חזרה קצרה - שברי DNA ומנגנוני תיקון
השיעור נפתח בחזרה על הנושאים של שיעור 13: תיקון שברים דו־גדיליים, ATM, BRCA, PARP inhibitors ושיטות לזיהוי שברי DNA.
הנקודות החשובות:
- פגם בחלבוני Ku יפגע בעיקר בתיקון שברים דו־גדיליים בשלב G1, כי בשלב הזה אין כרומטידה אחות זמינה ולכן התא תלוי ב־NHEJ.
- PARP inhibitors גורמים לרמות גבוהות של נזקי DNA, אבל הם לא מעלים ישירות את פעילות Homologous Recombination. הקשר ל־HR הוא שהנזקים שהם יוצרים צריכים להיות מתוקנים על ידי HR.
- טיפול ב־PARP inhibitors יעיל במיוחד בתאים עם פגם ב־BRCA1/2 בגלל synthetic lethality: שילוב של יצירת נזקים וחוסר יכולת לתקן אותם.
- פגם ב־ATM גורם לרגישות לקרינה מייננת כי התא לא מאותת נכון על שברים דו־גדיליים.
- פגם ב־Ligase IV יכול להסביר חוסר בתאי B ו־T, כי Ligase IV חיוני ל־NHEJ, ו־NHEJ חיוני ליצירת מערכת החיסון הנרכשת.
- תנאים שמעודדים HR על פני NHEJ הם שלב S/G2, פעילות BRCA1, ו־end resection.
- Comet assay מבוסס על ג׳ל אלקטרופורזה של DNA צבוע פלואורסצנטית, ולא על נוגדן.
- ב־END-seq שומרים את ה־DNA בתוך אגרוז כדי למנוע שברים מכניים בזמן הכנת הספרייה.
- התבנית ב־HR מגיעה מהכרומטידה האחות, לא מהכרומוזום ההומולוגי.
- ATM לא מחבר את קצוות ה־DNA בעצמו; הוא מתאם את תגובת הנזק, כולל פוספורילציה של H2AX, בקרת מחזור התא והכוונת בחירת מסלול התיקון.
- 53BP1 מקדם חסימת resection ולכן דוחף לכיוון NHEJ.
- BRCA-ness מתאר מצב של חסר ב־HR בלי מוטציה ישירה ב־BRCA.
מכאן ממשיכים לנושא של השיעור: נזקי DNA שמתרחשים בכוונה כחלק מתהליכים פיזיולוגיים.
נזקי DNA יזומים במערכת החיסון
בשיעורים הקודמים שברי DNA תוארו בעיקר כבעיה: נזק שיכול לגרום למוות תאי, אי־יציבות גנומית או סרטן. כאן הנקודה משתנה: יש תהליכים שבהם התא יוצר שברי DNA באופן יזום כדי לבצע פעולה חיונית.
דוגמאות לנזקי DNA יזומים:
- Meiotic recombination - שחלוף בין כרומוזומים הומולוגיים לפני מיוזה, בין השאר בעזרת SPO11.
- Topological stress relaxation - שברים זמניים שנוצרים על ידי topoisomerases כדי לשחרר מתח טופולוגי.
- V(D)J recombination - יצירת רפרטואר ראשוני של נוגדנים ו־T-cell receptors.
- Somatic hypermutation - יצירת מוטציות נקודתיות בנוגדנים כדי לשפר אפיניות לאנטיגן.
בשיעור הזה הדגש הוא על מערכת החיסון הנרכשת. לא צריך להיכנס לאימונולוגיה עצמה מעבר למה שנדרש כדי להבין את מנגנוני ה־DNA.
שני שלבים ביצירת רפרטואר חיסוני
בתאי B יש שני שלבים עיקריים הקשורים לגיוון הנוגדנים:
- יצירה ראשונית של רפרטואר מגוון - לפני חשיפה לאנטיגן. זה נעשה על ידי V(D)J recombination.
- שיפור האפיניות לאחר חשיפה לאנטיגן - אחרי שכבר יש נוגדנים שמזהים אנטיגן מסוים. זה נעשה על ידי somatic hypermutation ו־affinity maturation.
בתהליך הראשון נוצרים רצפטורים רבים באופן אקראי. בתהליך השני נוצרת סלקציה: תאים עם נוגדנים שקושרים טוב יותר את האנטיגן שורדים ומתרבים יותר.
V(D)J recombination ו־RAG
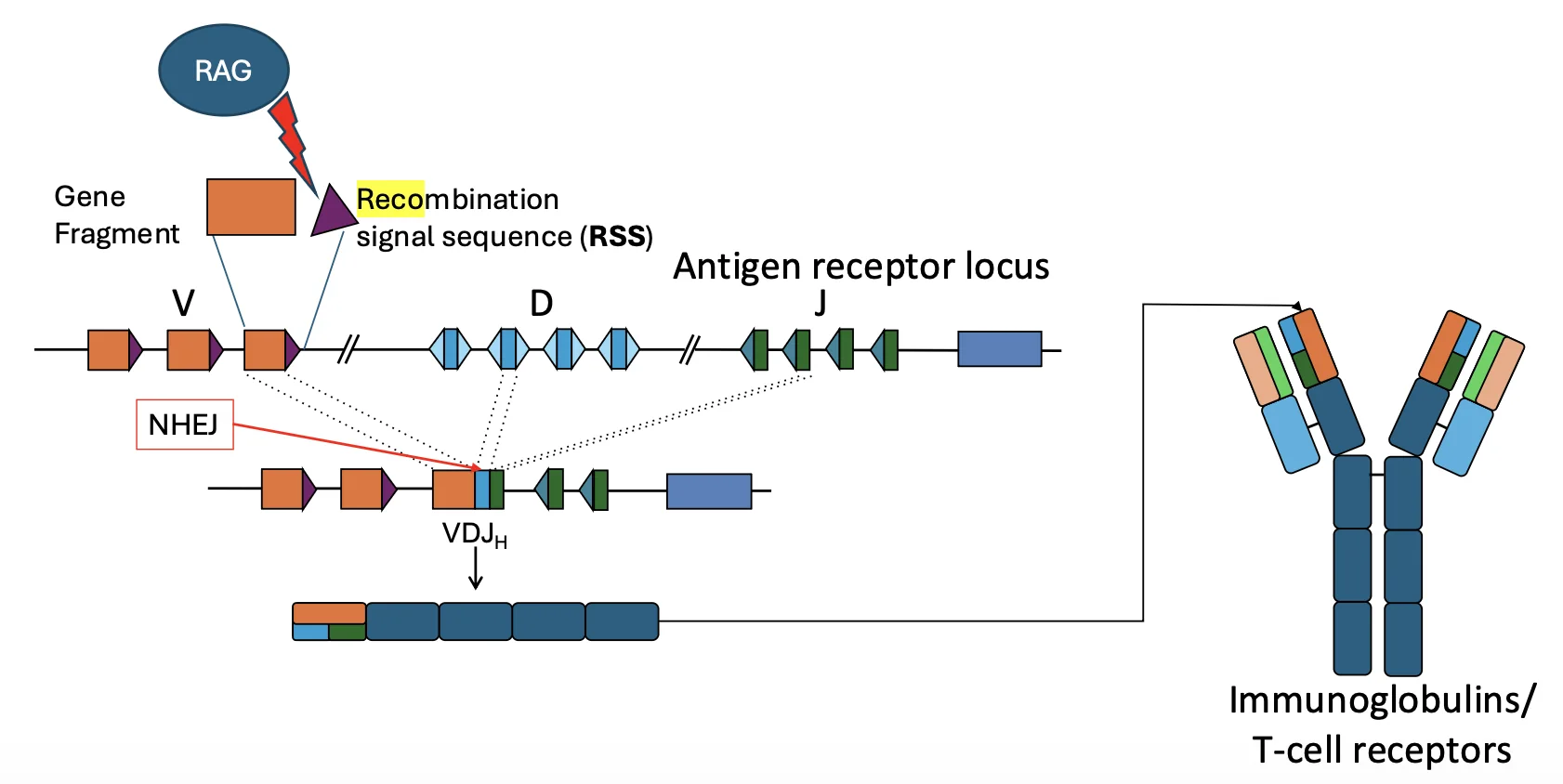
בדרך כלל גן אחד מכיל רצף אחד שמקודד לחלבון אחד. בנוגדנים וב־T-cell receptors צריך משהו אחר: אזור גנטי אחד צריך לייצר המון חלבונים שונים.
