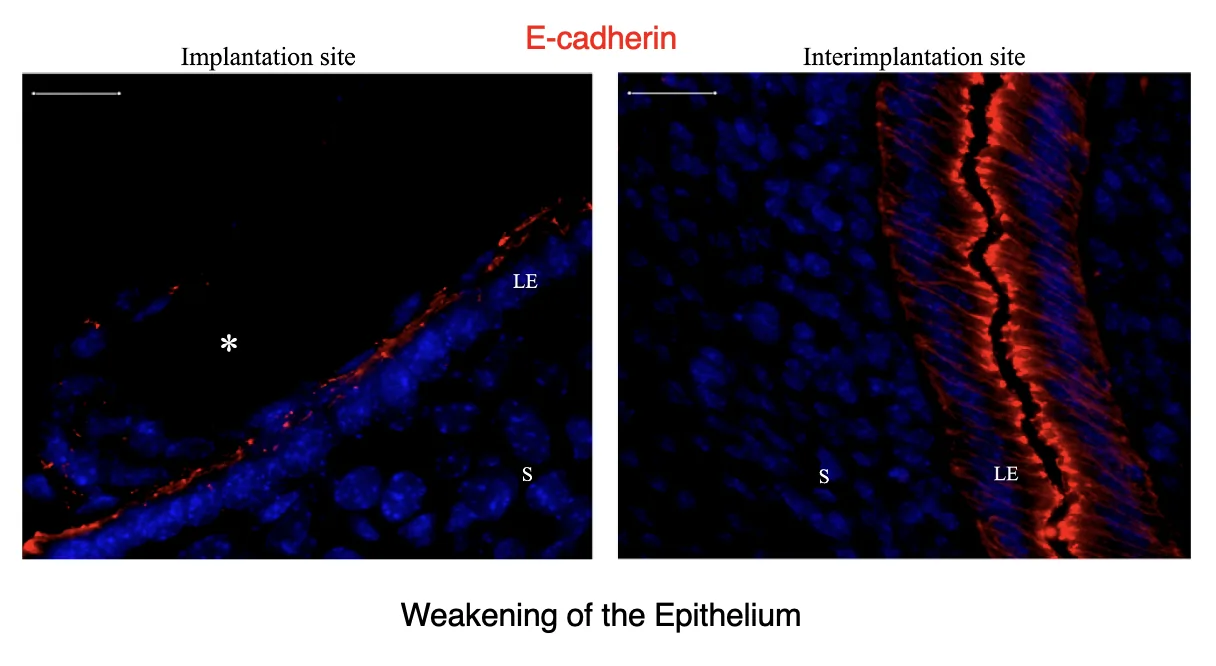
באזור שבו הבלסטוציסט נצמד, האפיתל של הרחם משתנה: תאי האפיתל נעשים שטוחים יותר, וה־Adherens junctions נחלשים.
זה הוסבר דרך צביעת cadherin (שעושים Adherens junctions): באזור ההשרשה (צד שמאל - implantation site) הצביעה חלשה ודלילה יותר לעומת אזור בין אתרי השרשה (צד ימין בתמונה, interimplantation site - אזור ברחם שאין בו עובר שנצמד), שבו רואים צבעיה אדומה חזקה של E-cadherin לאורך האפיתל הלומינלי.
כלומר, באתר ההשרשה, מחסום הקשרים בין תאי האפיתל נחלש. כך נוצר מעבר שמאפשר לתאי העובר להיכנס לתוך הסטרומה.
בתמונה:
| סימון | משמעות |
|---|---|
| LE | אפיתל לומינלי (Luminal epithelium) |
