חזרה - חיתוך וחיבור DNA (תהליך ה־Cloning)
נוקלאזות
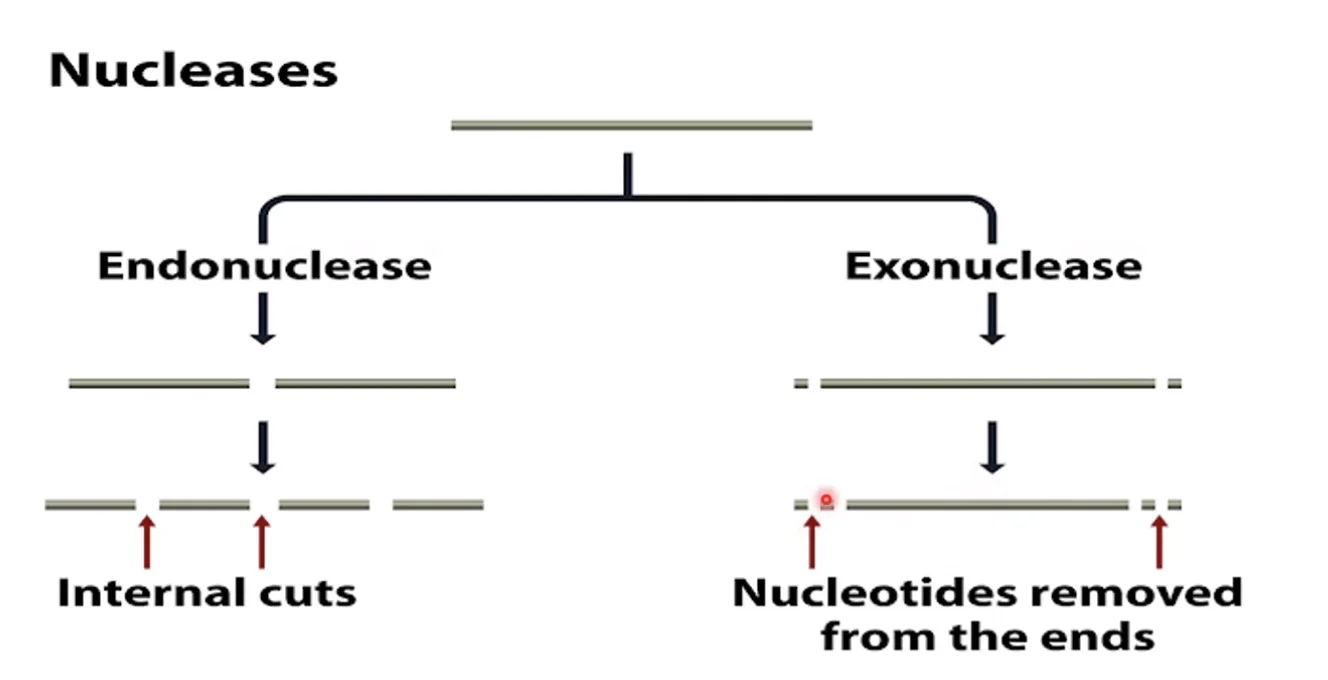
נוקלאז הוא שם כללי לאנזים שיודע לחתוך את השלד הפוספוסוכרי. יש שני סוגים עיקריים:
-
אקסונוקלאז - חותך נוקלאוטיד אחד מהקצה, ויכול להמשיך לחתוך נוקלאוטיד אחר נוקלאוטיד עד שיחסל את כל המולקולה. חשוב לשים לב: אקסונוקלאז לא יוכל לפעול על מולקולה מעגלית - שאין לה קצוות.
