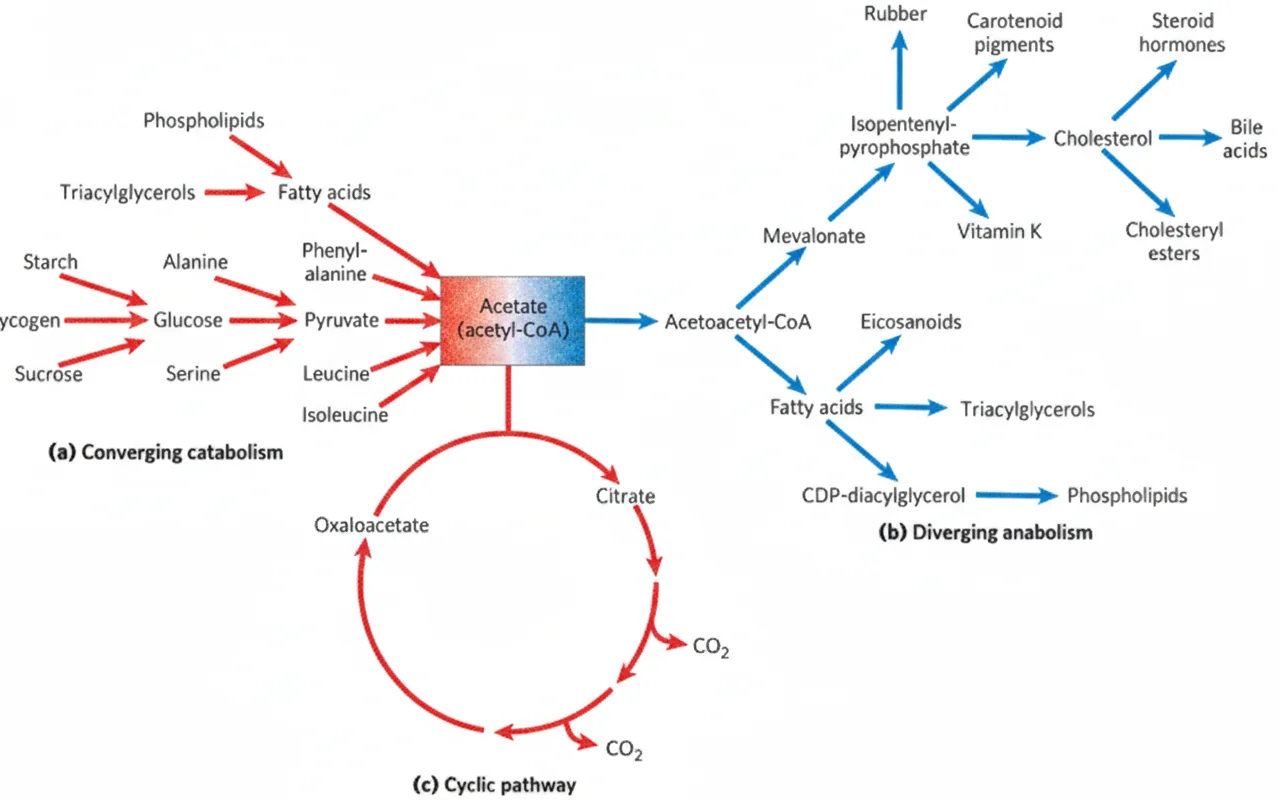
מבוא: קטבוליזם ואנאבוליזם
בשיעורים הקודמים דיברנו על קטבוליזם - פירוק של סוכרים (גליקוליזה), פירוק של חומצות אמיניות (מעגל האוריאה, טרנסאמינציה, דאמינציה אוקסידטיבית), ופירוק חומצות שומן (β-oxidation). כל המסלולים האלה מתנקזים בסופו של דבר ל־acetyl-CoA.
ל־acetyl-CoA יש שתי אפשרויות עיקריות:
- כניסה למעגל קרבס - שם הוא הופך לאנרגיה: $\ce{GTP}$ אחד (בתוך המעגל), וכוחות מחזרים $\ce{NADH}$ ו־$\ce{FADH2}$
- שימוש כאבן בניין - ל־Fatty Acids (ומהם לטריגליצרידים ולאיקוזנואידים), או למבלונאט (ומשם לכולסטרול)
היום נתמקד בעיקר בסינתזה של שומנים.
 איור לא מדויק של GPT
איור לא מדויק של GPT אנאבוליזם לעומת קטבוליזם
נקודה חשובה: אנאבוליזם זה לא אותן ריאקציות שהולכות פשוט בשיווי משקל לצד השני. במהלך הפירוק יש שלבי התחייבות (commitment steps) - שלבים מאוד אקסרגוניים שדורשים $\ce{ATP}$ והשקעת אנרגיה. ככלל, שלבי ההתחייבות הם בלתי הפיכים (irreversible) - כלומר, אי אפשר פשוט לחזור עליהם בכיוון הנגדי.
כשרואים בתרשים מטבולי חץ אחד בלבד (ולא שני חיצים בשני כיוונים), זה אומר שמדובר בשלב מחייב ובלתי הפיך. לכן, כשהולכים בכיוון ההפוך (בנייה), חייבים לעקוף את השלבים הרגולטוריים האלה דרך מסלול אחר.