זה מתאפשר בזכות המבנה של Antigen receptor locus.
בלוקוס הזה יש מקטעים מסוגים שונים:
- V
- D
- J
בכל תא נבחרים מקטעים מסוימים מתוך המאגר, והם מחוברים יחד. השילוב שנוצר מקודד לאזור המשתנה של הנוגדן או הרצפטור. האזור המשתנה הוא זה שקובע את הספציפיות לאנטיגן.
מדובר בעריכה של ה־DNA עצמו, לא של RNA. לכן תאי B או T שעברו רקומבינציה שונים זה מזה ממש ברמת הגנום.
RAG מבצע את החיתוך
האנזים שמתחיל את התהליך הוא RAG - Recombination Activating Gene.
RAG מזהה רצפים שנקראים:
RSS - Recombination Signal Sequences
הרצפים האלה נמצאים ליד מקטעי V, D ו־J. RAG חותך סמוך אליהם, וכך יוצר שברים דו־גדיליים מבוקרים.
אחרי החיתוך צריך לחבר מחדש את הקצוות. החיבור נעשה על ידי NHEJ.
לכן V(D)J recombination הוא למעשה תהליך של:
cut by RAG → repair/joining by NHEJ
זו גם הסיבה שפגם ב־NHEJ יכול לגרום ל־immunodeficiency: החיתוך יכול להתרחש, אבל החיבור מחדש נכשל.
איך שולטים ב־cut and paste?
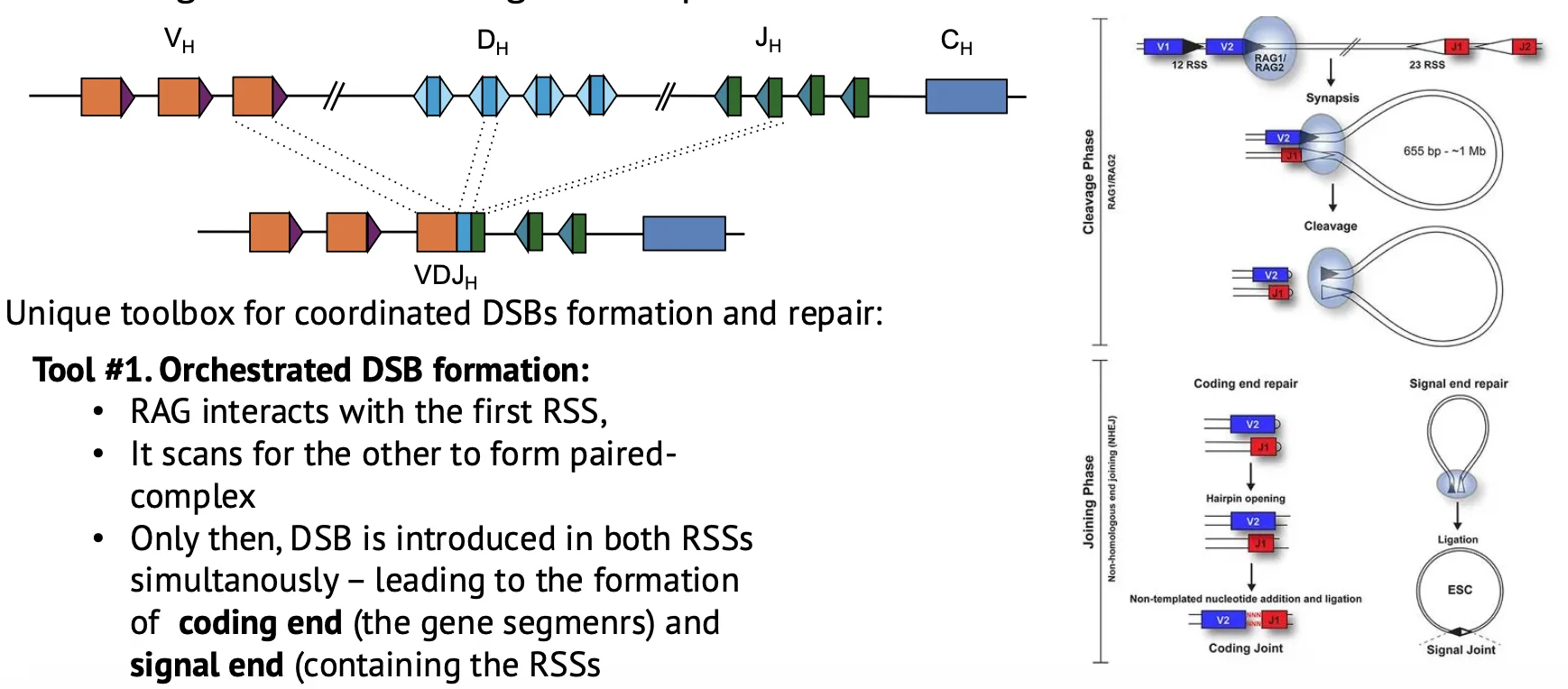
האתגר הגדול הוא לשלוט באנזים שיוצר שברי DNA. RAG חייב לחתוך רק באזורים הנכונים, בזמנים הנכונים, ולדאוג שהקצוות יתחברו נכון.
יש כמה שכבות בקרה.
חיתוך רק כאשר שני הצדדים נמצאים יחד
RAG לא אמור לחתוך כל RSS בנפרד. הוא יוצר paired complex: שני הרצפים שאמורים לעבור רקומבינציה מובאים יחד, ורק אז נוצרים השברים.
כך קטן הסיכון שקצה DNA אחד יישאר חופשי ויתחבר בטעות לשבר אחר בגנום.
לאחר החיתוך נוצרים שני סוגי קצוות:
- Coding ends - הקצוות שיחוברו ויהפכו לחלק מהרצף המקודד לנוגדן.
- Signal ends - הקצוות של המקטע שנחתך החוצה, לרוב כדנ״א מעגלי.