-
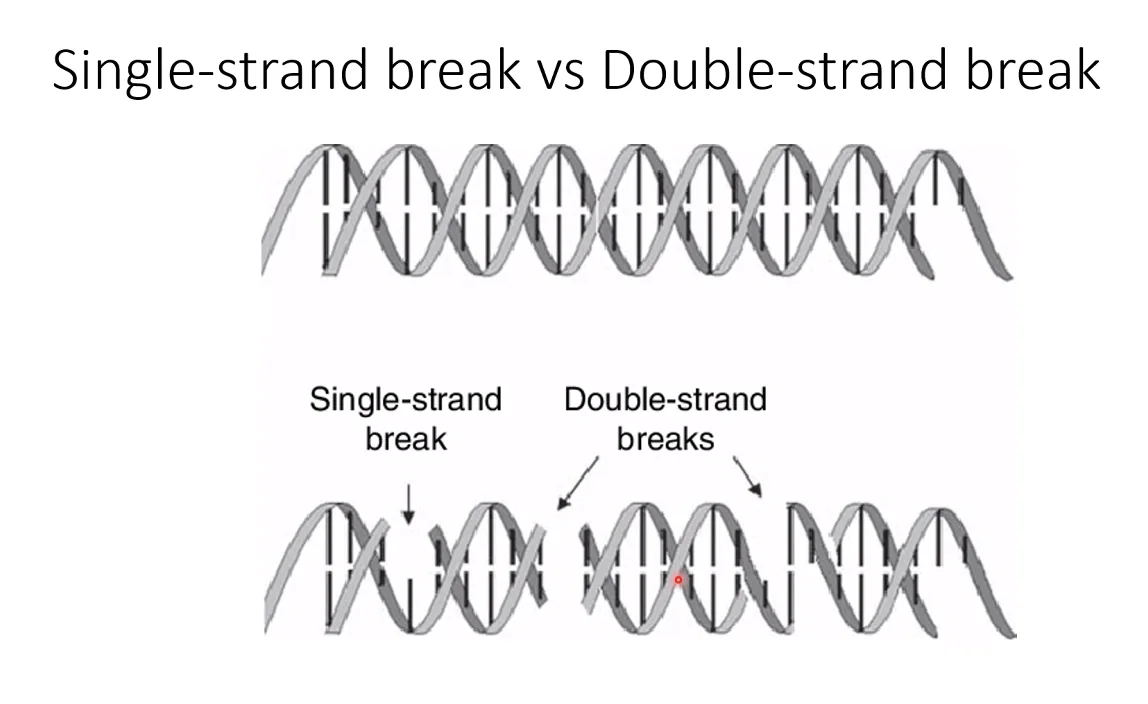
אנדונוקלאז - חותך בתוך המולקולה, לא מהקצה. יכול לחתוך חיתוך חד־גדילי (nick - שבר בשלד אחד בלבד, המולקולה נשמרת שלמה כי זיווג הבסיסים מחזיק את שני הגדילים) או חיתוך דו־גדילי (שבר בשני הגדילים, המולקולה מתפרקת לשני חלקים).
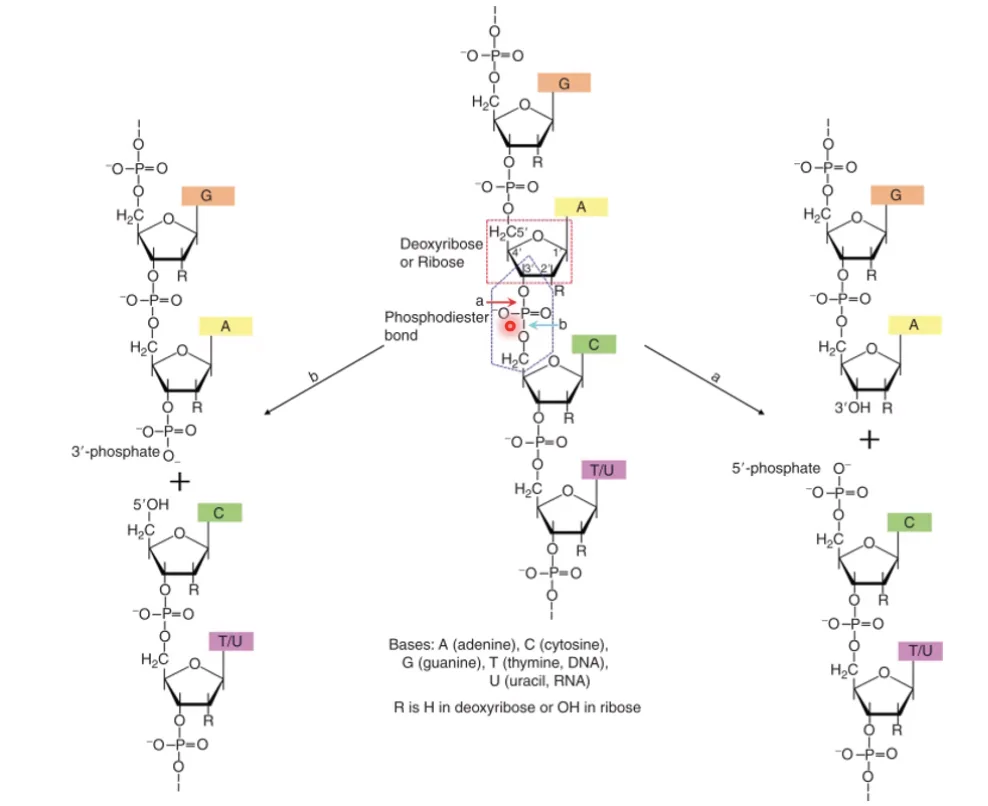
שבר דו־גדילי הוא אחד האירועים הכי מסוכנים לתא. הוא יכול לנבוע מנוקלאזות, מתהליכי רפליקציה, מקרינה או מחשיפה לכימיקלים. אם לא מתוקן - התא הולך לאפופטוזיס. זה הבסיס לטיפולים כימותרפיים וקרינתיים, שמייצרים כמות שברים דו־גדיליים מעבר ליכולת התיקון של התא הסרטני, ומכוונים אותו למוות.