ההבדלים העיקריים בין פירוק לבנייה
מבחינה קונספטואלית, אם ריאקציות של קטבוליזם הן ריאקציות של חמצון ופירוק, אז בריאקציות של אנאבוליזם מקבלים חיזור ודחיסה - בונים (דוחסים) שתי מולקולות ביחד.
גם בקטבוליזם וגם באנאבוליזם צריכים $\ce{ATP}$ - גם לשלבים ה־rate-limiting וגם לשלבים שדורשים אנרגיה. אבל הכוח המחזר שונה:
- בקטבוליזם: $\ce{FAD+}$ ו־$\ce{NAD+}$ הם מקבלי האלקטרונים
- באנאבוליזם: NADPH הוא הכוח המחזר
דוגמת הציפורים הנודדות
נדגים את החשיבות של סינתזת שומנים בעזרת ציפורים. יש ציפורים נודדות שעפות בשני אופנים - חלקן דואות (כמו עגורים וחסידות שאנחנו רואים באזור צפת בעת כתיבת שורות אלו), אבל ציפורים קטנות מאירופה בדרך כלל עפות ישירות מעל הים התיכון.
מה הציפורים האלה עושות לקראת סוף הקיץ? הן מגיעות לדרום אירופה, יושבות ואוכלות עוד ועוד, ואז מייצרות בבית החזה שלהן שכבת שומן אדירה שמשמשת אותן בכל הטיסה across the Mediterranean. הן מגיעות תשושות לחופי אפריקה על בסיס מאגרי השומן הללו. כאנקדוטה, זה מה שאולי ניתן להסביר ביחס לציפור השְׂלָו - ציפור שנודדת מאירופה לאפריקה, ומגיעה תשושה, וכך היה קל לבני ישראל לאסוף אותה ולאכול אותה במדבר.
למה שומן הוא צורת אגירה כל כך טובה?
שומנים הם צורת שימור האנרגיה העיקרית. אפשר לקבל מחמצון של שרשרת שומן אחת הרבה יותר מחמצון של מולקולת סוכר אחת!
פלמיטאט, חומצת השומן הבסיסית (16 פחמנים), מתפרקת לשמונה acetyl-CoA שנכנסים למעגל קרבס. לעומת זאת, מולקולת גלוקוז אחת מתפרקת רק לשני פירובטים, שהופכים לשני acetyl-CoA דרך פירובט דהידרוגנאז. מכאן, שבשומן נאגרת יחסית הרבה אנרגיה בצורה תמציתית.
מיקום הסינתזה של חומצות שומן
בניגוד ל־β-oxidation שמתרחשת במיטוכונדריה, סינתזת חומצות שומן נעשית בציטופלזמה. זה חשוב, כי acetyl-CoA צריך לצאת מהמיטוכונדריה ולהגיע לציטופלזמה כדי לבנות חומצות שומן.
מתי acetyl-CoA יוצא מהמיטוכונדריה לכיוון הציטופלזמה למטרת בנייה? כשהגוף במצב אנרגטי טוב (אם רוצים להשמין - אוכלים עוד ועוד, וכך בונים רקמת שומן מפורפלת).
רקמת השומן ותאי השומן
השומן שנבנה מאוכסן ברקמות שומן (adipose tissue), המכילות תאים שנקראים אדיפוציטים (adipocytes) - תאי שומן.
כשמסתכלים על תא שומן במיקרוסקופ אלקטרונים, רואים שהגרעין קטן מאוד ונדחק לשוליים. כמעט כל נפח התא תפוס על־ידי lipid droplet (טיפת שומן) גדולה. השומן נאגר בעיקר בצורת טריגליצרידים: מולקולת גליצרול שאליה קשורות שלוש חומצות שומן.
בקרה רציפרוקלית
בתא שומן, קיים כל הזמן מעין הלוך ושוב של סינתזה ופירוק חומצות שומן, תלוי במה שהגוף צריך לעשות מבחינה מטבולית. הבקרה היא תמיד רציפרוקלית:
- אם הגוף צריך חומצות שומן חופשיות (Free Fatty Acids) ל־β-oxidation ולאנרגיה ← הבקרה תעלה את הפירוק ותדכא את הסינתזה
- אם הגוף ברוויון אנרגטי ← נדכא את הפירוק ונעלה את הסינתזה
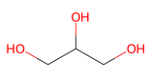
טריגליצרידים - המבנה
גליצרול הוא סוג של אלכוהול שמורכב משלד פחמני עם שלוש קבוצות $\ce{OH}$. קבוצות ה־$\ce{OH}$ הן הקבוצות הריאקטיביות שאליהן מתחברות שלוש חומצות שומן בקשר אסתרי.