Hairpin מגן על הקצוות
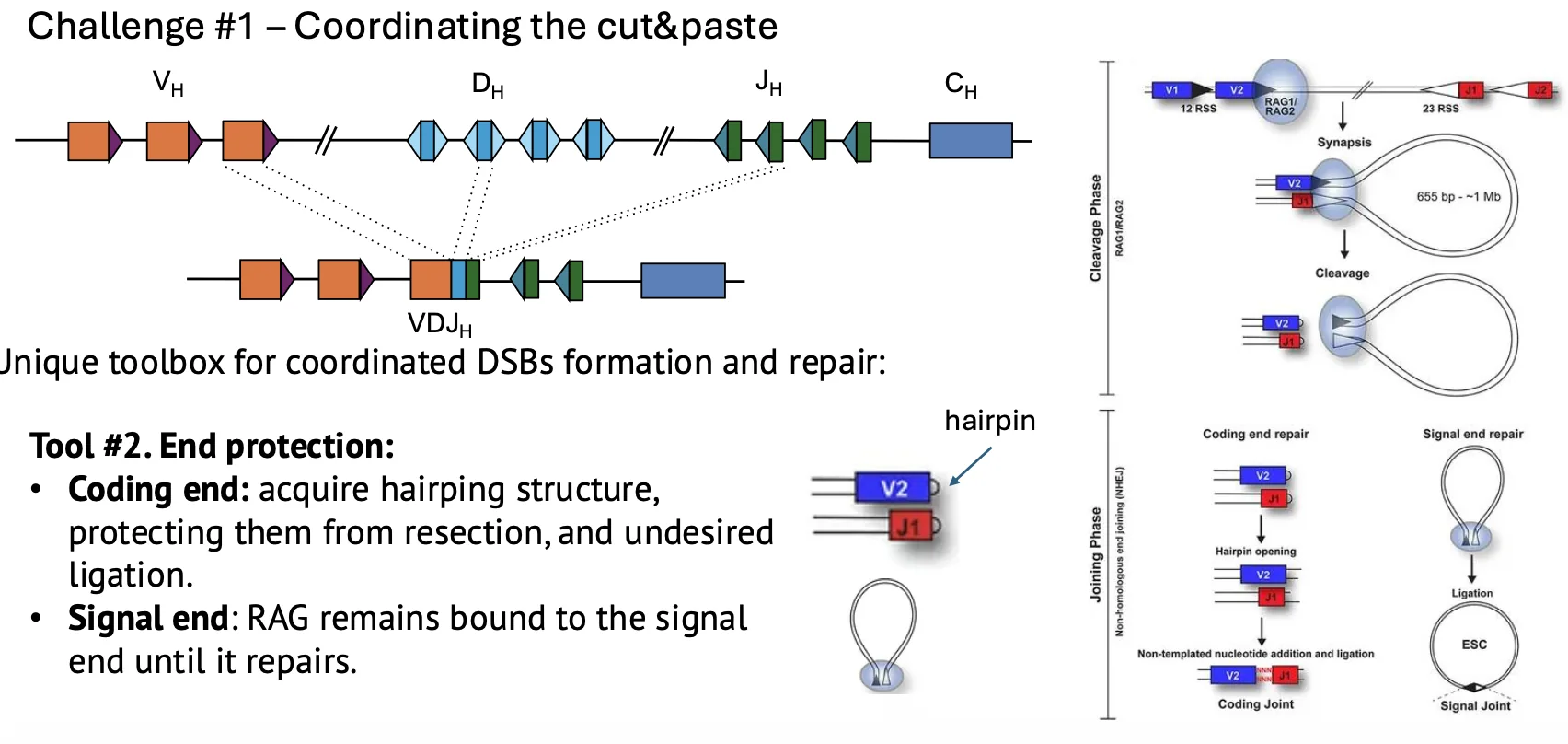
קצה DNA חופשי הוא מבנה מסוכן, כי הוא יכול להתחבר לקצה לא נכון וליצור טרנסלוקציה.
לכן ב־coding ends נוצר מבנה hairpin. זה סוג של חסימה של הקצה, שמקטינה את הריאקטיביות שלו עד שמנגנון התיקון המתאים יגיע.
במקביל, RAG נשאר קשור ל־signal ends ומגן גם עליהם.
גיוס מהיר של NHEJ
RAG לא רק חותך. הוא גם עוזר לגייס את חלבוני NHEJ, ובפרט את חלבוני Ku, שהם הראשונים שמזהים קצוות DNA בתיקון NHEJ.
המטרה היא שהחיבור יקרה מהר, לפני שהקצוות יספיקו להשתתף בתיקון שגוי.
כלל 12/23
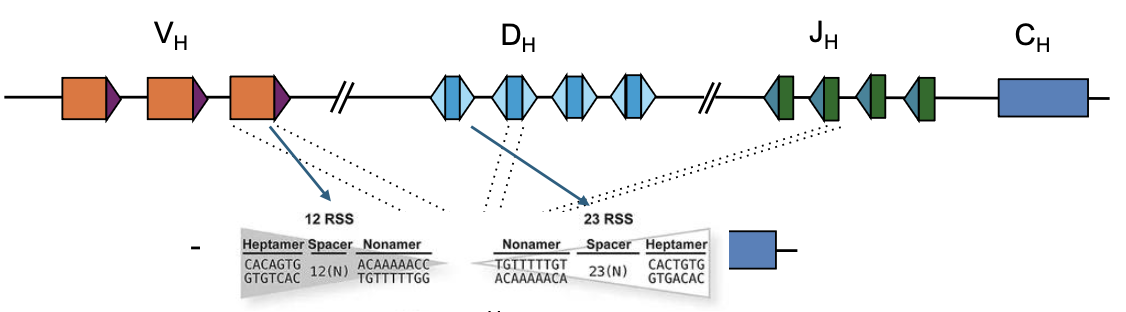
יש שני סוגים של RSS:
- 12 RSS
- 23 RSS
רקומבינציה תקינה מתרחשת בין RSS מסוגים שונים: 12 עם 23. הכלל הזה מצמצם חיבורים לא רצויים, למשל חיבור בין שני מקטעים מאותו סוג.