| S | Stroma |
| כוכבית | העובר / אזור ההשרשה |
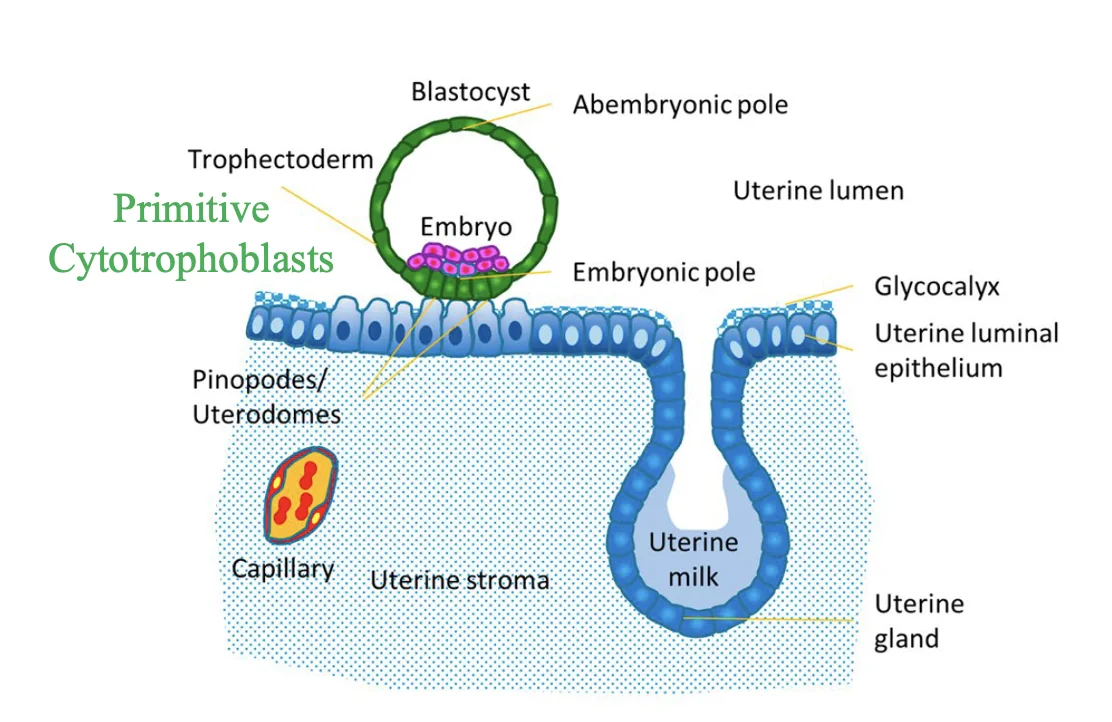
בשלב הפלישה, תאי ה־Trophectoderm בצד הפולרי מתחילים להתחלק ולהתמיין. נוצרים שני סוגים חשובים של טרופובלסטים:
| סוג תא | מאפיין |
|---|---|
| Cytotrophoblasts | תאים טרופובלסטיים דיפלואידיים יותר, עם גרעין יחיד; משמשים גם כמאגר לתאים חדשים |
| Syncytiotrophoblasts | תאים גדולים ורב־גרעיניים שנוצרים מאיחוי של תאים |
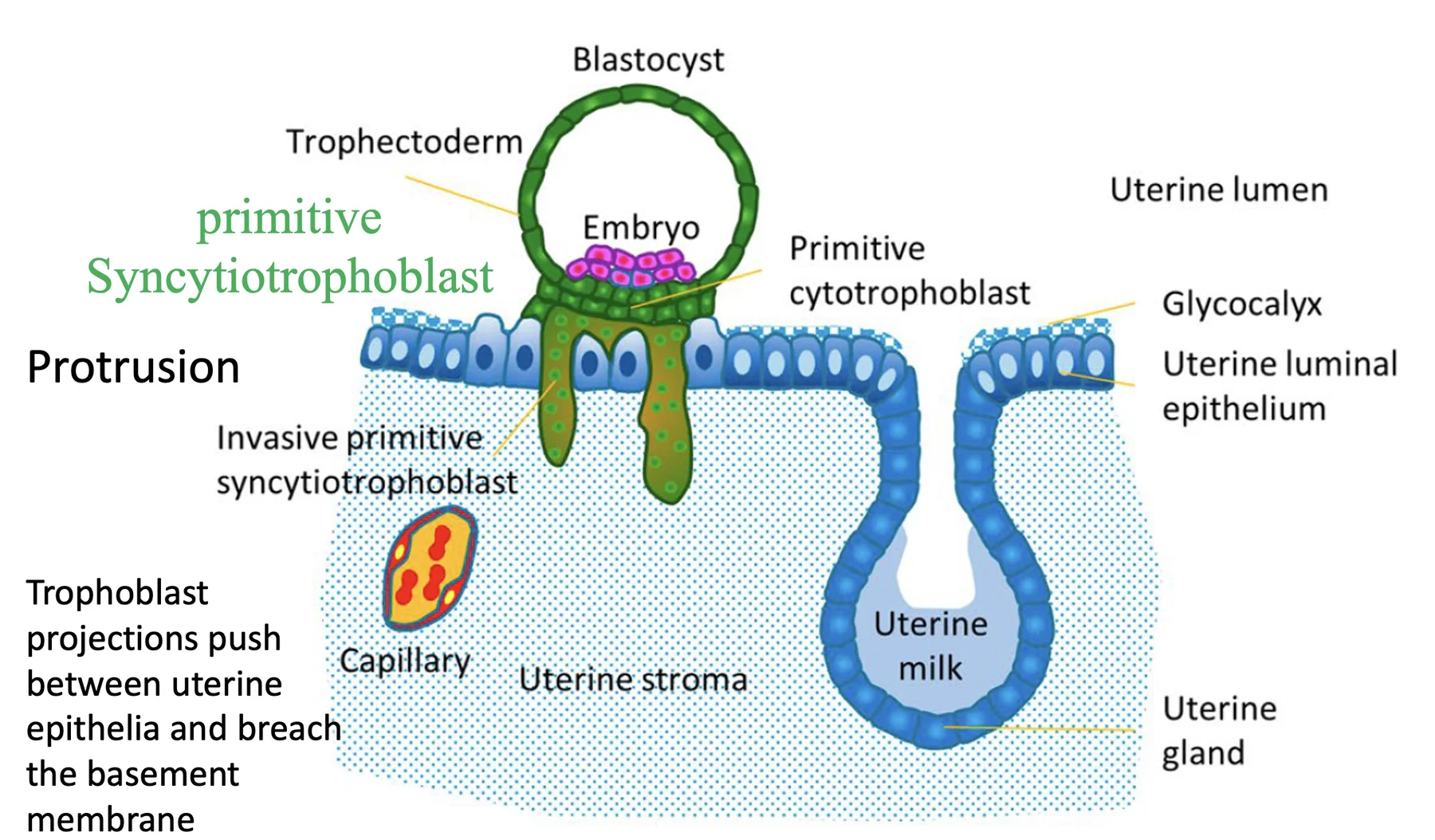
ה־Syncytiotrophoblasts הם התאים הפולשניים יותר. הם שולחים protrusions, נכנסים בין תאי האפיתל, ומגיעים לממברנה הבזלית. דרכם הבלסטוציסט יכול להיכנס בשלמותו לרירית הרחם.