אנזימי רסטריקציה
אנזימי רסטריקציה הם סוג מיוחד של אנדונוקלאזות, שמקורם בחיידקים - שם הם משמשים להגנה מפני וירוסים. מה שמייחד אותם הוא זיהוי רצף ספציפי - הם לא חותכים בכל מקום, אלא רק כשהם מזהים רצף בסיסים מוגדר מראש (לדוגמה, EcoRI מזהה את הרצף GAATTC).
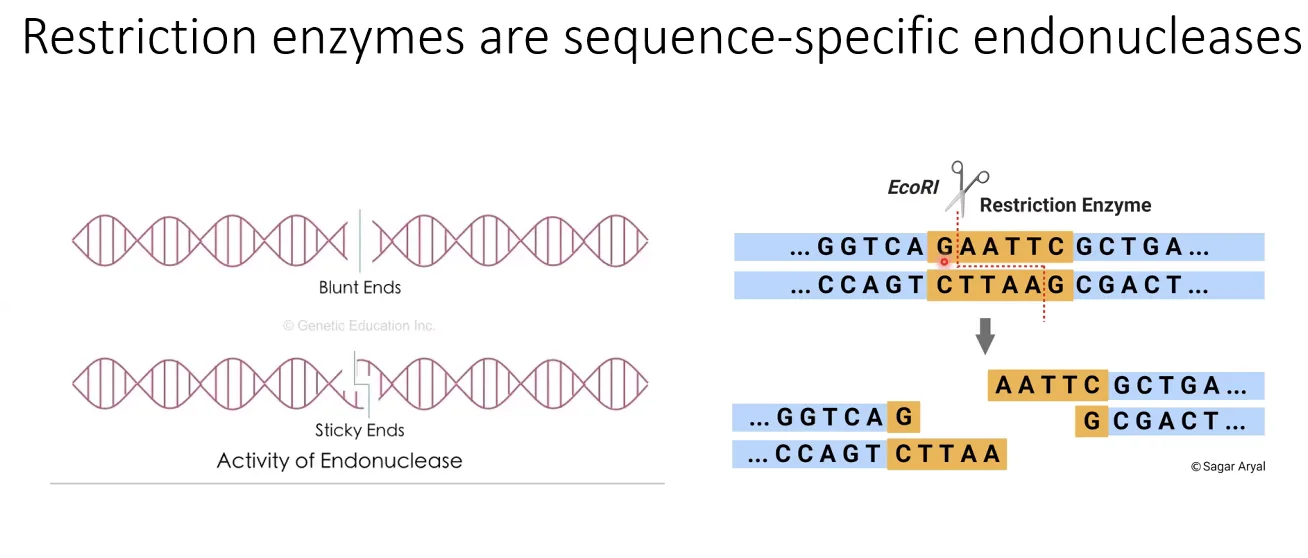
הרצף שמזהה אנזים הרסטריקציה חייב להיות פלינדרומי - כלומר, זהה בשני הגדילים (כשקוראים בכיוון 5’←3’). האנזים נכנס דרך ה־Major Groove, מזהה את הרצף, ועובד כדימר - חותך גדיל אחד מצד אחד וגדיל שני מהצד השני.