RAG כטרנספוזאז שעבר ביות
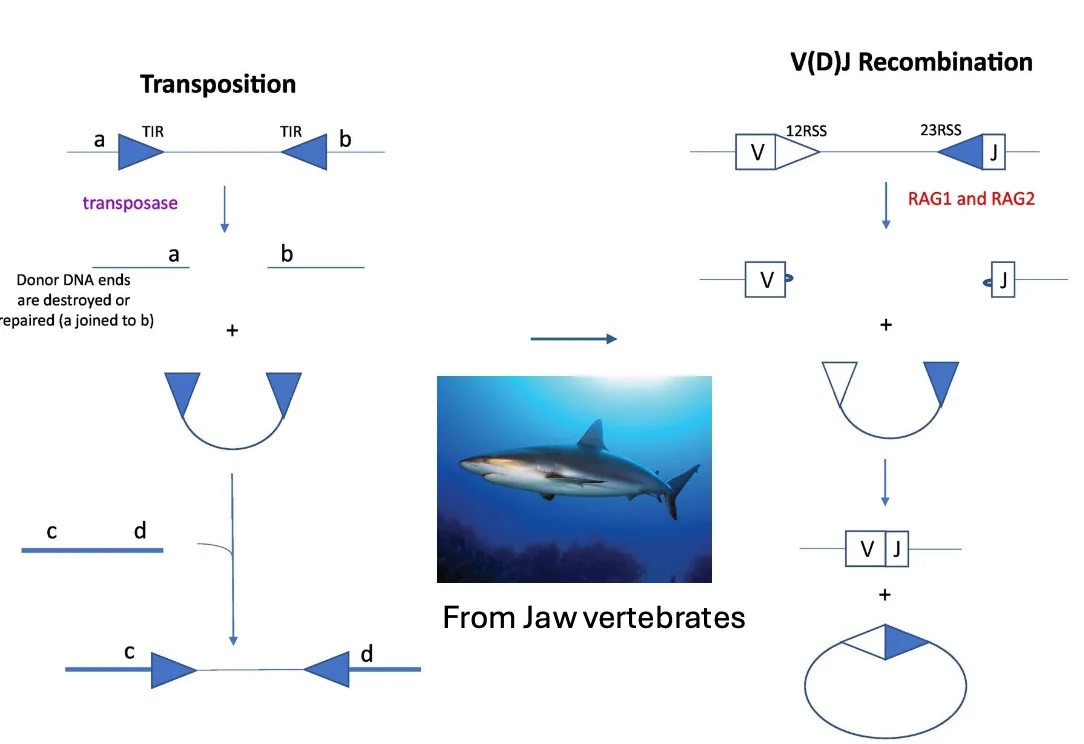
RAG התפתח מאנזים דמוי transposase.
Transposase הוא אנזים שיודע להזיז מקטעי DNA ממקום למקום בגנום. לכן יש דמיון רעיוני בין טרנספוזיציה לבין V(D)J recombination: בשניהם יש זיהוי רצפים, חיתוך DNA וחיבור מחדש.
אבל RAG עבר domestication - ביות אבולוציוני. הוא איבד חלק מהתכונות ההרסניות של transposase והותאם לתפקיד מבוקר במערכת החיסון.
הבדלים מרכזיים בין transposase לבין RAG:
- RAG דורש זיהוי זוגי של שני רצפים, ולא רק רצף יחיד.
- RAG עובד לפי כלל 12/23.
- RAG אינו מבצע טרנספוזיציה מחדש של המקטע למקום אחר בגנום.
- RAG מגייס את מנגנון NHEJ לתיקון הקצוות.
- RAG מגן על הקצוות שנוצרו, כולל יצירת hairpin.
כך אנזים שמקורו במנגנון נייד ומסוכן הפך לכלי מבוקר ליצירת מגוון חיסוני.
הצד המסוכן של RAG
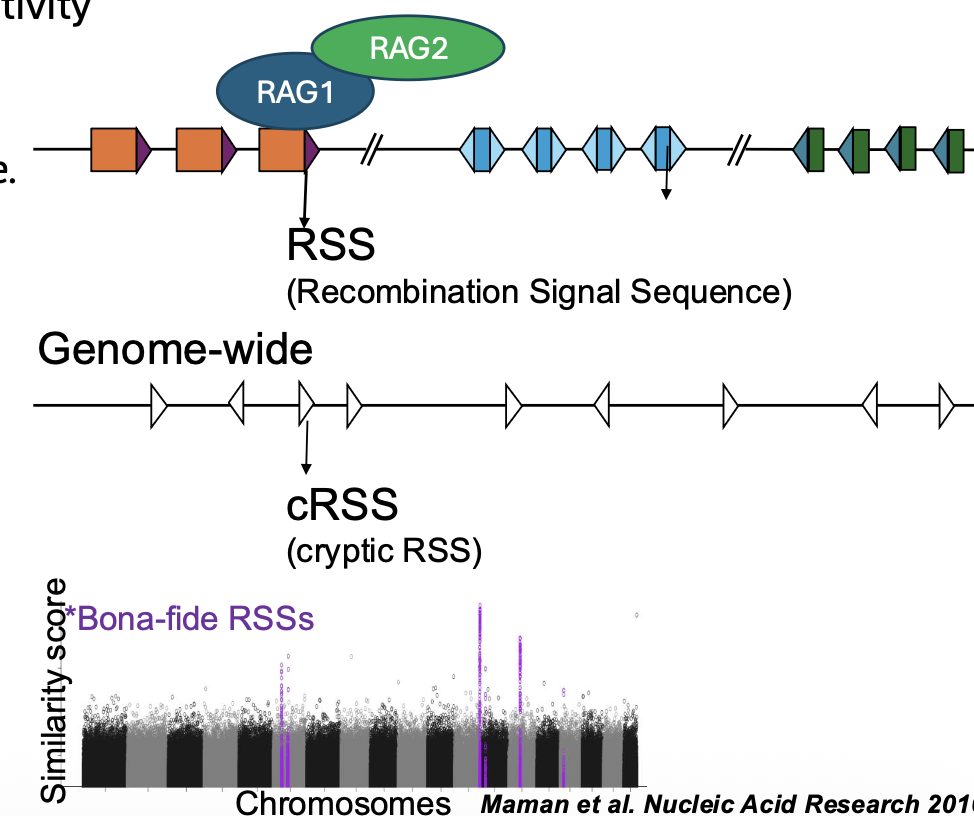
RSS הוא רצף עם חלקים שמורים וחלקים פחות שמורים. בגלל זה קיימים בגנום הרבה רצפים שדומים ל־RSS, גם אם הם לא אמורים להשתתף ב־V(D)J recombination.
רצפים כאלה נקראים cryptic RSS.
הבעיה: אם RAG מזהה בטעות שני cryptic RSS, הוא עלול לחתוך אזור שלא אמור להיחתך. אם שני רצפים כאלה נמצאים סביב tumor suppressor gene, התוצאה יכולה להיות מחיקה של הגן.
זהו אחד ההסברים לפוטנציאל הסרטני של RAG, בעיקר בסרטנים לימפואידיים.
למה זה לא קורה כל הזמן?
אם יש הרבה cryptic RSS, למה RAG לא יוצר נזק בכל הגנום?
התשובה היא שיש עוד שכבות של ספציפיות. RAG לא מסתמך רק על רצף RSS. הוא מגויס בעדיפות לאזורים מסוימים בכרומטין.