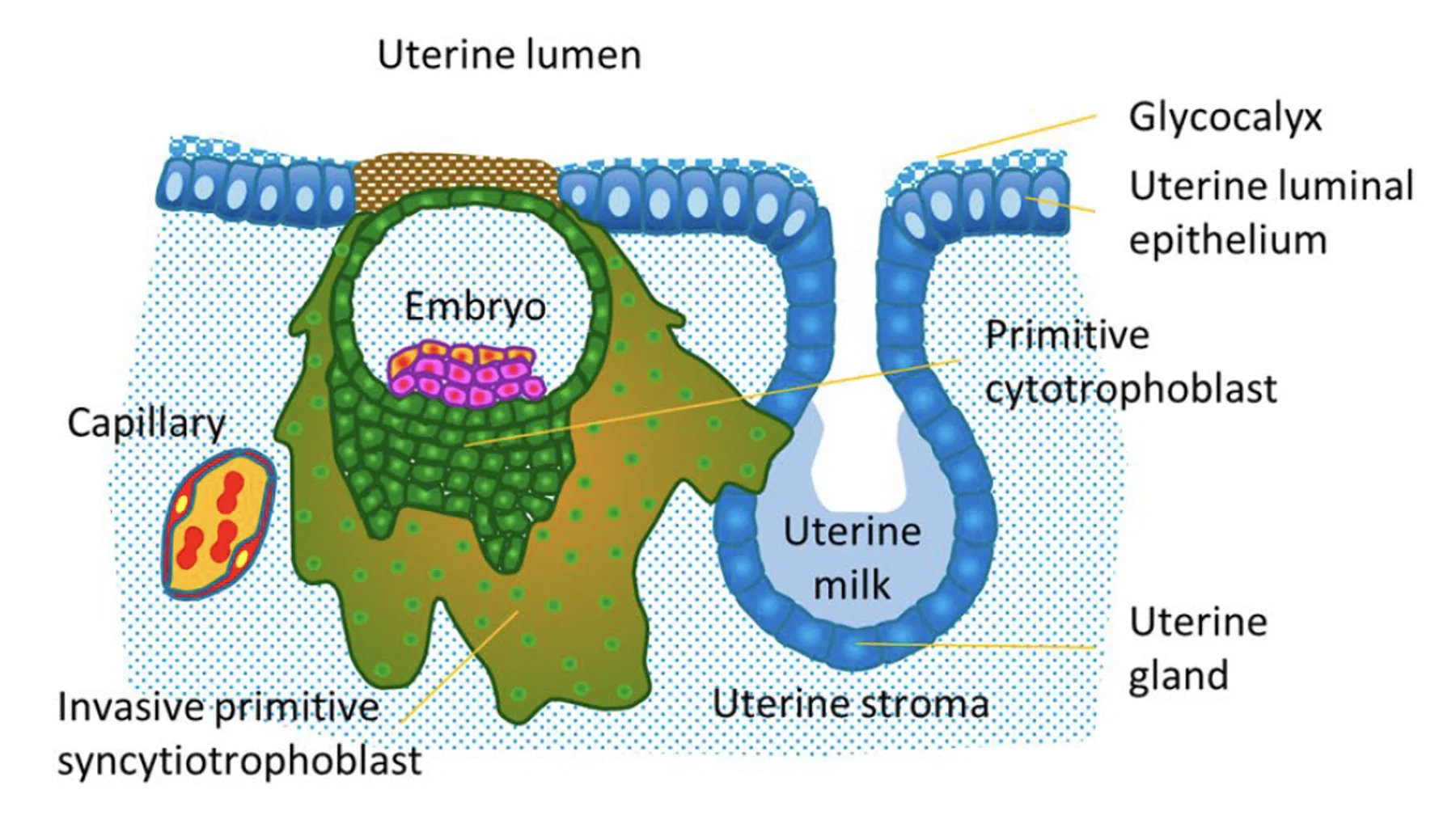
כדי שהעובר ייכנס לסטרומה צריך לפרק שני מחסומים:
- קשרים בין תאי האפיתל, בעיקר Adherens junctions
- הממברנה הבזלית שמתחת לאפיתל
פירוק הממברנה הבזלית ו־Entosis
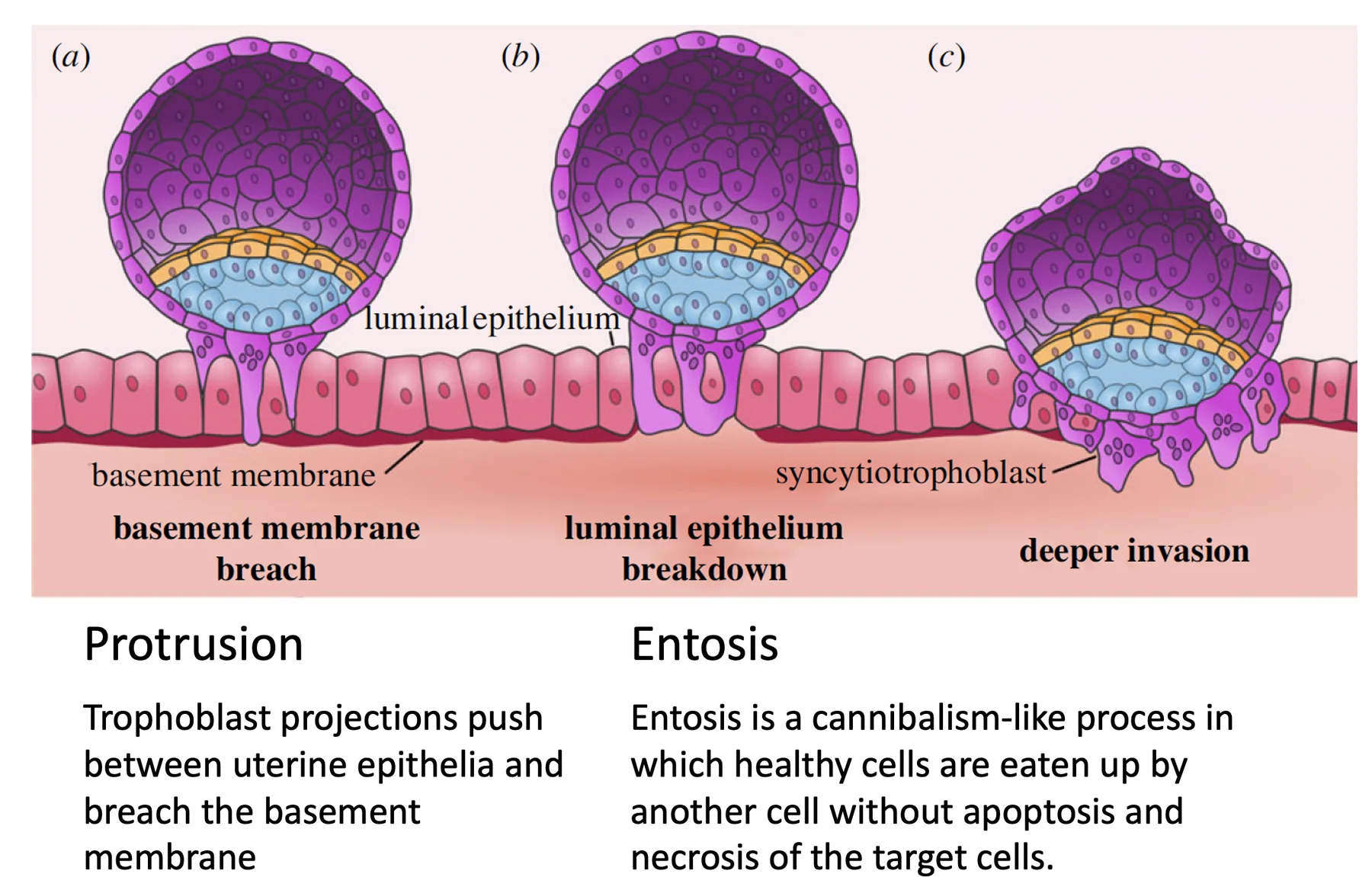
Entosis הוא תהליך שבו תאי הטרופובלסט בולעים תאי אפיתל חיים. זה שונה מפאגוציטוזה רגילה של תא שכבר עבר אפופטוזיס.
כך תאי האפיתל מפונים מהדרך, והעובר יכול להיכנס לסטרומה.
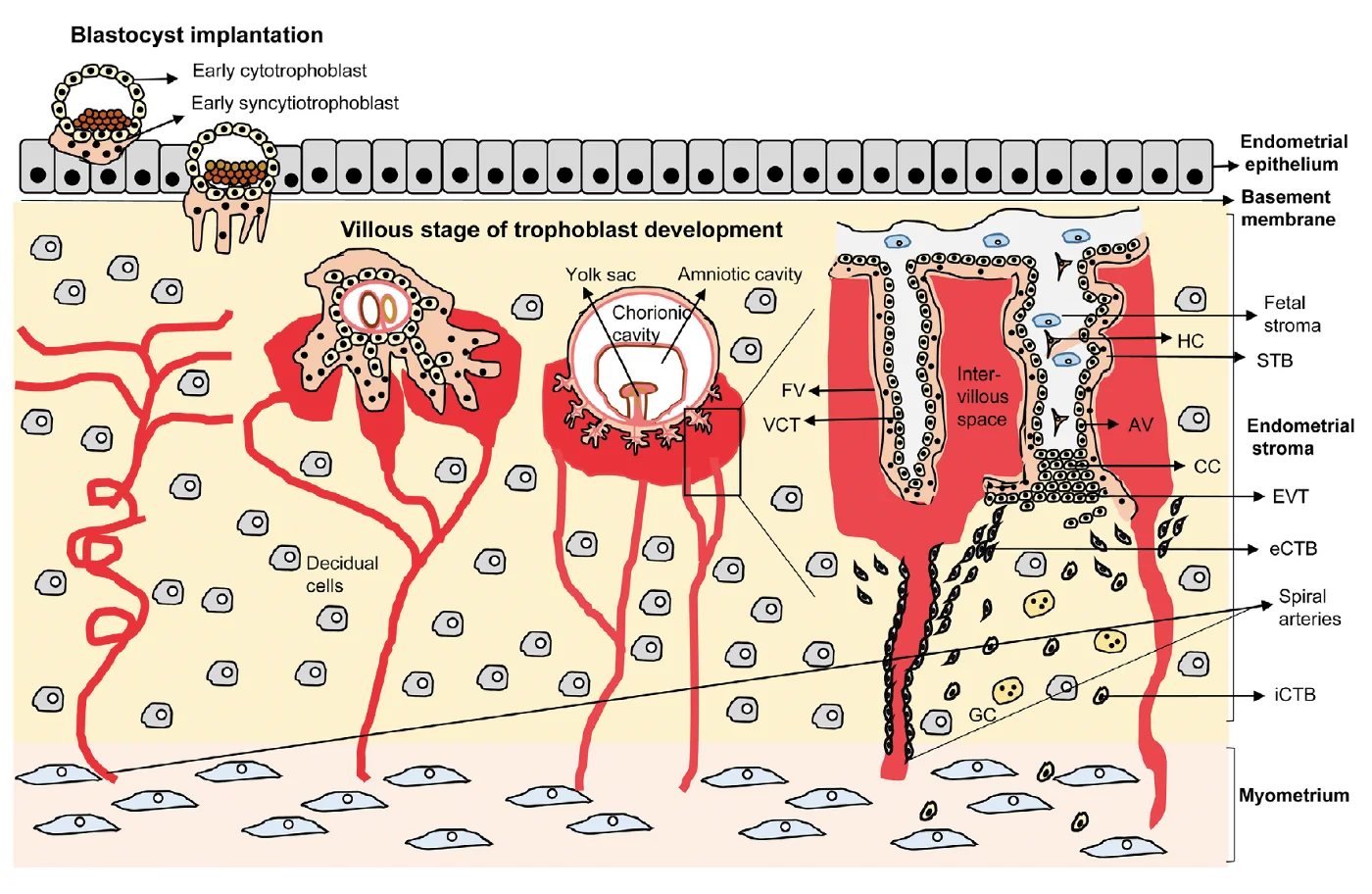
השליה מתפתחת כהמשך ישיר של הטרופובלסטים. יש לה רכיבים עובריים שמקורם בטרופובלסט, ורכיבים אימהיים שמקורם ברקמת הרחם ובסטרומה שעברה decidualization.