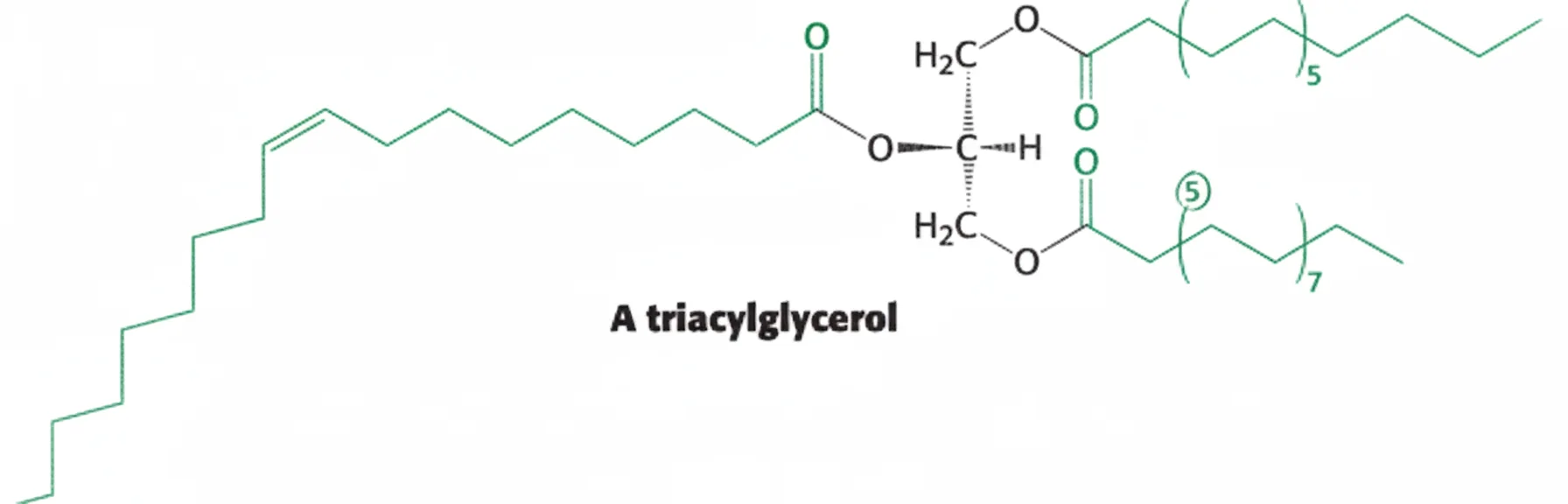
כשרואים בנוסחה סימון כמו $(\backslash)_7$, זה חזרה של שבע פעמים של שני פחמנים, כלומר 14 פחמנים. ביחד עם הפחמנים שמחוץ לסוגריים מקבלים 16 פחמנים - פלמיטאט.
פירוק טריגליצרידים על ידי ליפאז
ליפאז הוא אנזים שמופרש על ידי הלבלב ותפקידו לפרק את הטריגליצרידים ל־דיאצילגליצרידים ול־מונואצילגליצרידים. הוא לא מפרק ישירות לגליצרול ולחומצות שומן - תמיד מתקבל גליצרול עם חומצת שומן אחת או שתיים (בדרך כלל האמצעית נשארת).
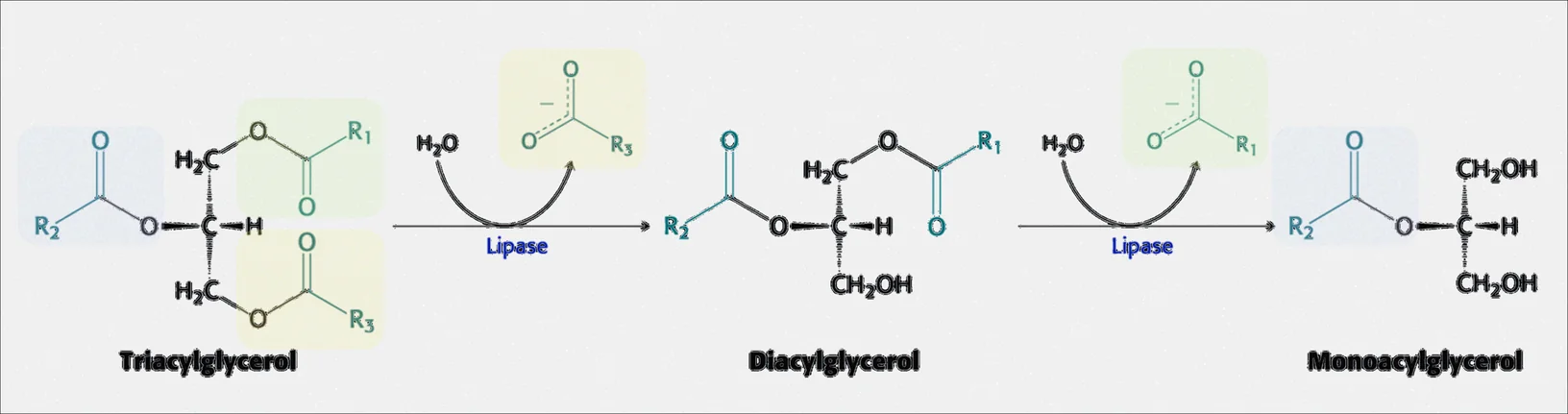
התהליך הוא הידרוליזה: $\ce{OH}$ הולך לפחמן של הגליצרול, ו־$\ce{H}$ נוסף מהמים הולך לחמצן, ומקבלים $\ce{COOH}$.