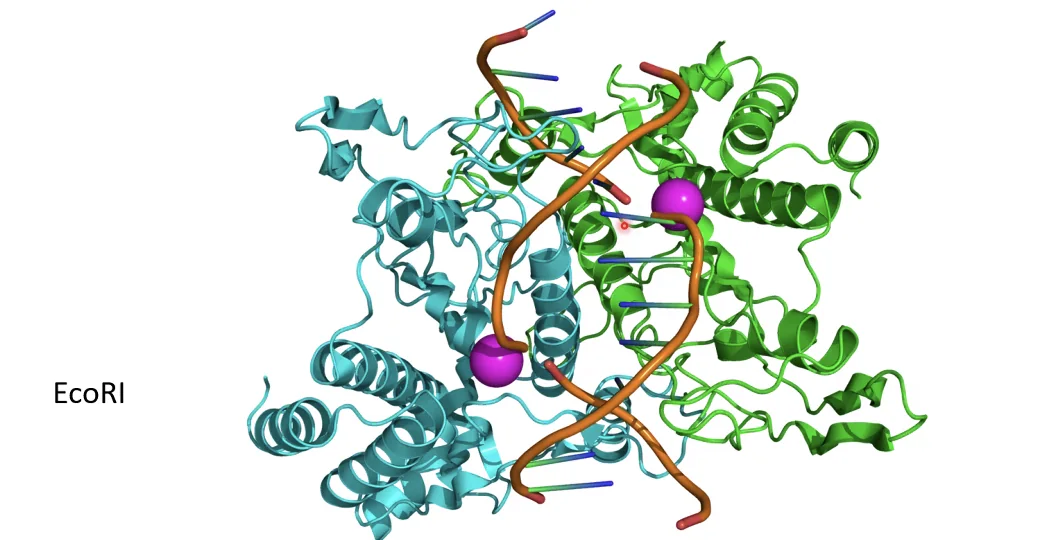
רוב אנזימי הרסטריקציה לא חותכים בנקודה זהה בשני הגדילים, אלא בא-סימטריה - ואז נוצרים Sticky Ends (קצוות דביקים): קטע חד־גדילי קצר שנחשף בשני קצוות החיתוך, משלים זה את זה. מכיוון שהיברידיזציה היא ספונטנית, שני מקטעי DNA שנחתכו באותו אנזים רסטריקציה יעשו היברידיזציה ספונטנית כשמביאים אותם יחד - וזה הבסיס לשיבוט. לעומתם, Blunt Ends (קצוות חלקים) נוצרים כשהחיתוך בשני הגדילים הוא בדיוק בנקודה זהה.
ליגאז
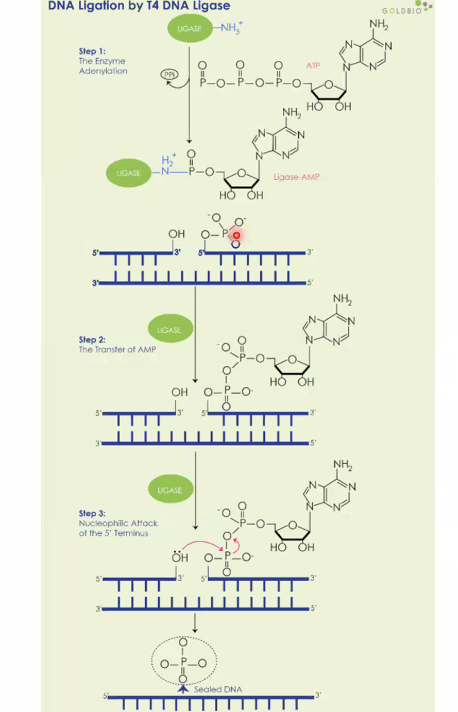
לאחר שמקטעי DNA התחברו זה לזה על ידי היברידיזציה ספונטנית של הקצוות הדביקים, עדיין יש ניקים - שברים חד־גדיליים בשלד הפוספוסוכרי. האנזים ליגאז סוגר אותם. אבל בניגוד לסינתזת DNA רגילה (שבה האנרגיה טמונה בנוקלאוטיד הטריפוספט), כאן יש רק מונופוספט, ולכן אין מספיק אנרגיה לסגירה ספונטנית. הליגאז פורק ATP, משתמש באנרגיה שלו כדי ליצור מצב ביניים עתיר אנרגיה, ואז מבצע התקפה נוקלאופילית שסוגרת את הקשר הפוספודיאסטרי.
דוגמה קלינית: ייצור אינסולין רקומביננטי. לוקחים את הגן האנושי לאינסולין, מכניסים אותו לתוך פלסמיד חיידקי על ידי חיתוך בשני אנזימי רסטריקציה זהים ואז ליגציה, מגדלים חיידקים שמייצרים כמויות עצומות של אינסולין - ומנקים אותו לשימוש קליני. זה הבסיס לכל ביולוגיה מולקולרית רקומביננטית.