השרשה בעכברים
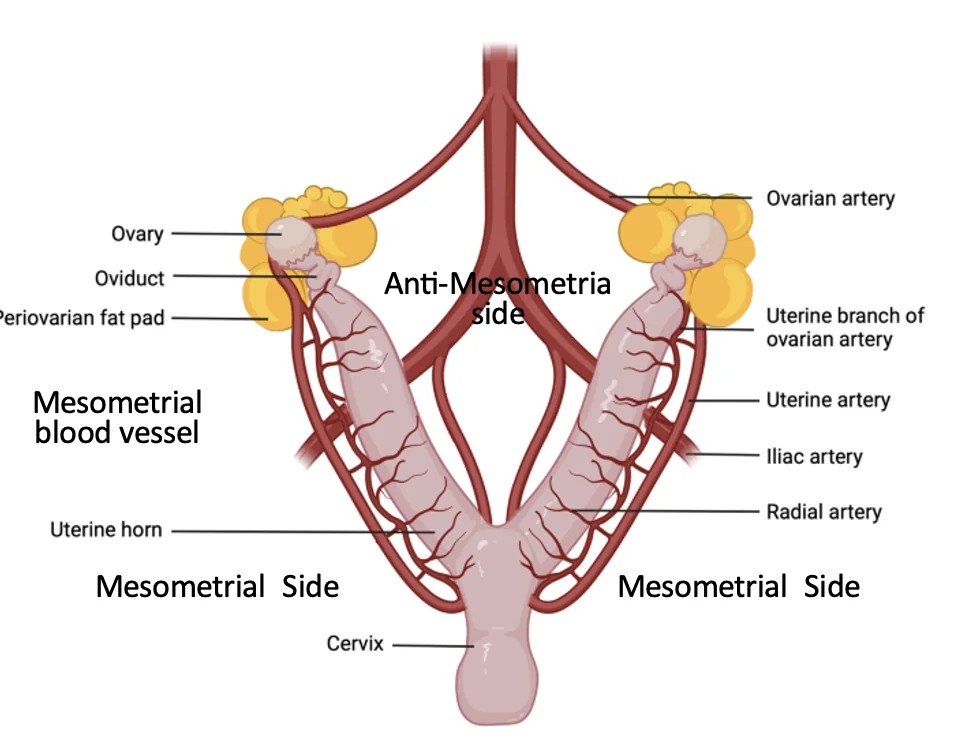
בעכבר, הרחם דו־קרני, וההיריון כולל כמה עוברים לאורך הקרניים. כל עובר עובר השרשה באזור משלו.
האספקה הדמית מגדירה ברחם שני צדדים:
| צד | משמעות |
|---|---|
| Mesometrial side | הצד שבו נכנסים כלי הדם לרחם |
| Anti-mesometrial side | הצד הרחוק מכלי הדם |
העובר מתמקם בצד ה־anti-mesometrial, כלומר רחוק יותר מכלי הדם הגדולים.
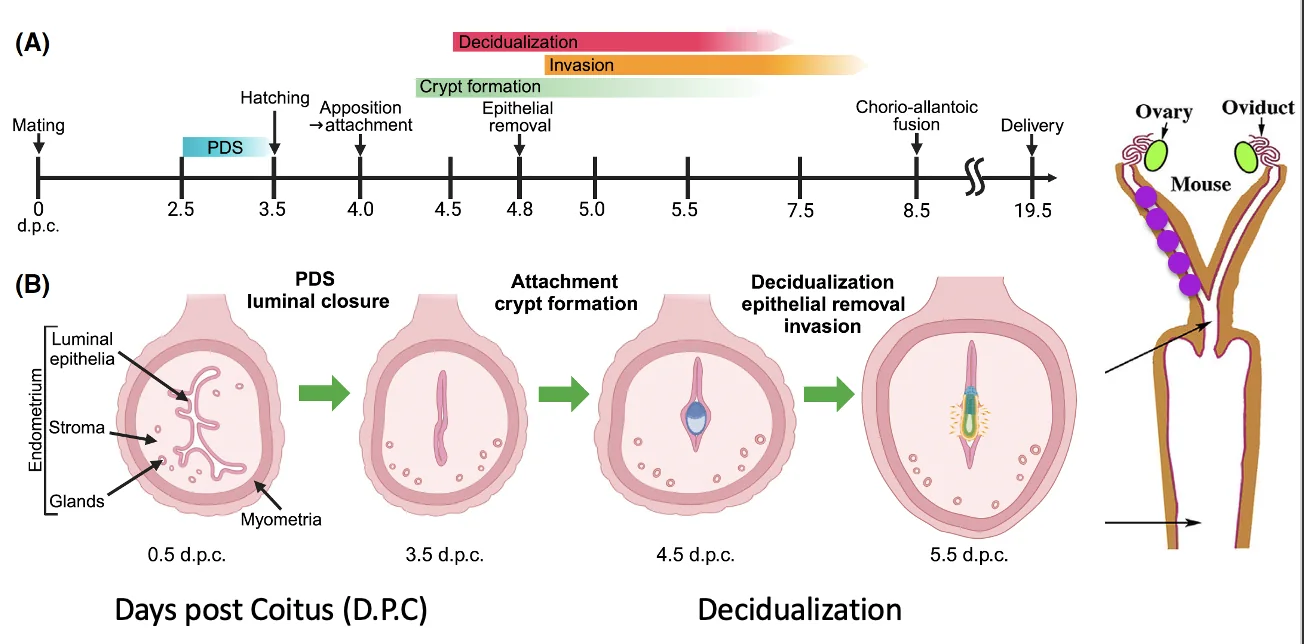
במהלך ההשרשה בעכבר מתרחשת luminal closure: חלל הרחם נסגר סביב העובר ונוצר מעין חלל (chamber) שבו העובר נמצא. משם מתחילה הפלישה לסטרומה.