סינתזת חומצות שומן - סקירה כללית
מה אנחנו צריכים?
acetyl-CoA- חומר הבנייהNADPH- קופקטור, חומר מחזר- התוצר: פלמיטאט - חומצת שומן רוויה עם 16 פחמנים
סינתזת חומצות שומן היא איטרטיבי (חזרתית): הוספת שתי מולקולות פחמן מה־acetyl-CoA בכל מחזור, עד שמגיעים ל־16 פחמנים.
אבן הבניין - מלוניל קו־איי
אבן הבניין שמתווספת במחזורי הבנייה היא לא acetyl-CoA עצמו, אלא מולקולה אחרת בשם malonyl-CoA.
Malonyl-CoA היא מולקולה תלת־פחמנית: שני פחמנים הגיעו מה־acetyl-CoA, והפחמן השלישי הגיע מ־$\ce{CO2}$ (או ליתר דיוק מ־$\ce{HCO3^-}$, יון קרבונט). זו אבן הבניין לסינתזה של חומצות שומן.
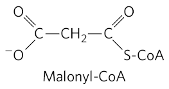
שלב ההתחייבות: יצירת מלוניל קו־איי
האנזים: acetyl-CoA Carboxylase (ACC)
האנזים שהופך acetyl-CoA ל־malonyl-CoA נקרא acetyl-CoA Carboxylase (בקיצור ACC). הוא עושה קרבוקסילציה - מוסיף קרבונט.
יצירת malonyl-CoA היא שלב ההתחייבות לסינתזת חומצות שומן. ל־malonyl-CoA אין תפקיד אחר - הוא משמש רק לבניית חומצות שומן - אז ברגע שהופכים acetyl-CoA למלוניל, מתחייבים לבניית חומצות שומן.
מבנה האנזים ומנגנון הפעולה
ל־ACC יש שלושה דומיינים:
- Biotin Carboxylase Domain
- Biotin Carrier Protein Domain
- Transcarboxylase Domain
יש קופקטור שנקרא ביוטין, שקשור לשייר ליזין של החלבון בדומיין אחד. לביוטין יש זרוע ארוכה של ארבעה פחמנים, והוא יודע לגשת לאתר הפעיל.
בשלב הראשון, יון קרבונט נכנס לאתר הפעיל של ה־Biotin Carboxylase Domain, ונוצר קשר קוולנטי בין הביוטין ליון הקרבונט. הריאקציה הזו דורשת $\ce{ATP}$ - יש כאן השקעת אנרגיה.
עכשיו הביוטין עושה 180 מעלות לאתר של הסינתזה (ה־Transcarboxylase Domain). שם מחכה לו acetyl-CoA באתר הפעיל. ה־$\ce{CO2}$ נמצא במעין holder מולקולרי שמחזיק אותו באוריינטציה הנכונה כדי שיידחס ל־acetyl-CoA, ומה שיוצא מהאתר הפעיל זה malonyl-CoA.
Fatty Acid Synthase - האנזים שבונה חומצות שומן
ההבדל בין חיידקים ליונקים
בחיידקים (וברוב הצמחים ובמיטוכונדריה של חולייתנים), סינתזת חומצות שומן נעשית באמצעות מערכת מפוצלת (dissociated system) של אנזימים נפרדים בשם FAS II: כל שלב בסינתזה (דחיסה, חיזור, דהידרציה וכו’) מזורז ע”י אנזים אחר, ולכן המערכת יכולה לייצר מגוון תוצרים.
ביונקים, מדובר באנזים אחד רב־תפקודי - FAS I: שבעה אתרים פעילים נמצאים בדומיינים נפרדים בתוך שרשרת פוליפפטידית אחת (multifunctional polypeptide chain). האנזים נמצא כ־הומודימר (שתי שרשראות פוליפפטידיות), כאשר שתי השרשראות מתפקדות באופן עצמאי אך כחלק מהדימר. FAS I נוטה לייצר תוצר יחיד עיקרי - פלמיטאט (C16:0).