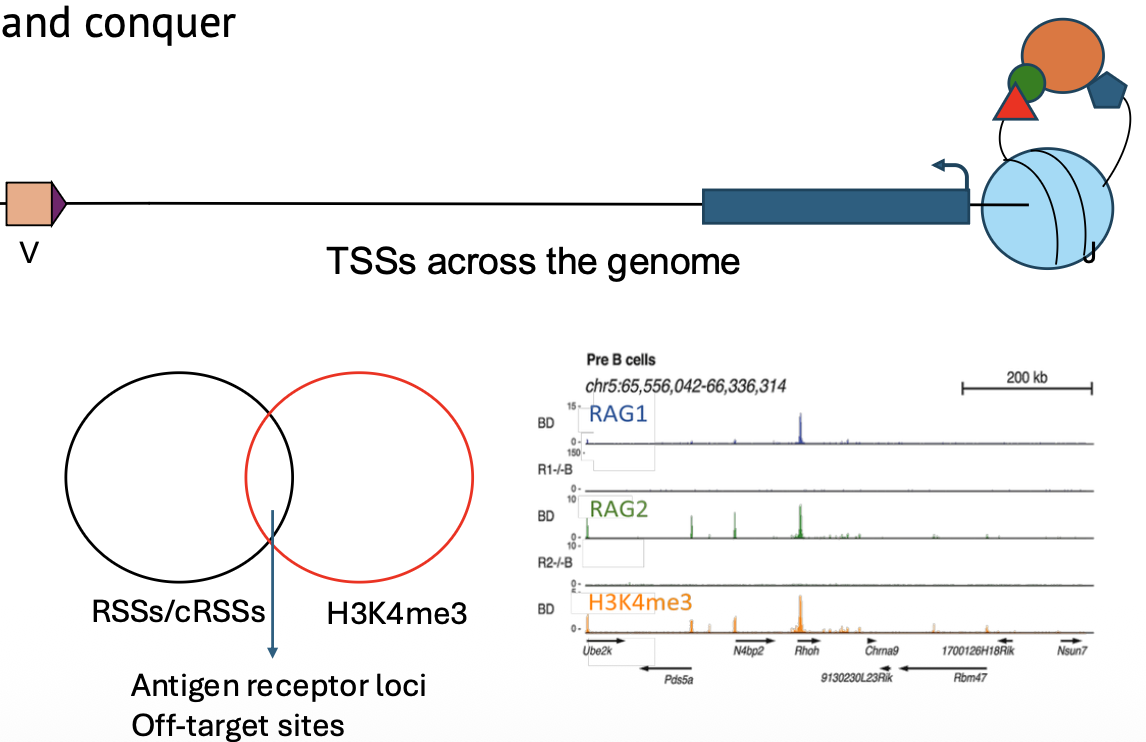
אחד הסמנים החשובים הוא H3K4me3 - טרימתילציה של ליזין 4 בהיסטון H3. הסמן הזה מופיע בפרומוטורים, והוא עשיר במיוחד באזורי recombination center בלוקוסים של antigen receptors.
RAG, ובפרט RAG2, נקשר לאזורים עם H3K4me3. לכן כדי ש־RAG יפעל ביעילות, לא מספיק שיהיה רצף דומה ל־RSS; צריך גם סביבה כרומטינית מתאימה.
ועדיין, זה לא מסביר את כל הספציפיות. יש אזורים בגנום שיש בהם גם רצפים דמויי RSS וגם סימנים כרומטיניים מתאימים, ובכל זאת כמעט אין בהם חיתוך. כלומר, קיימים כנראה רכיבי בקרה נוספים.
במדידות של קישור RAG מול שברים שנוצרים בפועל רואים ש־RAG יכול להיקשר להרבה מקומות, אבל חיתוך משמעותי קורה בעיקר בלוקוסים הנכונים. כלומר, קישור לבד אינו מספיק; צריך תנאים נוספים כדי שהאתר יעבור רקומבינציה.
RAG ולוקמיות
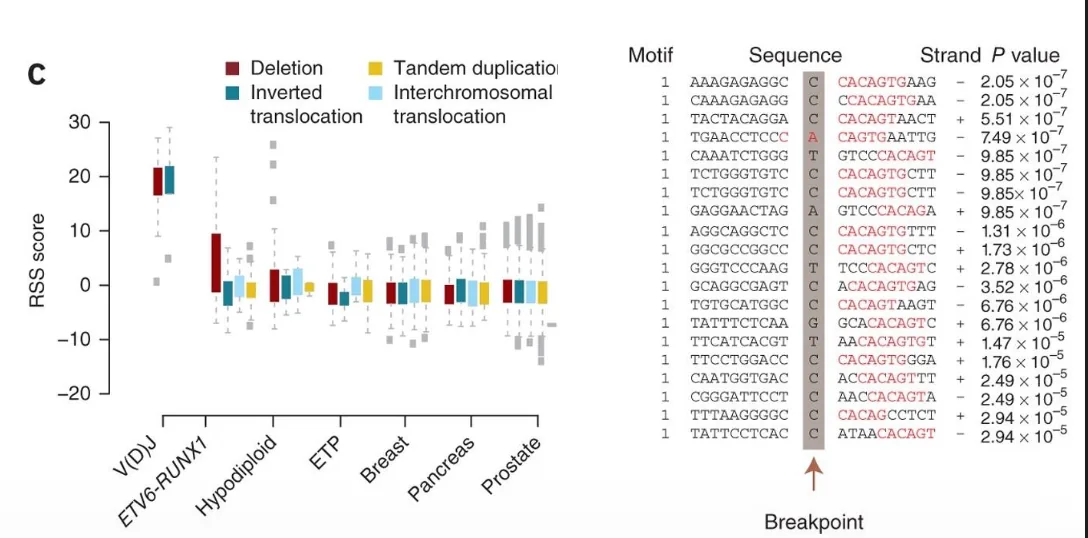
כאשר RAG פעיל מדי או מבוטא בזמן לא נכון, עולה הסיכון לחיתוכים לא רצויים.
דוגמה שנדונה היא B-cell acute lymphoblastic leukemia - B-ALL עם רקע גנטי של ETV6-RUNX1. ברקע כזה RAG יכול להיות פעיל מדי, ואז מחיקות גנטיות רבות מופיעות ליד רצפים שדומים ל־RSS.
כלומר, חלק מהשינויים הגנטיים בלוקמיות כאלה יכולים לנבוע מפעילות RAG מחוץ למקום הרצוי.