בעכברים, הצד שעובר הידבקות ראשונית לרירית הוא בעיקר ה־mural trophectoderm. עם זאת, השליה נוצרת בהמשך מהאזור הפולרי של ה־Trophectoderm.
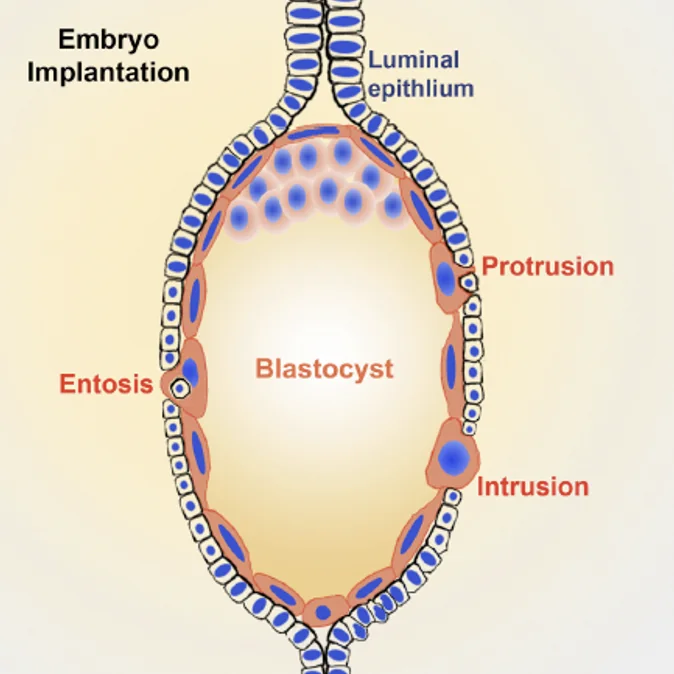
גם בעכבר מתוארים protrusion, intrusion ו־entosis. תאי הטרופובלסט מסייעים בפינוי תאי האפיתל ובהכנסת העובר לסטרומה.
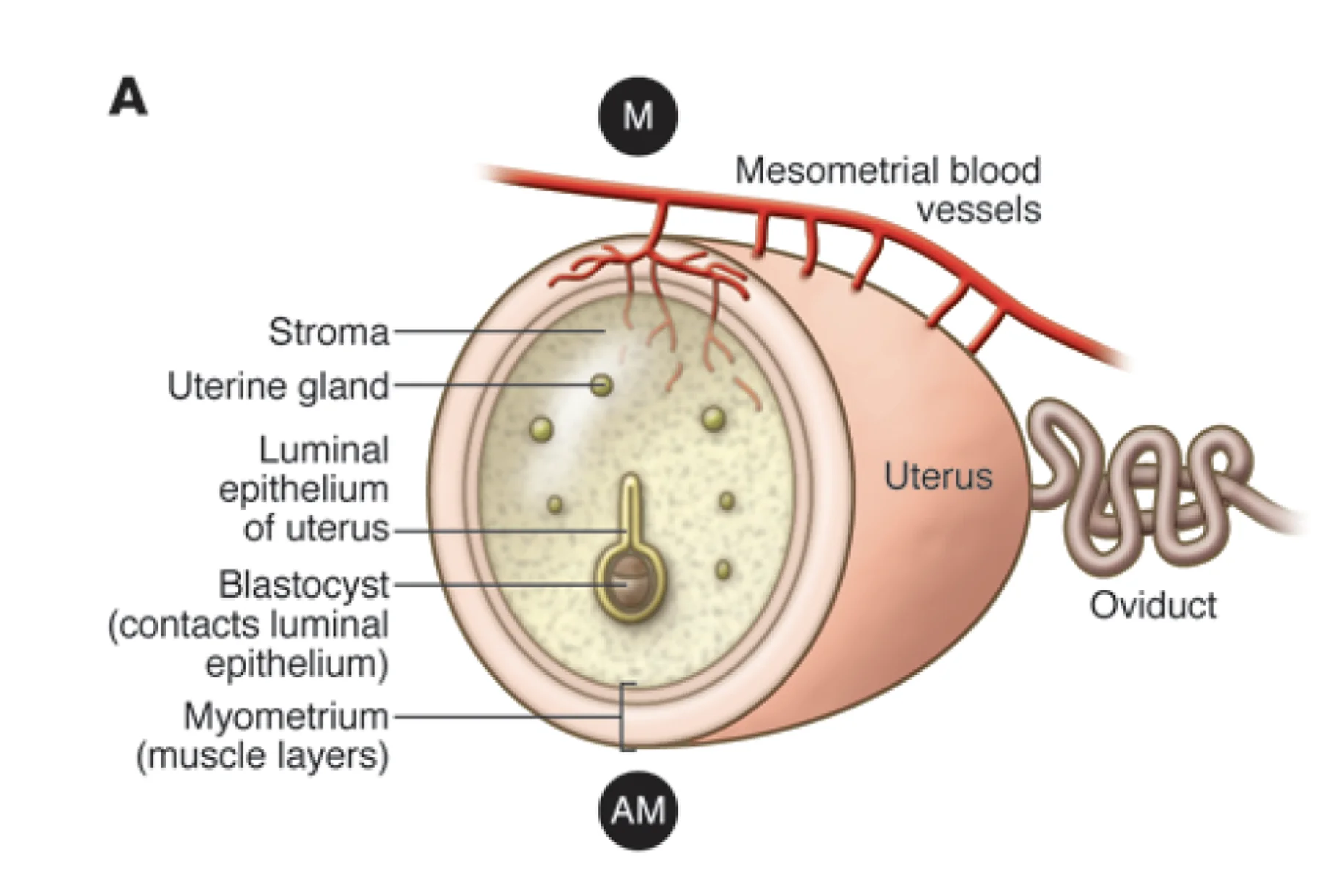
השליה נוצרת באזור ה־polar trophectoderm, גם אם שלב ההידבקות הראשונית בעכבר מתרחש בצד אחר של הבלסטוציסט. כלומר, ההידבקות הראשונית והפלישה קשורות בעיקר ל־mural TE, אבל התפתחות השליה תתרחש מה־polar TE, קרוב יותר לכלי הדם בצד ה־mesometrial.
Decidualization בעכבר
בעכבר, ה־decidualization מופעלת בעקבות adhesion של העובר לרחם, יחד עם הרקע ההורמונלי המתאים. כשמשרים את התהליך באופן ניסויי, רואים גדילה משמעותית של אזור הסטרומה.
הסטרומה סביב העובר מתעבה ומתארגנת מחדש. זה חלק מרכזי מהיכולת של הרחם לקלוט את העובר ולבנות בהמשך את התמיכה הדרושה להתפתחותו.