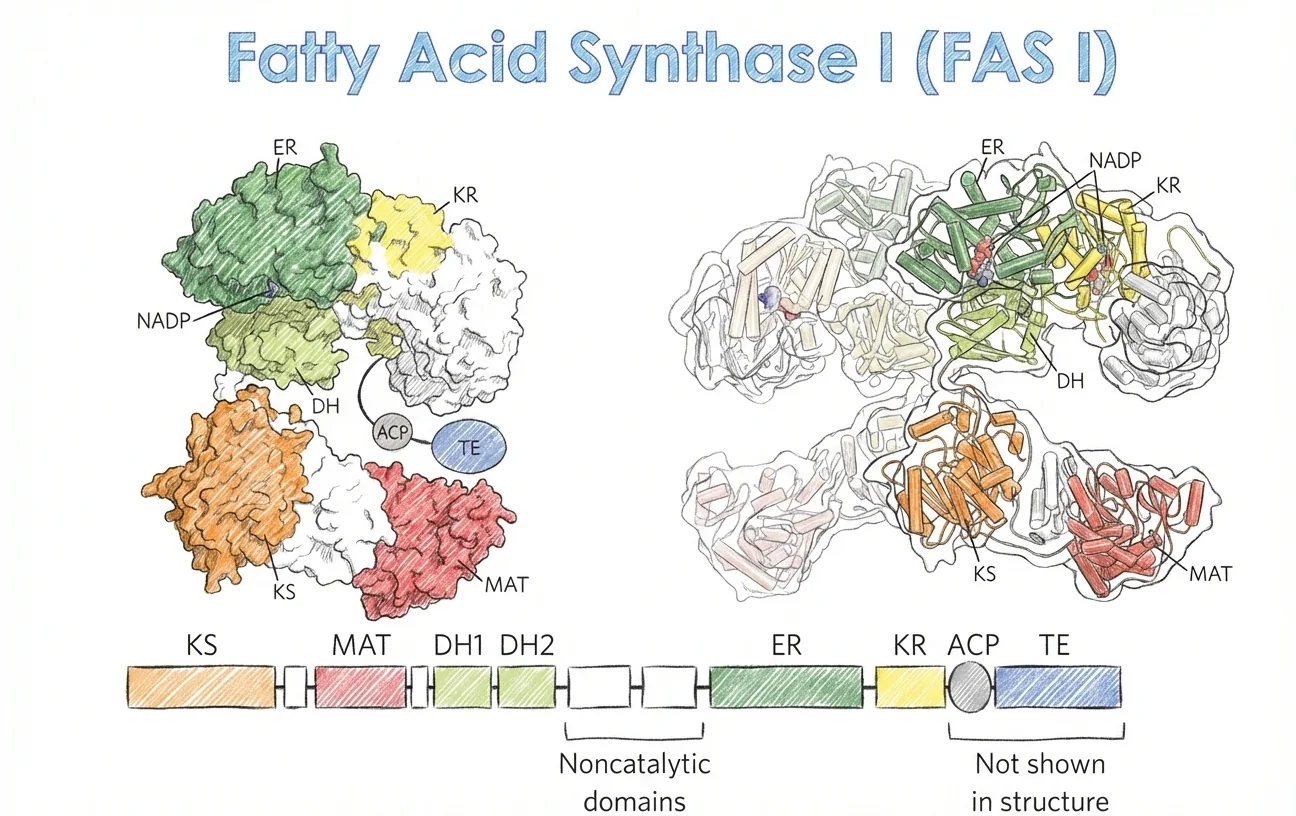
הדומיינים של Fatty Acid Synthase
לכל דומיין ראשי תיבות בהתאם לתפקידו:
- KS = β-Ketoacyl-ACP Synthase (דחיסה)
- DH = β-Hydroxyacyl-ACP Dehydratase (דהידרציה)
- ER = Enoyl-ACP Reductase (חיזור - כאן נמצא ה־NADPH)
- KR = β-Ketoacyl-ACP Reductase (חיזור)
- ACP = Acyl Carrier Protein
ACP - Acyl Carrier Protein
ACP הוא דומיין קטן (77 חומצות אמיניות) שמהווה חלק מה־Fatty Acid Synthase. הוא מאוד גמיש / פלקסיבילי, ולכן קשה לקבוע את המבנה שלו - הוא צריך לעבור מדומיין לדומיין.
על שייר סרין ב־ACP מחוברת קבוצה שנקראת פוספופנטתנאט - זו בדיוק אותה קבוצה שיש לקו־אנזים A. בקצה שלה יש טיול ($\ce{SH}$), ואליו נקשרת השרשרת הנבנית של חומצת השומן.
התפקיד של ACP הוא לקחת את השרשרת הנבנית ולהעביר אותה מדומיין לדומיין. כל הריאקציות מתבצעות על ה־ACP.
ארבעת שלבי הסינתזה
התהליך כולל ארבעה שלבים שחוזרים על עצמם שבע פעמים:
הכנת המערכת (Priming)
בהתחלה, קבוצת $\ce{Acetyl}$ (שהגיעה מ־acetyl-CoA) נקשרת לשייר ציסטאין שנמצא על דומיין אחר (ה־KS). ה־malonyl-CoA נקשר ל־ACP. עכשיו המערכת מוכנה.
שלב 1: דחיסה (Condensation)
קבוצת ה־Acetyl שהייתה על ה־KS נדחסת עם ה־malonyl-CoA, ויוצא $\ce{CO2}$. אחרי הדחיסה הראשונה, השרשרת הנבנית נמצאת על ה־ACP.
נוצרת קבוצת β-Keto - פחמן עם קטון על פחמן ה־β (כמו ב־β-Ketobutyrate ממעגל קרבס).
שלב 2: חיזור ראשון (Reduction)
יש לנו חיזור של הפחמן הזה על ידי מולקולה אחת של NADPH, ונוצר כוהל - הקשר הקרבונילי נעלם ובמקומו יש $\ce{OH}$.
זה נעשה בדומיין שנקרא β-Ketoacyl Reductase. רדוקטז = מחזר, והוא צריך קופקטור כדי לחזר - ה־NADPH צריך להיכנס לאתר הפעיל כדי לעשות את הפעולה.