טופולוגיה של DNA
מהי טופולוגיה?
טופולוגיה היא ענף מתמטי שמתאר מבנים גיאומטריים במרחב. כשמסתכלים על DNA מנקודת מבט טופולוגית, מבינים שהמולקולה לא תמיד מתנהגת כמו קו ישר - יש משמעות לצורה המרחבית שבה היא מתארגנת. ניתן גם להכניס אנרגיה למולקולת DNA על ידי קיפולה על עצמה.
התנאי הבסיסי להכנסת מתח פיתולי: הקצוות של המולקולה חייבים להיות קבועים/מוחזקים. אם הקצוות חופשיים, כל מתח שנכניס ישתחרר בסיבוב החופשי של המולקולה. יש שתי דרכים עיקריות לקבע קצוות:
- חלבונים שאוחזים בקצוות (היסטונים, קוהיזין, טלומרים) - אופייני לאאוקריוטים עם DNA ליניארי
- מבנה מעגלי - אין קצוות כלל, אופייני לכרומוזומים חיידקיים
שלוש צורות טופולוגיות
-
Knotting (קשירה): מולקולה שנכנסת בתוך עצמה ויוצרת קשר - בדיוק כמו שאנחנו קושרים שרוכים. נדיר, לא נדון בו לעומק.
-
Catenane (קטננט): ⚭ שתי מולקולות DNA מעגליות שמוחזקות יחד כמו שתי טבעות בשרשרת - כל אחת שלמה ונפרדת, אבל לא ניתן לנתק אותן ללא חיתוך. רלוונטיות ביולוגית: זהו תוצר לוואי של רפליקציה - שני כרומוזומים בנות שנוצרים מהרפליקציה יכולים להישאר קשורים כקטננט, ולא ניתן לחלק אותם לשני תאי בת ללא שחרורם. האנזים שעושה זאת הוא טופואיזומראז.
-
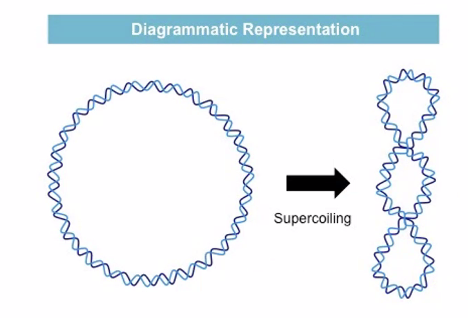