Somatic hypermutation ו־AID
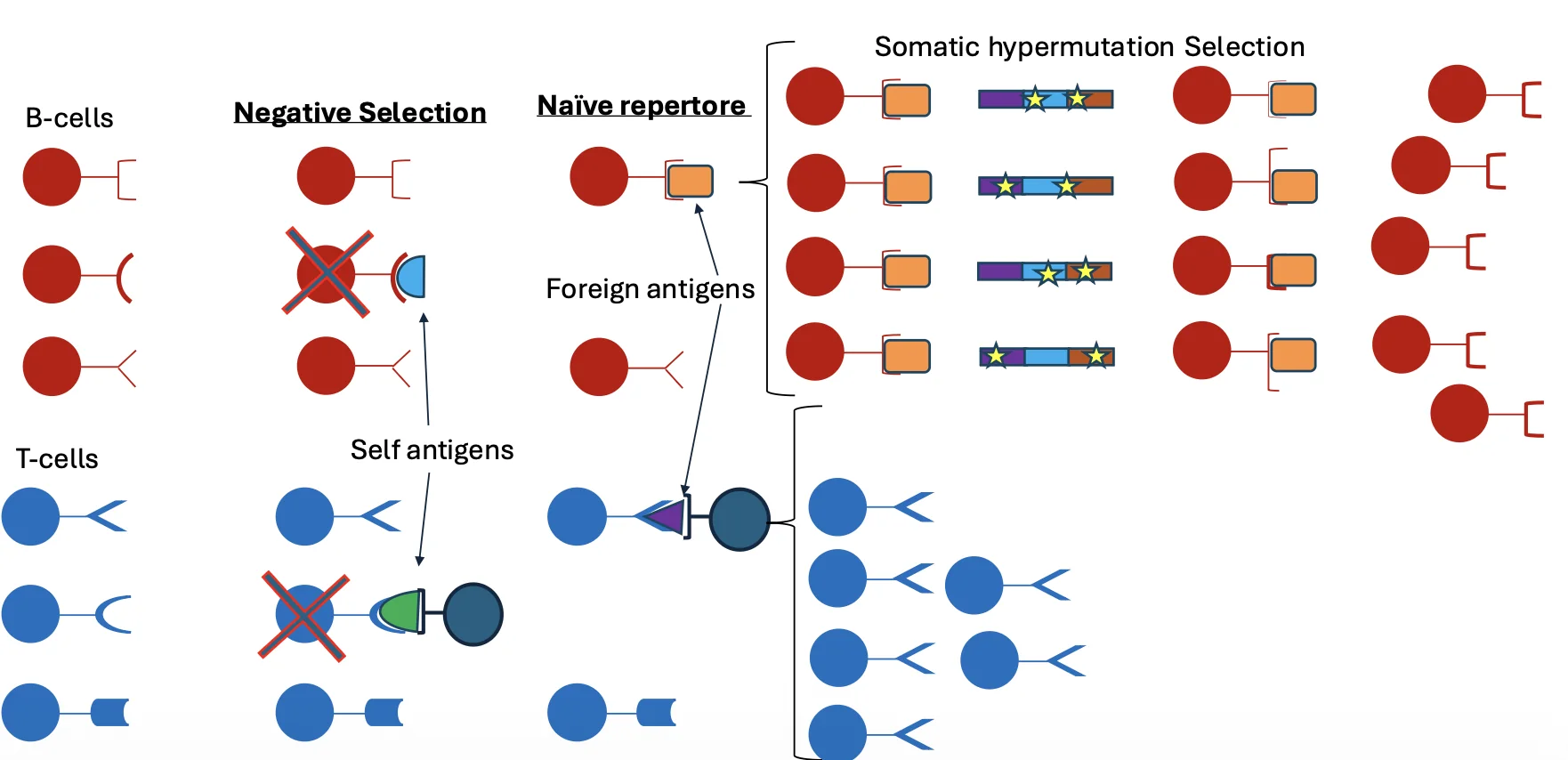
לאחר V(D)J recombination כבר יש רפרטואר גדול של תאי B, וכל תא מבטא נוגדן אחר. השלב הבא הוא affinity maturation: שיפור האפיניות של נוגדן לאנטיגן מסוים לאחר חשיפה לאותו אנטיגן.
השלב הזה מתרחש בתאי B, לא בתאי T.
בתהליך הזה נוצרים שינויים נקודתיים באזורי האימונוגלובולינים. חלק מהמוטציות פוגעות בקישור, חלק לא משנות הרבה, וחלק משפרות את הקישור לאנטיגן. תאים שבהם הנוגדן השתפר שורדים ומתרבים יותר.
זה הבסיס לזיכרון חיסוני: בפעם הבאה שהגוף יפגוש את אותו אנטיגן, כבר יהיו תאים עם נוגדנים טובים יותר.
AID מתחיל את התהליך
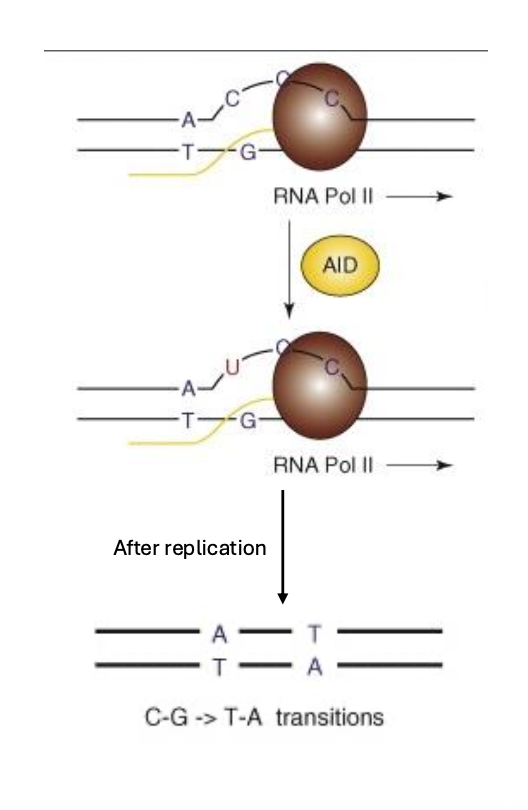
האנזים המרכזי הוא AID - Activation-Induced Deaminase.
AID עושה דיאמינציה של ציטוזין:
\[\ce{C -> U}\]AID מעדיף DNA חד־גדילי. לכן הוא פועל בעיקר באזורים שעוברים שעתוק, שבהם הגדילים נפתחים לזמן קצר.
בניגוד ל־RAG, ל־AID אין רצף זיהוי חד וברור כמו RSS. יש לו העדפה חלשה לקונטקסטים מסוימים, למשל מוטיב שנקרא WRCY, אבל זו אינה ספציפיות חזקה. לכן השאלה המרכזית היא איך AID גורם למוטציות בעיקר באימונוגלובולינים ולא בכל הגנום.
איך דיאמינציה הופכת למוטציה?
דיאמינציה בפני עצמה אינה בהכרח מוטציה קבועה. היא הופכת למוטציה כאשר התא משכפל או מתקן את האזור בצורה שמקבעת שינוי ברצף.
יש שלוש דרכים עיקריות:
1. שכפול מעל U
אם C עבר דיאמינציה ל־U, והשכפול מתרחש לפני תיקון, הפולימראז יכול לקרוא את U כמו בסיס שמזדווג עם A. כך השינוי מתקבע כמוטציה לאחר שכפול.
2. Base Excision Repair עם פולימראזות error-prone
במצב רגיל, BER מתקן דיאמינציה. אבל ב־somatic hypermutation, מנגנון התיקון מנוצל ליצירת מוטציות.
במקום תיקון נקי ומדויק, מגויסות פולימראזות פחות מדויקות. כך נוצרת אפשרות להכניס מוטציות בזמן השלמת האזור.