שלב 3: דהידרציה (Dehydration)
התוצר עובר דהידרציה - מולקולת מים יוצאת. מימן אחד מהפחמן ה־α וה־$\ce{OH}$ מהפחמן ה־β יוצאים ביחד כמולקולת מים, ומקבלים קשר כפול.
שלב 4: חיזור שני (Reduction)
כדי לפתוח את הקשר הכפול ולקבל חומצת שומן רוויה, צריך לחזר אותו. מוסיפים לו אלקטרונים עם עוד NADPH, ומקבלים קשר בודד - $\ce{CH2}$, קשר רווי.
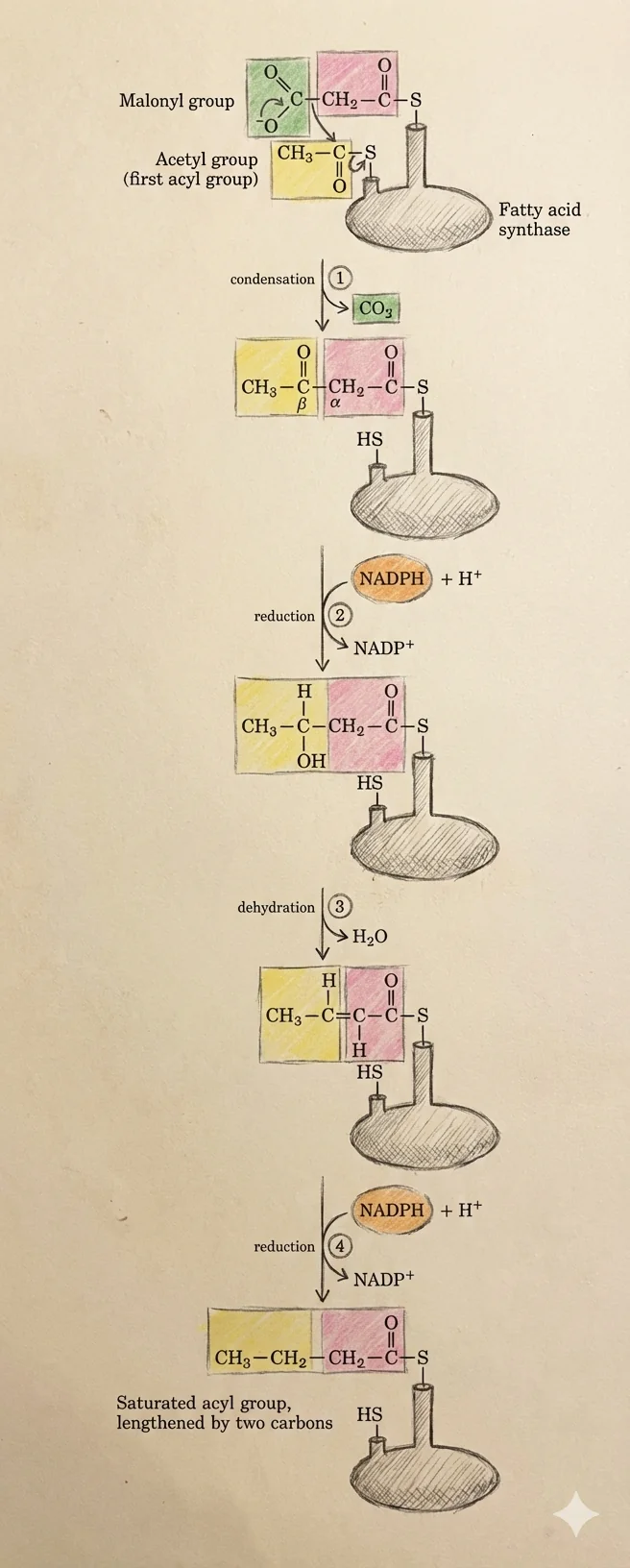
מה קורה אחרי ארבעת השלבים?
מה שנבנה על ה־ACP מועבר לציסטאין שב־KS, כדי לפנות מקום ל־malonyl-CoA חדש על ה־ACP. ואז מתחיל מחזור חדש.
הריאקציה מתרחשת שבע פעמים, ובסוף מגיעים לפלמיטאט עם 16 פחמנים. בשלב של 16 פחמנים, כל המבנה כבר לא יציב על Fatty Acid Synthase, ואז מולקולת מים גורמת להידרוליזה - מפרקת את הקשר עם ה־ACP ומשחררת חומצת שומן חופשית לסביבה.
חישוב הדרישות לסינתזת פלמיטאט
כדי לייצר חומצת שומן של 16 פחמנים, אנחנו צריכים:
acetyl-CoAאחד להתחלה (priming)- $\ce{7 Malonyl-CoA}$ (כל אחד מוסיף 2 פחמנים, סה”כ 14, ועוד 2 מההתחלה = 16)
ליצירת 7 מלוניל קו־איי (ב־ACC)
\[\ce{7 Acetyl-CoA + 7 CO2 + 7 ATP -> 7 Malonyl-CoA + 7 ADP + 7 P_i}\]לסינתזה עצמה
\[\ce{Acetyl-CoA + 7 Malonyl-CoA + 14 NADPH + 14 H+ -> Palmitate + 7 CO2 + 8 CoA + 14 NADP+ + 6 H2O}\](מולקולת מים אחת משמשת להידרוליזה של הקשר שמחבר את הפלמיטאט לאנזים)
זה תהליך שדורש השקעת אנרגיה משמעותית - לסנתז כל חומצת שומן.