Pre-gastrulation
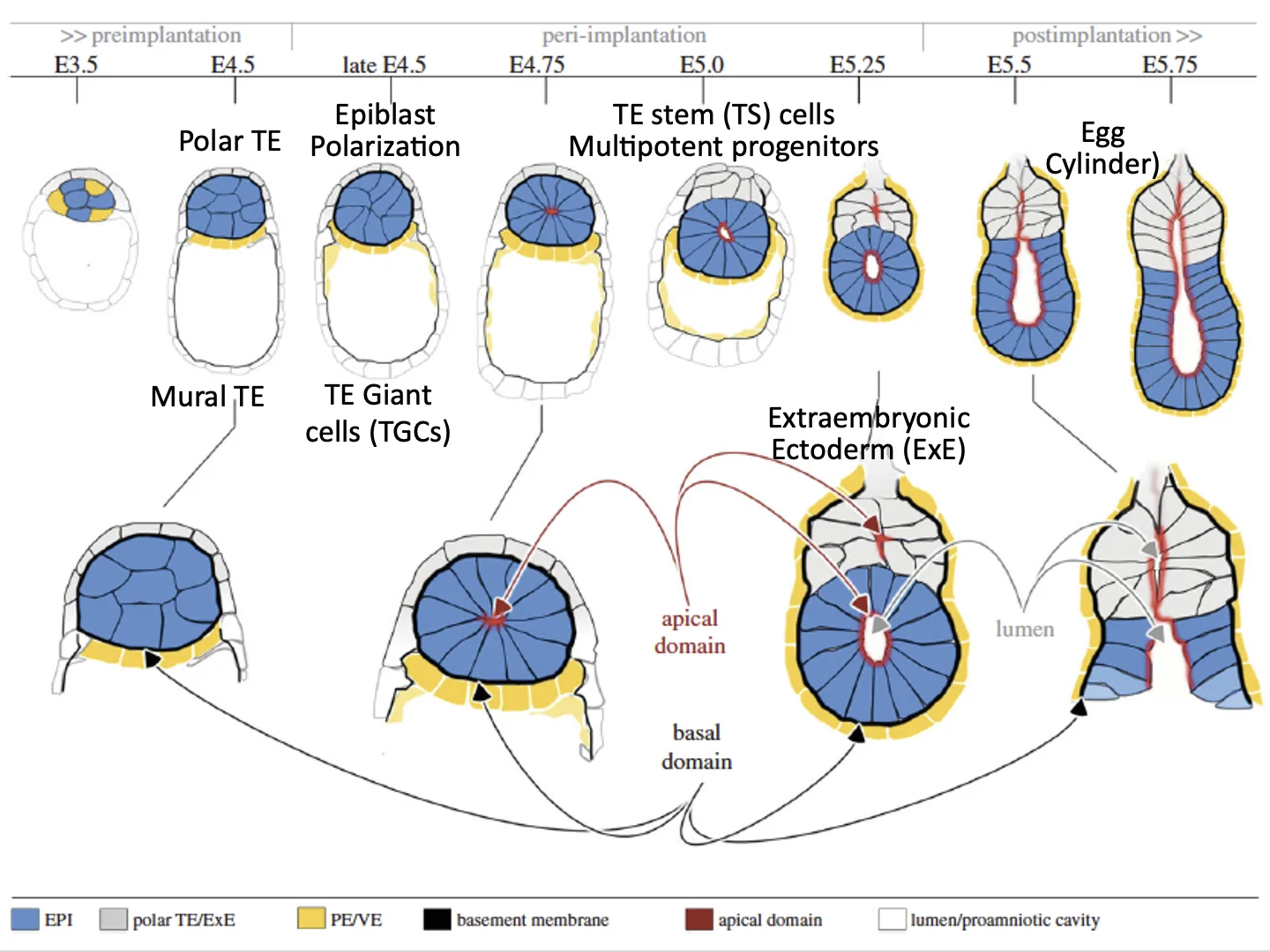
בזמן ההשרשה ואחריה, העובר מתחיל להתארגן לקראת גסטרולציה.
אפשר לחלק את הרצף לשלושה שלבים:
| שלב | מה כולל? |
|---|---|
| Pre-implantation | cleavage, יצירת בלסטוציסט, polar/mural trophectoderm |
| Peri-implantation | ההשרשה עצמה והשינויים הראשונים במבנה העובר |
| Post-implantation | ארגון העובר למבנה שמוכן לגסטרולציה |
הדגש הוא על שני חלקים: peri-implantation, כלומר התהליכים שמתרחשים במהלך ההשרשה, ו־post-implantation, שבו החללים שנוצרו מתחברים והעובר מוכן יותר לגסטרולציה.
יצירת חללים באפיבלסט וב־Extraembryonic Ectoderm
תאי ה־Epiblast עוברים פולריזציה. ברגע שיש להם צד אפיקלי וצד בזלי, מתחיל להיווצר חלל פנימי.
במקביל, תאי ה־polar trophectoderm מתחלקים ויוצרים אוכלוסייה שנקראת extraembryonic ectoderm. גם התאים האלה עוברים פולריזציה ויוצרים חלל.
בהמשך (בשלב ה־Post-implantation), שני החללים מתחברים, ונוצר מבנה מוארך יותר בעובר העכבר: egg cylinder.
סוגי האנדודרם: Primitive, Parietal ו־Visceral
ה־Primitive endoderm מתחיל להתפצל לשתי אוכלוסיות:
| אוכלוסייה | מיקום / תפקיד כללי |
|---|---|
| Parietal endoderm | תאים שנודדים לאורך הצד הפנימי של הטרופובלסטים |
| Visceral endoderm | תאים שנשארים סביב האפיבלסט וה־extraembryonic ectoderm |
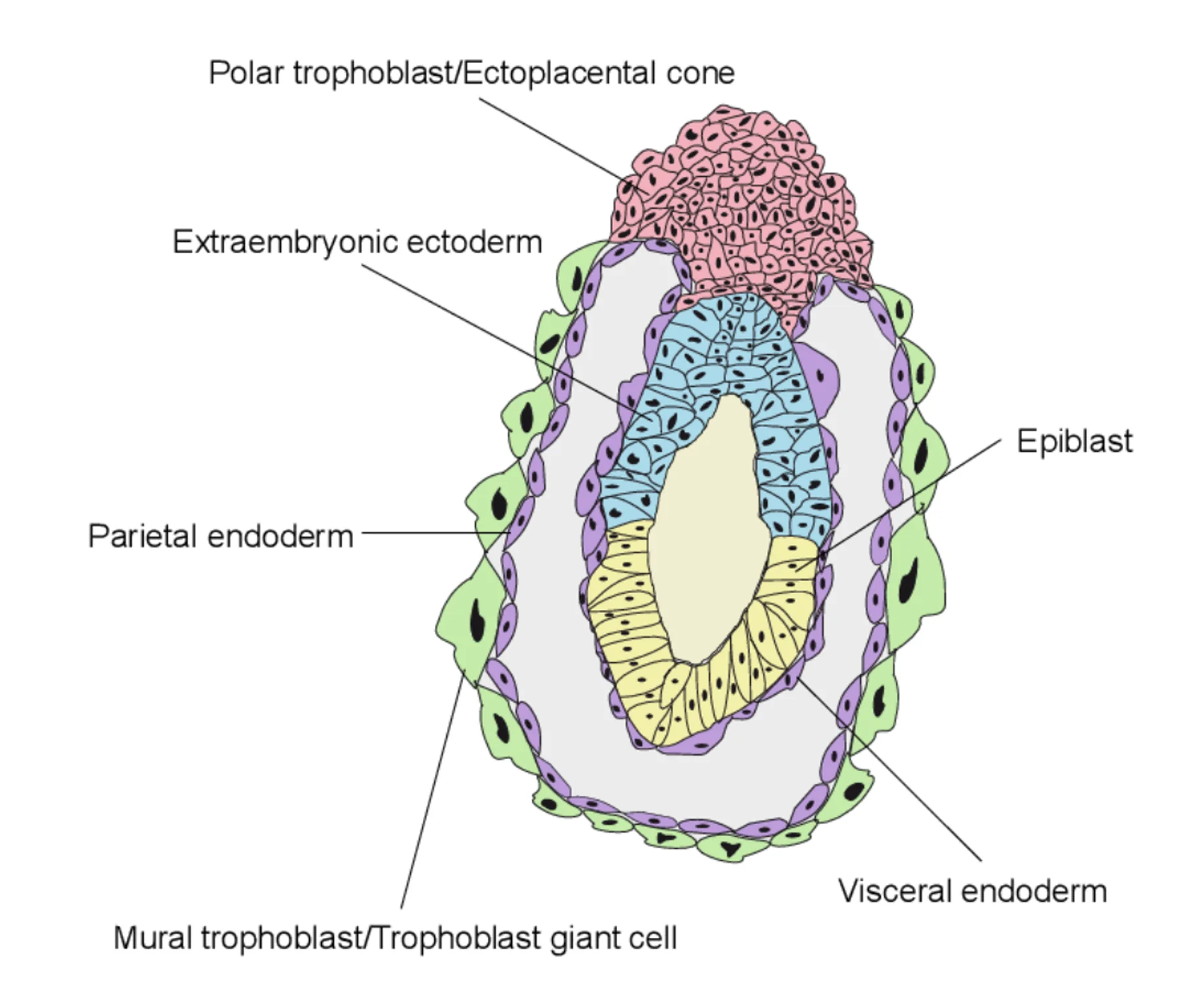
בנוסף נוצר ה־ectoplacental cone, שמקורו גם הוא ב־polar trophectoderm, והוא חלק מהמשך התפתחות השליה.