3. Mismatch Repair עם Polymerase η
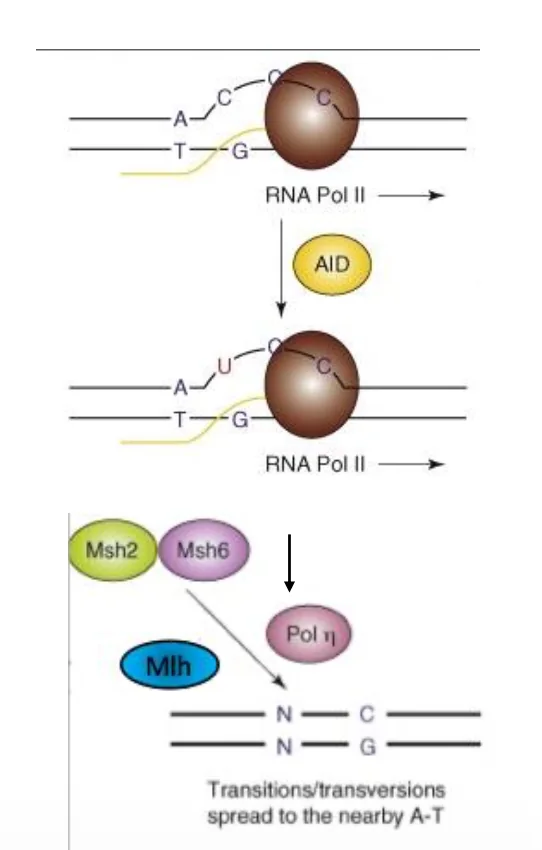
גם MMR יכול לזהות את ה־U/G mismatch שנוצר לאחר דיאמינציה. במקרה כזה מוסר מקטע שלם, ולא רק בסיס אחד.
כאשר המקטע מסונתז מחדש על ידי Polymerase η, יכולות להיכנס מוטציות גם בבסיסים סמוכים. לכן חלק גדול מהמוטציות בתהליך הזה מופיע דווקא ב־A ו־T, ולא רק ב־C.
זה שימוש חריג במנגנוני תיקון: BER ו־MMR, שבדרך כלל נועדו לשמור על הגנום, מנוצלים כאן כדי ליצור גיוון מכוון.
איך AID נשאר יחסית ספציפי?
AID מסוכן לפחות כמו RAG, אולי אפילו יותר מבחינת הספציפיות. הוא יכול עקרונית לעשות דיאמינציה בכל C שנחשף כחד־גדילי. לכן חייבים להבין למה רוב המוטציות מופיעות באימונוגלובולינים ולא בכל גן משועתק.
שעתוק פעיל וחד־גדיליות
AID צריך DNA חד־גדילי. לכן התנאי הראשון הוא active transcription.
אבל שעתוק לבדו לא מספיק. יש הרבה גנים משועתקים שלא עוברים somatic hypermutation ברמה גבוהה.
מה שמגביר את הסיכוי לפעילות AID:
- שעתוק גבוה.
- Pol II stalling - פולימראז RNA II נתקע או מתקדם לאט, ולכן הגדיל החד־גדילי נשאר חשוף יותר זמן.
- R-loops - ה־RNA שנוצר נשאר מזווג ל־DNA, וכך הגדיל השני נשאר חד־גדילי.
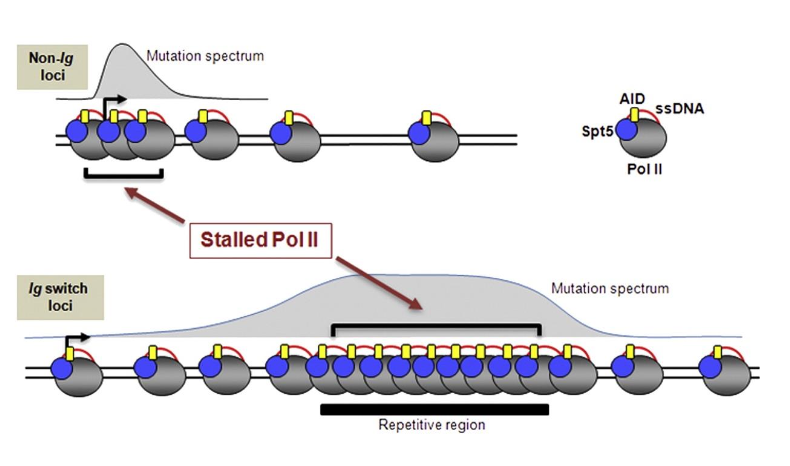
באזורי אימונוגלובולינים יש רמות גבוהות של שעתוק, Pol II stalling ו־R-loop formation. כל אלה הופכים אותם לאזורים נוחים יותר לפעילות AID.
הסביבה הגנטית חשובה יותר מהרצף של הגן
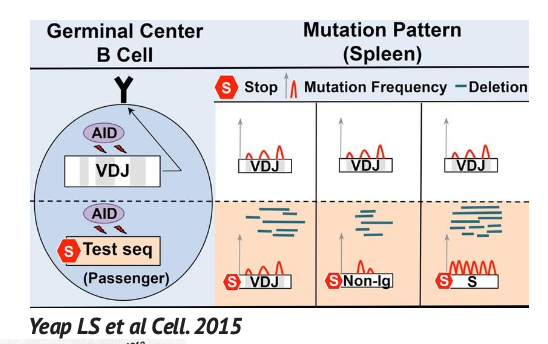
ניסוי חשוב הראה שאם מכניסים רצף בדיקה במקום לוקוס אימונוגלובולין, גם הרצף הזר יכול לעבור מוטציות ברמה גבוהה.
המסקנה: הספציפיות של somatic hypermutation אינה תלויה רק ברצף של הגן עצמו. היא תלויה גם בסביבה הגנטית והרגולטורית שבה הגן נמצא.
אינהנסרים של אימונוגלובולינים
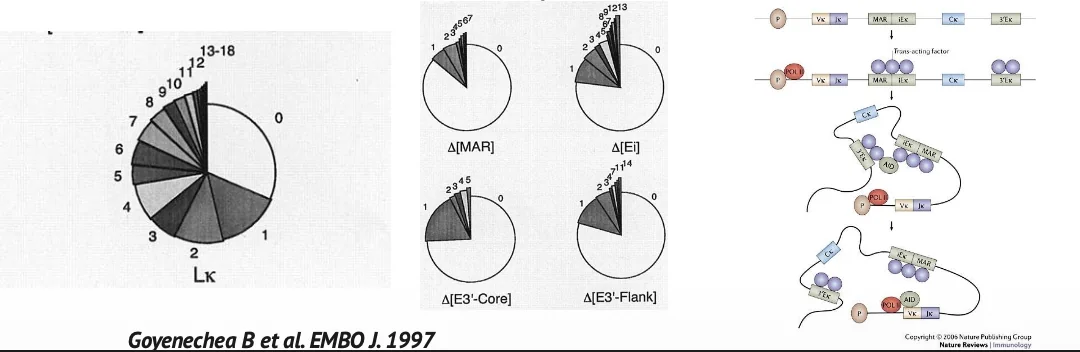
אינהנסרים באזורי אימונוגלובולינים חשובים מאוד ל־somatic hypermutation.
כאשר מסירים חלק מהאינהנסרים, רמת המוטציות יורדת. זה אומר שהאינהנסרים לא רק מגבירים שעתוק; הם יוצרים סביבה רגולטורית שמאפשרת פעילות AID וצבירת מוטציות.
עדיין לא ברור לגמרי מהו הקוד המדויק. ייתכן שהאינהנסרים משפיעים על Pol II stalling, על R-loops, על מבנה הכרומטין, או על גיוס עקיף של פקטורים נוספים.