Supercoiling (סופרקויל): קיפול של הדו־גדיל על עצמו - כמו גומייה שמסובבים עד שהיא מתכווצת על עצמה. זה המצב הטופולוגי העיקרי שנדון בו.
סופרקויל - Supercoiling
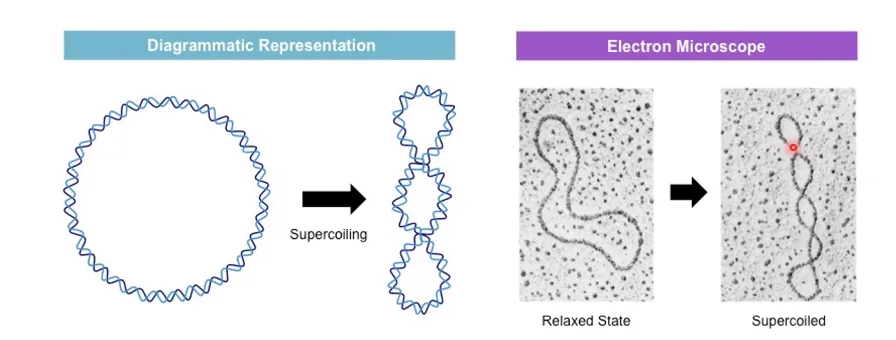
חיובי לעומת שלילי
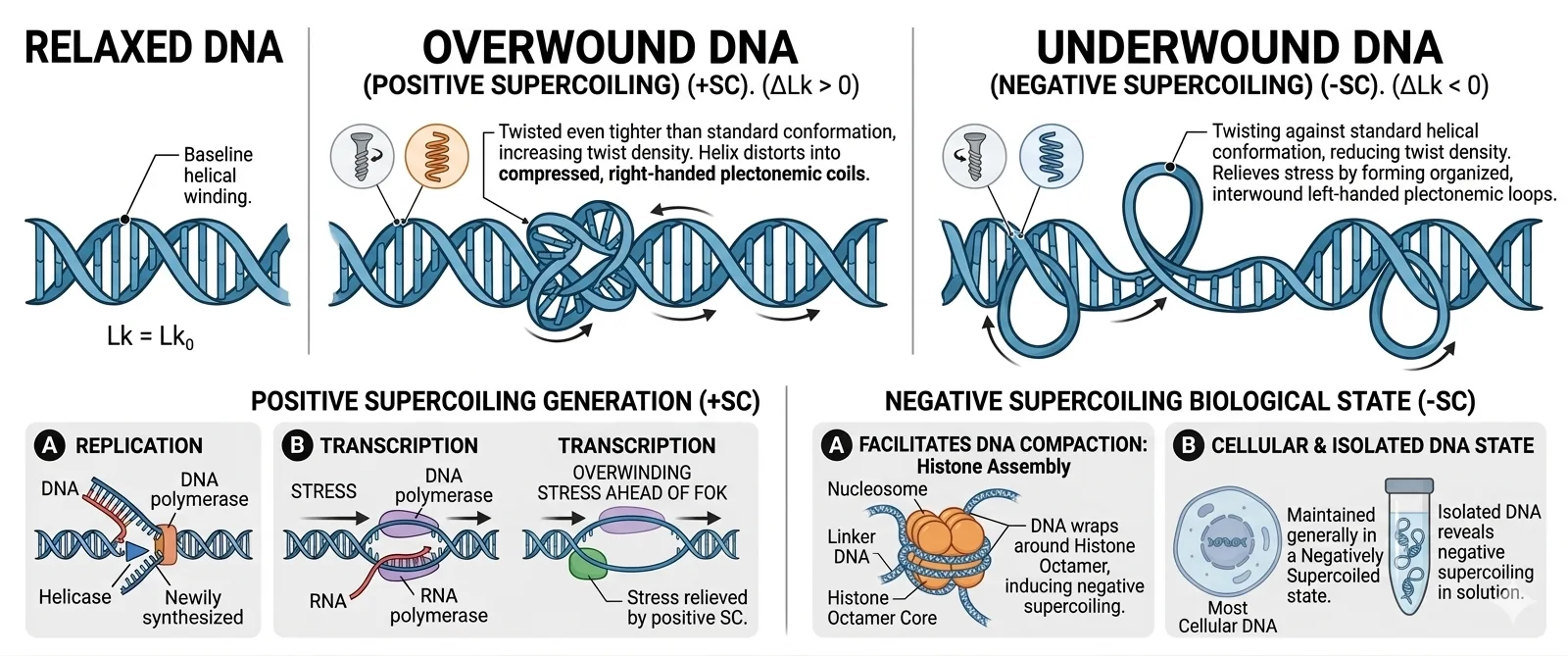
ל־B-DNA יש סיבוב ימני (עם כיוון השעון). כשמכניסים סופרקויל, ניתן לעשות זאת בשני כיוונים:
-
Positive supercoiling (פוזיטיבי): הסיבוב של הסופרקויל הוא ימני - באותו כיוון כמו הגדילים. זה מחזק את הדו־גדיל, מהדק אותו עוד יותר.
-
Negative supercoiling (שלילי): הסיבוב הוא שמאלי - כנגד כיוון הגדילים. זה מרפה את הדו־גדיל, ומקל על פתיחתו לשני חד־גדילים.
מה קיים בטבע? DNA בתאים נמצא בדרך כלל בנגטיב סופרקויל. זה לא המצב האנרגטי היציב ביותר (שהוא מצב relaxed), אבל יש לו משמעות ביולוגית חיונית: DNA שנמצא בנגטיב סופרקויל קל יותר לפתיחה לשני חד־גדילים - וזה הכרחי לטרנסקריפציה ולרפליקציה. לדוגמה, פרומוטורים (אזורים שבהם מתחיל ביטוי גנים) נמצאים תמיד במצב קצת יותר פתוח, וזה מאפשר לגורמי שעתוק (Transcription Factors) להיקשר שם.
סופרקויל פוזיטיבי הוא גם תוצר לוואי של תהליכים ביולוגיים כמו טרנסקריפציה ורפליקציה - כתוצאה מהתקדמות ה־RNA פולימראז, נוצר סופרקויל חיובי לפני הפולימראז ושלילי אחריו. לכן חובה לשחרר את המתח הזה - אחרת ה־DNA ייפרם ויישבר.