עכבר לעומת אדם לאחר השרשה
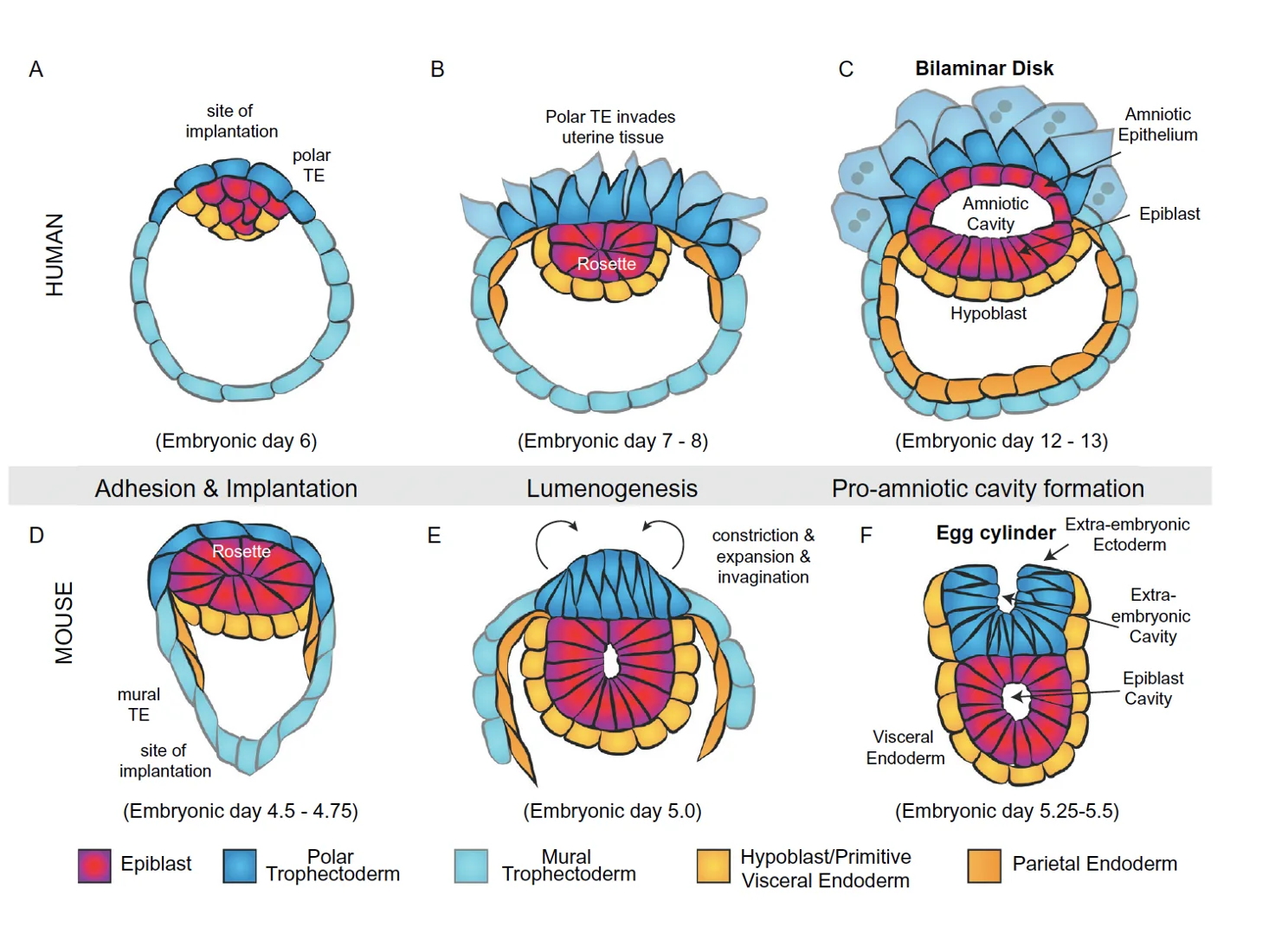
העקרונות המולקולריים דומים, אבל המבנה האנטומי שונה.
בעכבר, האפיבלסט וה־extraembryonic ectoderm יוצרים מבנה דמוי כוס או גליל - egg cylinder.
באדם, האפיבלסט יוצר מבנה שטוח יותר, דיסק של תאים. בשלב הזה נוצר גם amniotic cavity, וה־amnion שמקורו בתאי האפיבלסט. מתחת לאפיבלסט נמצא ה־hypoblast, המקביל בהקשר הזה ל־visceral endoderm.
ההבדל הוא בעיקר במבנה המרחבי. התהליכים המרכזיים - פולריזציה, יצירת חללים, ארגון אפיבלסט ורקמות חוץ־עובריות - נשארים דומים ברמת העיקרון.