אזורים חמים, DIVAC ומיפוי פעילות AID
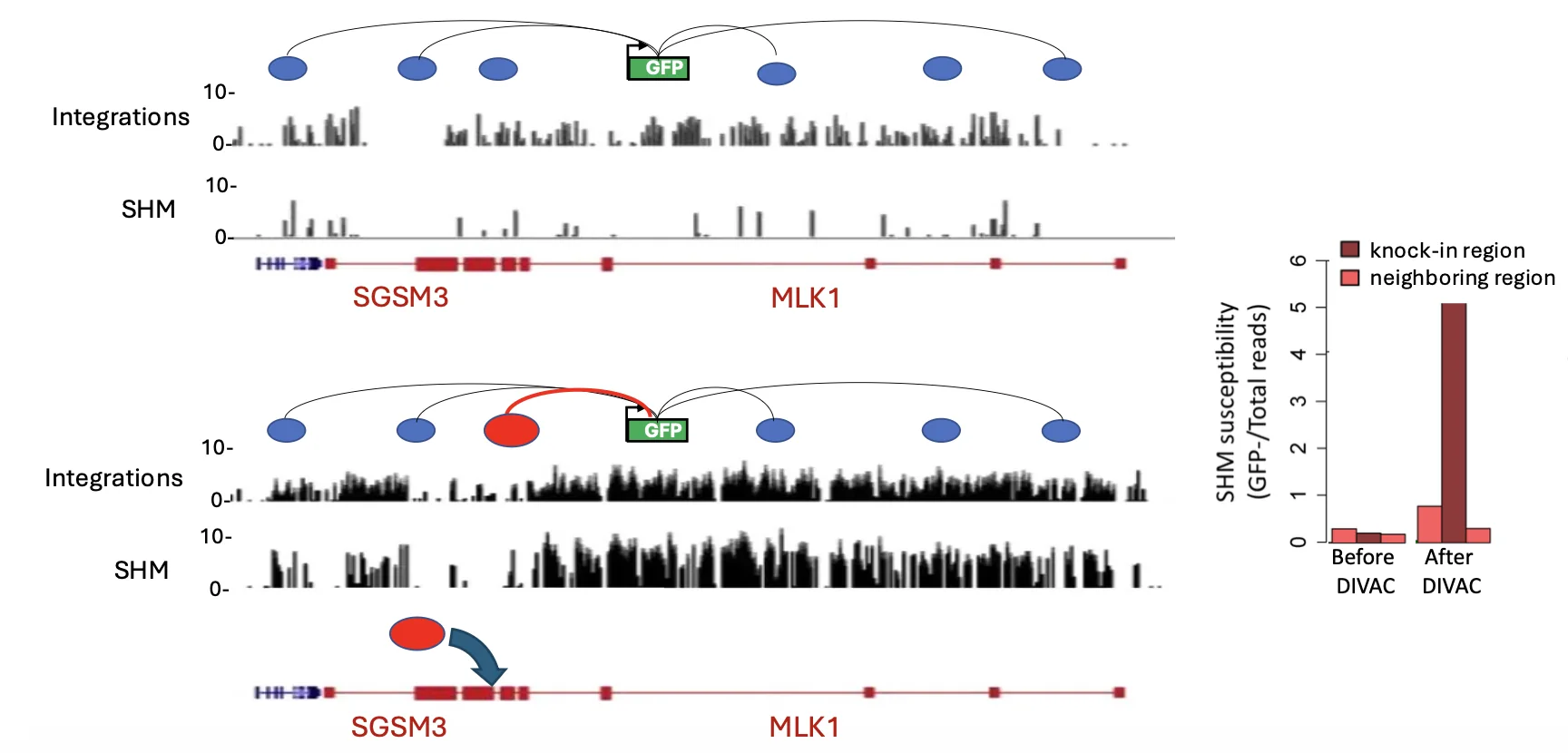
לא כל הגנום רגיש באותה מידה ל־AID. יש אזורים “חמים” שבהם יש הרבה somatic hypermutation, ואזורים “קרים” שבהם יש מעט מאוד.
בהתחלה אפשר לחשוב שההבדל הוא רק רמת השעתוק, אבל זה לא מספיק. אזורים חמים וקרים יכולים להיות משועתקים ברמה דומה, ובכל זאת לעבור רמות שונות מאוד של מוטציות.
אחד ההבדלים שנמצאו קשור לאינטראקציות עם אינהנסרים. אזורים חמים נוטים להיות באינטראקציה עם הרבה אינהנסרים, בין השאר דרך גורמים שקשורים לארגון הכרומטין ול־loops.
DIVAC
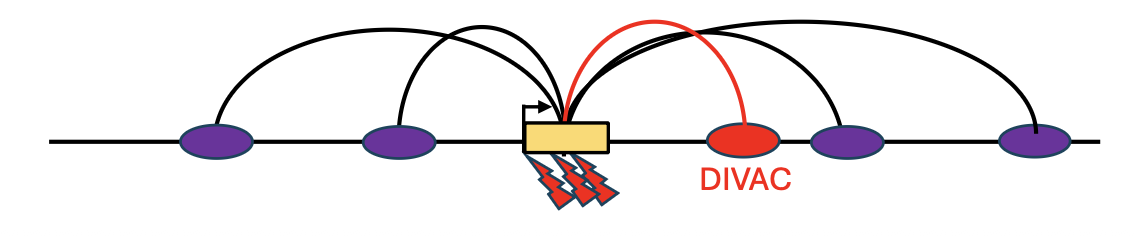
DIVAC - Diversification Activator הוא מקטע רגולטורי קצר יחסית שמקורו באינהנסרים של אימונוגלובולינים, ויכול לעודד somatic hypermutation באזור שבו הוא נמצא.
העיקרון הניסויי:
- לוקחים מקטע רגולטורי קטן מאזורי אימונוגלובולינים.
- מחברים אותו לגן דיווח כמו GFP.
- מכניסים את המבנה לתאים שמבטאים AID.
- אם GFP מאבד פעילות, כלומר התאים מפסיקים לזרוח בירוק, זה סימן שנוצרו מוטציות שפגעו ב־GFP.
כך אפשר למדוד אילו מקטעים רגולטוריים מסוגלים להפוך אזור “קר” לאזור “חם” מבחינת somatic hypermutation.
מיפוי אזורים חמים בעזרת GFP
גישה נוספת היא להכניס GFP למקומות שונים בגנום בעזרת רטרווירוס, בתאים שמבטאים AID, ואז לאסוף תאים שאיבדו את GFP.
אם GFP הפסיק לפעול, סביר שהאזור שבו הוא נכנס הוא אזור שנוטה לעבור מוטציות. לאחר מכן מרצפים וממפים איפה GFP השתלב.
כך ניתן לזהות אזורים חמים וקרים בגנום.
התוצאה הכללית:
- אזורי אימונוגלובולינים הם החמים ביותר.
- יש גם אזורים אחרים בגנום שיכולים להיות חמים יותר מהרגיל.
- אזורים חמים קשורים לאינטראקציות רגולטוריות רבות, בעיקר עם אינהנסרים.
המודל הוא שאזורים עם הרבה אינטראקציות עם אינהנסרים מקבלים באופן מצטבר חלק מהתכונות של DIVAC. הם לא בהכרח מכילים את ה־DIVAC “המלא” של אימונוגלובולינים, אבל הם יכולים לדמות אותו ברמה מסוימת.
דור פסקל