להלן איור ספק מדויק להמחשה, מג׳מניי:
המספרים הטופולוגיים (Lk, Tw, Wr)
על מנת לתאר את המצב הטופולוגי של מולקולת DNA באופן מדויק, משתמשים בשלושה פרמטרים:
-
Tw - Twisting Number (מספר הסיבובים): מספר הפעמים שחד־גדיל אחד מלופף סביב חד־הגדיל השני. זה בעצם מבנה הדו־גדיל עצמו - בתנאים פיזיולוגיים, הערך הוא בערך 10.5 בסיסים לסיבוב הליקלי שלם (לצרכי חישוב: 10). ה־Tw תמיד חיובי ב־
B-DNA. מבחינה אנרגטית, הוא תמיד שואף לחזור לערך “ברירת המחדל” שלו (אורך מולקולה חלקי 10). -
Wr - Writhing Number (מספר ה־supercoils, ״מתח פיתולי״): מספר הפעמים שהדו־גדיל חוצה את עצמו. במצב relaxed, Wr=0. בסופרקויל חיובי: Wr חיובי; בסופרקויל שלילי: Wr שלילי.
-
Lk - Linking Number (המספר הטופולוגי): הסכום הכולל: Lk = Tw + Wr. זה “המספר הטופולוגי” של המולקולה - הוא תמיד מספר שלם, תמיד גדול מאפס (אפס = שני חד־גדילים נפרדים, לא דו־גדיל), ומייצג את מצב המולקולה כולו.
החוק החשוב ביותר
כל עוד השלד הפוספוסוכרי סגור - ה־Lk לא משתנה. כלומר, אפשר לשחק עם Tw ועם Wr כמה שרוצים, אבל אם המולקולה סגורה, כל שינוי ב־Tw ישפיע בדיוק באותה מידה על Wr (ולהפך) - ו־Lk יישאר קבוע. ניתן לשנות את Lk רק על ידי חיתוך וחיבור מחדש של השלד הפוספוסוכרי.
דוגמאות חישוב
שאלה בסיסית: מולקולה דו־גדילית באורך 5,000 זוגות בסיסים במצב relaxed - מה ה־Lk?
\[\text{Tw} = \frac{\text{length}}{\text{base per turn}} = \frac{5000}{10} = 500\]שאלה מורכבת: מולקולה של 10,000 זוגות בסיסים עם Wr = −500 - מהו ה־Lk?
\[\text{Lk} = \text{Tw} + \text{Wr} = 1000 + (-500) = 500\](נגטיב סופרקויל חזק).
הוסיפו אנזים ששינה את ה־Lk ב $+100$: מה ערכי הפרמטרים החדשים?
\[\text{Lk new} = \text{Lk old} + 100 = 500 + 100 = 600\] \[\text{Tw new} = \text{Tw old} = 1000\] \[\text{Wr new} = \text{Lk new} - \text{Tw new} = 600 - 1000 = -400\]השפעת נוקלאזות על טופולוגיה
-
אנדונוקלאז על מולקולה מעגלית: חותך את השלד - Wr מתאפס (כל המתח הפיתולי משתחרר), והמולקולה הופכת לליניארית. Lk = Tw.
-
אקסונוקלאז על מולקולה מעגלית: לא קורה שום דבר - המולקולה מעגלית, אין לה קצוות לחתוך מהם.
-
Nick (שבר חד־גדילי) על מולקולה מעגלית: גם nick אחד מספיק לאפס את Wr ולשחרר את כל המתח הפיתולי, אך המולקולה שומרת על צורתה המעגלית (כי שני הגדילים עדיין מחוברים).
-
חימום מולקולה ליניארית: הגדילים נפרדים, Tw ← 0, לא נותר מצב טופולוגי.
-
חימום מולקולה מעגלית: הגדילים לא יכולים להיפרד לגמרי - מקבלים קטננט ⚭: שתי לולאות מעגליות המשולבות זו בזו, עם Lk = 1.
טופואיזומראזות
טופואיזומראזות הם האנזימים שיודעים לשנות את ה־Lk של מולקולת DNA. הם עושים זאת על ידי חיתוך מבוקר של השלד הפוספוסוכרי, שינוי מבני, וחיבור מחדש. נדון בהם בהרחבה בשיעורים הבאים.