מקורות ל־NADPH
מעגל הפנטוזות
דרך אחת לייצר NADPH היא מעגל הפנטוזות. התפקיד של מעגל הפנטוזות הוא לייצר סוכרים בעלי חמישה פחמנים (נוקלאוטידים - אבן בניין לנוקלאוטידים). בדרך, הוא מייצר NADPH שהוא כוח מחזר לתהליכים אנאבוליים.
Malic Enzyme
ביונקים יש דרך נוספת לייצר NADPH - אנזים שנקרא Malic Enzyme. הוא יודע לקחת מלאט ולהפוך אותו לפירובט:
\[\ce{Malate + NADP+ -> Pyruvate + CO2 + NADPH}\]רגע, אבל מלאט נמצא בדרך כלל במיטוכונדריה (חלק ממעגל קרבס), אז מה הוא עושה בציטוזול? כאן נכנס ה־Citrate Shuttle.
Citrate Shuttle - איך מוציאים acetyl-CoA מהמיטוכונדריה
הבעיה
acetyl-CoA נמצא בדרך כלל במיטוכונדריה - שם הוא נכנס למעגל קרבס, שם פירובט דהידרוגנאז יוצר אותו. אבל כדי לסנתז חומצות שומן, צריך להעביר אותו לציטוזול.
הפתרון
הופכים את acetyl-CoA לציטראט. יש לנו Citrate Synthase (אנזים שהוא חלק ממעגל קרבס) שלוקח אוקסלואצטט ו־acetyl-CoA ומחבר אותם לציטראט.
מתי ציטראט יעבור מהמיטוכונדריה החוצה? כשהגוף במצב של עודף אנרגיה: כשמעגל קרבס עובד בקיבולת מלאה (Full Capacity) וכבר לא יכול לעבוד מהר יותר, הציטראט העודף יוצא החוצה דרך Citrate Transporter.
בציטופלזמה
בציטופלזמה יש אנזים שנקרא Citrate Lyase שדורש $\ce{ATP}$ ו־$\ce{Coenzyme A}$. הוא מפרק את הציטראט ומחזיר לנו אוקסלואצטט וacetyl-CoA - בדיוק מה שהתחלנו איתו בהתחלה.
עכשיו ה־acetyl-CoA יכול ללכת לסינתזת חומצות שומן. אבל מה עושים עם האוקסלואצטט? צריך למחזר אותו חזרה כדי שהוא לא ייצטבר.
מחזור האוקסלואצטט
האוקסלואצטט עובר על ידי Malate Dehydrogenase ציטופלזמטי (יש לנו את השאטל הזה - Malate-Aspartate Shuttle) ונהפך למלאט.
המלאט יכול לעשות שני דברים:
- להיכנס חזרה למיטוכונדריה דרך ה־Malate-α-Ketoglutarate Transporter
- לעבור דרך Malic Enzyme - להפוך לפירובט, לשחרר $\ce{CO2}$, ולייצר NADPH שהולך לסינתזת חומצות שומן
הפירובט נכנס למיטוכונדריה דרך ה־Pyruvate Transporter.
היופי של המערכת
כל הדבר הזה הוא מעגל - צריך שלא יצטבר משהו כדי שהוא לא יתקע. אנחנו חייבים לקחת את האוקסלואצטט ולסלק אותו כדי להזרים את כל המערכת. זה הכל שיווי משקל.
הערות נוספות
על שריפת שומן בפעילות גופנית
אם אנחנו בפעילות אירובית עד 20 דקות - זה לא בהכרח מספיק בשביל להגיע לשריפת חומצות שומן. ב־20 הדקות הראשונות אנחנו מחלפים את הקריאטין פוספט, את הגלוקוז בדם ואת הגליקוגן. רק מ־20 דקות ומעלה מתחילים לשרוף שומן (לא בכמויות אדירות, אבל עדיין). אז כדי לייצר מטבוליזם של שומנים, חייבים לעשות פעילות אירובית לאורך זמן.
על דיאטת קטו
ככלל, המוח עובד על גלוקוז. אם עושים לו אדפטציה לגופי קטון (כמו ברעב או בדיאטת קטו), הוא יודע לעבוד גם על גופי קטון. אבל יש לזה חסרונות: עלייה של גופי קטון בדם מובילה לאסידוזיס (חומציות הדם), יש התפלמרות של גופי קטון, יש יצירת אצטואצטון ו־$\beta\text{-Ketobutyrate}$.
גופי קטון בדרך כלל טובים לזמן קצר, ולא בטוח שלעבוד עליהם כל הזמן זו דיאטה אידיאלית. מי שבכל זאת עושה דיאטת קטו צריך לנטר את רמות הטריגליצרידים ולעקוב אחרי תוצאות בדיקות הדם, כל הזמן. רוב מי שעושה קטו בגיל 40+ ככל הנראה צריך גם טיפול תרופתי כדי לשמור על טריגליצרידים נמוכים.