צירי הגוף
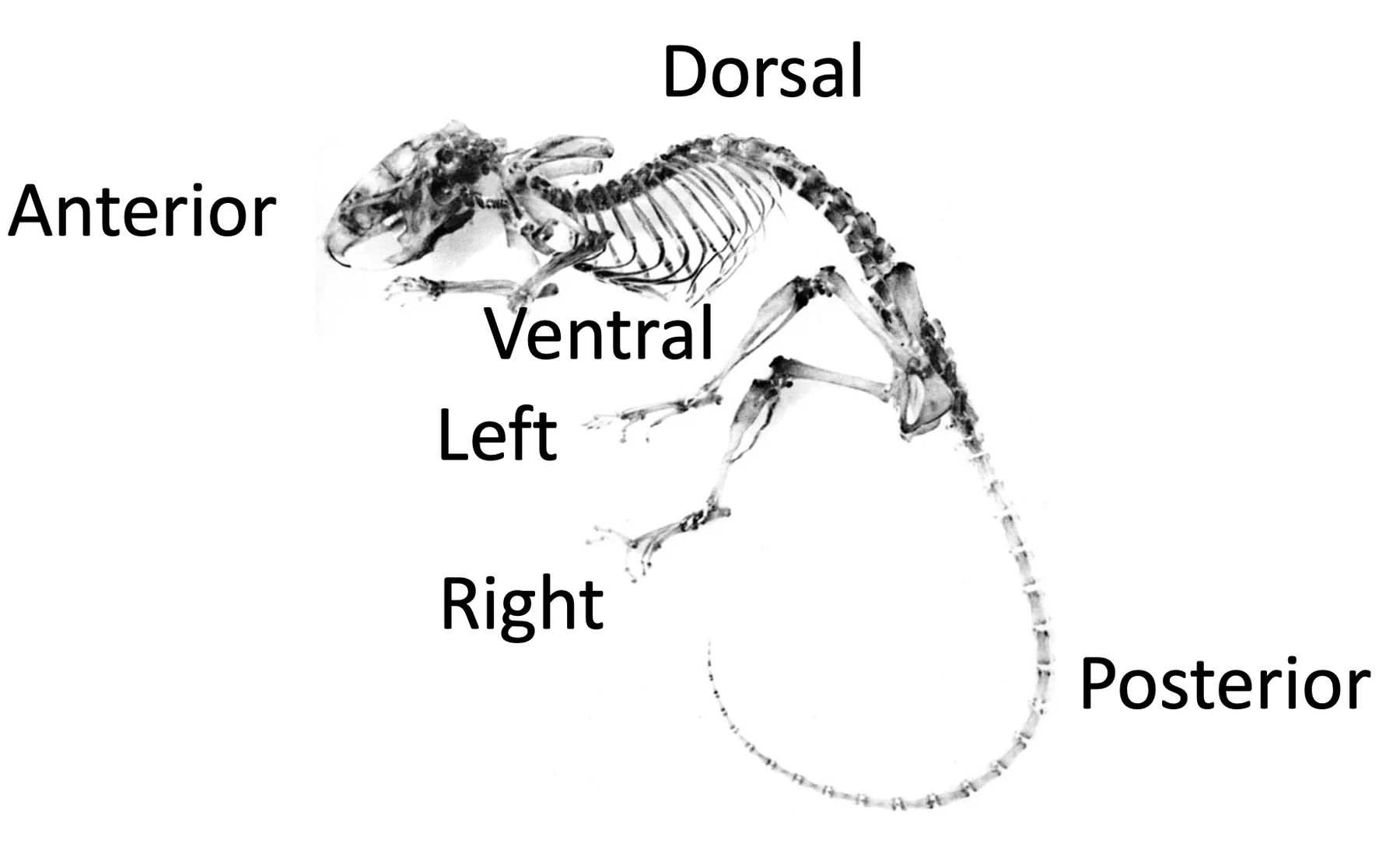
לקראת גסטרולציה מתחילים להשתמש בשלושה צירי גוף מרכזיים:
| ציר | משמעות |
|---|---|
| Anterior-Posterior | ראש־זנב |
| Left-Right | שמאל־ימין |
| Dorsal-Ventral | גב־בטן |
המשך ההתפתחות, כולל גסטרולציה ואורגנוגנזה, יעסוק בשאלה איך הצירים הללו נוצרים ואיך הם מכוונים את התפתחות הגוף.
רצף האירועים המרכזי
Oocyte
↓
Maternal RNA and proteins stored in the egg
↓
SCMC located under the membrane near cortical actin
↓
Fertilization
↓
Pronuclear fusion and chromosome positioning near the center
↓
First cleavage along animal-vegetal axis
↓
Two blastomeres of similar size
↓
Balanced distribution of essential maternal transcripts
↓
ZGA and early probabilistic differences between blastomeres
↓
Cleavage → morula → blastocyst
↓
Hatching from zona pellucida
↓
Implantation window
↓
Apposition → adhesion → invasion
↓
Decidualization and trophoblast differentiation
↓
Pre-gastrulation organization
↓
Epiblast, extraembryonic ectoderm, visceral/parietal endoderm
↓
Embryo prepared for gastrulation
משפטי מפתח
- SCMC הוא קומפלקס אימהי שנמצא מתחת לממברנה בביצית ובשלבי cleavage מוקדמים.
- FLOPED, TLE6, MATER ו־FILIA הם חלק מהקומפלקס שהוצג בשיעור.
- TLE6 שומר על ארגון הקומפלקס; ב־Tle6 null החלבונים קיימים אך מפוזרים, וההתפתחות נעצרת מוקדם בדומה ל־Floped null.
- פגיעה ב־MATER נותנת פנוטיפ דומה, ולכן הבעיה היא בתפקוד ה־SCMC כקומפלקס ולא רק בחלבון יחיד.
- תפקיד מרכזי של SCMC הוא למקם את הכרומוזומים בצורה שמאפשרת חלוקה ראשונה לשני תאים דומים בגודלם.
- Maternal effect genes פועלים דרך RNA וחלבונים שהאם הכניסה לביצית עוד לפני שהגנום העוברי פעיל.
- פיזור לא הומוגני של RNA בביצית יכול ליצור הטיות מוקדמות בין בלסטומרים.
- המודל שהוצג מסביר שבירת סימטריה כהטיה פרובביליסטית, לא כקביעה מוחלטת של גורל התא.
- בעכבר ZGA מתרחש סביב שני תאים; באדם סביב ארבעה תאים.
- IVF סטנדרטי מפגיש ביצית ותאי זרע בצלחת; ICSI מחדיר תא זרע יחיד לתוך הביצית.
- PGD מחפש מוטציה ידועה; PGS מחפש בעיקר הפרעות כרומוזומליות רחבות, במיוחד בהקשר של הפלות חוזרות או אובדני היריון.
- ב־PGD אפשר לדגום בלסטומר לפני compaction, תאי TE מהבלסטוציסט, או גופיפים פולריים מהביצית/זיגוטה.
- השרשה דורשת בלסטוציסט קומפטנטי ורחם רצפטיבי באותו חלון זמן.
- Decidualization מכינה את הסטרומה לקליטת העובר ולתמיכה בשליה; בעכברים יכולה להופיע גם polyploidization של תאי הסטרומה.
- באדם decidualization מתרחשת גם בלי עובר; בעכבר היא תלויה בהידבקות של העובר.
- בהשרשה, הטרופובלסטים פולשים דרך אפיתל הרחם אל הסטרומה.
- Syncytiotrophoblasts נוצרים מאיחוי תאי טרופובלסט, והם משתתפים בפלישה לרירית, בפירוק הממברנה הבזלית וב־entosis של תאי אפיתל חיים.
- לפני גסטרולציה, האפיבלסט וה־extraembryonic ectoderm עוברים פולריזציה ויוצרים חללים.
- בעכבר מתקבל egg cylinder; באדם מתקבל דיסק אפיבלסטי עם amniotic cavity.
- הצירים המרכזיים להמשך ההתפתחות הם anterior-posterior, left-right ו־dorsal-ventral.