ג’ל אלקטרופורזה
עיקרון הטכניקה
אלקטרופורזה היא הרצה של מולקולות בג’ל לפי גודלן בשדה חשמלי. DNA הוא מולקולה שלילית מאוד (בגלל קבוצות הפוספט בשלד), ולכן כשמפעילים שדה חשמלי, מולקולות ה־DNA ינועו לכיוון האנוד (הצד החיובי). ככל שמולקולה קטנה יותר - קל לה יותר לעבור בנקבי הג’ל, היא נעה מהר יותר ועוברת מרחק גדול יותר.
ג’ל אגרוז
ג’ל האגרוז עשוי מאגרוז - פוליסכריד שמופק מאצות. כשמחממים אגרוז הוא נמס; כשמקררים אותו הוא יוצר קשרי מימן בין הסוכרים ומתמצק ליצירת רשת בעלת נקבים (פורות). ריכוז האגרוז קובע את גודל הנקבים: ריכוז נמוך (0.7%) נותן נקבים גדולים ורזולוציה טובה לגדלים גדולים; ריכוז גבוה (1.5%) נותן נקבים קטנים ורזולוציה טובה לגדלים קטנים. ריכוז סטנדרטי לעבודה: 0.8%–1%.

ויזואליזציה
DNA בעצמו שקוף - לא ניתן לראות אותו בעין. לצביעה משתמשים בשתי שיטות עיקריות:
-
אתידיום ברומיד (EtBr): מולקולה פלואורסנטית שעוברת אינטרקלציה - נכנסת בין מדרגות הסולם של ה־DNA. הנוכחות שלה בתוך ה־DNA משנה את הזווית של השלד בכ־26 מעלות לכל מולקולה שנכנסת, ובכך משפיעה על ה־Tw. שימושית לראיית DNA על ג’ל, אך מוטגנית ומצריכה זהירות. חשוב: הרבה תרופות כימותרפיות פועלות על ידי אינטרקלציה לתוך DNA של התאים הסרטניים, גורמות לעיוות מבני, ומובילות לאפופטוזיס.
-
SYBR Green: נקשר ל־Major Groove (לא עושה אינטרקלציה), פלואורסנטי. משמש ב־qPCR - כמות הפלואורסנציה פרופורציונלית לכמות ה־DNA, ולכן ניתן לעקוב בזמן אמת אחרי ההגדלה האקספוננציאלית של ה־DNA בתהליך ה־PCR. כשמרתיחים את ה־DNA לדנטורציה, ה־SYBR Green משתחרר (אין לו במה להיקשר), ולכן הפלואורסנציה יורדת - ועולה שוב עם הסינתזה של הגדיל החדש.
ג’ל אלקטרופורזה ומצבים טופולוגיים

ג’ל אגרוז מפריד DNA לפי גודל וצורה. לאותה מולקולה עם מספר בסיסים זהה יהיה ניוד שונה, כתלות במצב הטופולוגי שלה:
-
סופרקויל (closed circular): הצורה הכי קומפקטית — נעה הכי מהר (הרחוק ביותר מנקודת הטעינה).
-
Linear (ליניארי - אחרי חיתוך דו־גדילי): נעה במהירות בינונית.
-
Relaxed circular (nicked - אחרי שבר חד־גדילי): המעגל שחרר את המתח (Wr=0), אבל שמר על צורתו המעגלית. מבחינה מרחבית קשה לו יותר לעבור דרך נקבי הג’ל — הוא נע הכי לאט, ונמצא הכי גבוה בג’ל.
לכן, כשמפיקים פלסמיד מחיידק ורואים שני בנדים בג’ל (כאשר הם מתחילים מלמעלה ומתפשטים מטה) - זה נורמלי:
- הבנד התחתון - הוא ה־DNA במצב סופרקויל (טבעי בתא).
- הבנד העליון - הוא ה־DNA שספג ניקים בתהליך ההפקה, ועבר לצורת relaxed circular.
Pulsed-Field Gel Electrophoresis (PFGE)
DNA גדול מאוד (כמו כרומוזום שלם) לא יכול להיכנס לנקבי ג’ל אגרוז רגיל - הוא ייתקע בכניסה. הפתרון הוא PFGE - שינוי תמידי של כיוון השדה החשמלי. המולקולה מתנהגת כמו נחש, כל הזמן משנה כיוון ועוברת בפורות. בצורה כזאת ניתן להפריד אפילו כרומוזומים שלמים - כמו ב־karyotyping של שמר, המפריד בין DNA בגדלים של 50,000 עד 400,000 זוגות בסיסים.
דור פסקל