בקיצור: אין תוצאות בלי עבודה קשה, גם בביוכימיה.
שאלות מהתרגול
שאלה 1: rate-limiting step בסינתזה דה־נובו של חומצות שומן ביונקים
איזה משפט מתאר הכי נכון את הצעד קובע המהירות (rate-limiting step) בסינתזה דה־נובו של חומצות שומן ביונקים?
- דחיסת
acetyl-CoAעם malonyl-CoA ע״י fatty acid synthase ליצירת acetoacetyl-ACP, ללא צריכת אנרגיה. - קרבוקסילציה של
acetyl-CoAל־malonyl-CoA ע״יacetyl-CoA carboxylase, הדורשת ביוטין $\ce{ATP}$ ו־$\ce{CO2}$. - העברת קבוצת אצטיל מ־
acetyl-CoAלציטראט במיטוכונדריה לצורך יצירת malonyl-CoA בציטוזול. - הפחתה של acyl-ACP ל־saturated acyl-ACP בעזרת $\ce{NADH}$ בציטוזול
פתרון
התשובה הנכונה היא (2): הצעד קובע המהירות (rate-limiting step) בסינתזה דה־נובו של חומצות שומן ביונקים הוא הקרבוקסילציה של acetyl-CoA ל־malonyl-CoA על ידי האנזים acetyl-CoA carboxylase (ACC). ריאקציה זו דורשת ביוטין, $\ce{ATP}$ ו־$\ce{CO2}$, והיא מהווה את שלב ההתחייבות לתהליך סינתזת חומצות השומן.
למה התשובות האחרות לא נכונות?
- 1. הקונדנסציה ע״י fatty acid synthase חשובה, אך אינה הצעד קובע המהירות; היא מונעת ע״י דה־קרבוקסילציה של malonyl-ACP ולא דורשת ATP ישירות.
- 3. יצירת/ייצוא ציטראט (citrate shuttle) הוא שלב אספקת אצטיל־CoA לציטוזול, אך אינו הצעד קובע המהירות של הסינתזה עצמה.
- 4. שלבי ההפחתה ב־fatty acid synthase משתמשים ב־NADPH (לא NADH) ואינם ה־rate-limiting step.
שאלה 2: תפקיד ה־ACP ב־Fatty Acid Synthase I
מהו התפקיד העיקרי של acyl carrier protein (ACP) בקומפלקס fatty acid synthase I (FAS I) ביונקים?
- קושר את ה־
acetyl-CoAלאתר הפעיל של β-ketoacyl-synthase לצורך הדחיסה הראשונית בלבד. - משמש כ״זרוע נדנדה״ ניידת הנושאת את acyl/malonyl-CoA כקבוצות תיו־אסטר ומעבירה אותם בין הדומיינים השונים של האנזים.
- מפרק את חומצת השומן המוגמרת מהקומפלקס בעזרת פעילות thioesterase.
- מספק NADPH לתגובות ההפחתה דרך קבוצת פוספו־פנטתאין שלו.
פתרון
התשובה הנכונה היא תשובה 2. ה־ACP (Acyl Carrier Protein) משמש כ״זרוע נדנדה״ ניידת בתוך קומפלקס ה־Fatty Acid Synthase I (FAS I) ביונקים. הוא נושא את קבוצות ה־acyl/malonyl-CoA כקבוצות תיו־אסטר ומעביר אותן בין הדומיינים השונים של האנזים במהלך תהליך סינתזת חומצות השומן.
שאלה 3: בקרה של acetyl-CoA carboxylase (ACC) במטבוליזם חומצות שומן בכבד
לאחר ארוחה עתירת פחמימות, מהו השילוב הכי נכון של שינויים ברמת acetyl CoA carboxylase (ACC) ובמטבוליזם חומצות שומן בכבד?
- רמות ציטראט נמוכות, ACC מזורחן (לא פעיל), רמות malonyl CoA נמוכות, ו־β-אוקסידציה מוגברת.
- רמות ציטראט גבוהות, אינסולין גורם לדה־פוספורילציה ואקטיבציה של ACC, רמות malonyl CoA עולות ומדכאות כניסת acyl-CoA למיטוכונדריה, כך ש־β-אוקסידציה מעוכבת וסינתזת חומצות שומן מוגברת.
- גלוקגון מפעיל phosphatase שמסיר פוספט מ־ACC מה שמגביר סינתזת חומצות שומן ומדכא גלוקונאוגנזה.
- אינסולין מפעיל AMP-activated protein kinase (AMPK) שמזרחן את ACC ומגביר בו־זמנית גם סינתזה וגם פירוק של חומצות שומן.
פתרון
התשובה הנכונה היא (2). לאחר ארוחה עתירת פחמימות, רמות הציטראט עולות, מה שמעיד על מצב אנרגטי טוב בתא. אינסולין גורם לדה־פוספורילציה של acetyl-CoA carboxylase (ACC), מה שמפעיל אותו. כתוצאה מכך, רמות ה־malonyl-CoA עולות, מה שמדכא את כניסת acyl-CoA למיטוכונדריה ומעכב את β-אוקסידציה. במקביל, סינתזת חומצות השומן מוגברת.