משוואת מיכאליס-מנטן - הגזירה המלאה
נזכיר שהנוסחה לראקציה אנזימטית פשוטה היא:
\[\ce{E + S <=>[k_1][k_{-1}] ES ->[k_2] E + P}\]הנחות היסוד
בפיתוח משוואת מיכאליס-מנטן, אנחנו מבססים את הניתוח על מספר הנחות יסוד קריטיות:
הנחה ראשונה: הזנחת הריאקציה ההפוכה
\[k_2 \gg k_{-2}\]כלומר, קצב יצירת התוצר גדול בהרבה מקצב הריאקציה ההפוכה (מתוצר לקומפלקס $\ce{ES}$). ההנחה הזאת מוצדקת כשאנחנו מודדים את מהירות הריאקציה ההתחלתית - כשריכוז התוצר הוא כמעט אפסי.
הנחה שנייה: עודף סובסטרט
\[\left[ \text{S} \right] \gg \left[ \text{E} \right]_{total}\]ריכוז הסובסטרט תמיד בעודף גדול מאוד ביחס לריכוז האנזים הכולל. משמעות ההנחה הזאת היא שהסובסטרט הוא לא הגורם המגביל בריאקציה.
הנחה שלישית: המצב העמיד (Steady State)
\[\frac{d\left[ \text{ES} \right]}{dt} = 0\]ריכוז הקומפלקס $\ce{ES}$ נשאר קבוע לאורך המדידה. זאת הנחת המצב העמיד - היא שונה משיווי משקל, כי היא מתייחסת לרכיב בודד במערכת ולא למערכת כולה.
גזירת המשוואה
במצב העמיד, קצב היצירה של $\ce{ES}$ שווה לקצב הפירוק שלו:
יצירת $\ce{ES}$:
\[\text{Formation rate} = k_1\left[ \text{E} \right]\left[ \text{S} \right]\]במבחן של הטכניון ראיתי את הביטוי הבא לקצב יצירת הקומפלקס $\ce{ES}$:
\[\text{Formation rate} = k_1\left( \left[ \text{E} \right]_{total} - \left[ \text{ES} \right] \right)\left[ \text{S} \right]\]להבנתי, היא מתקבלת מכך שניתן לבטא את ריכוז האנזים החופשי כ־$\left[ \text{E} \right] = \left[ \text{E} \right]_{total} - \left[ \text{ES} \right]$ (כי האנזים יכול להיות רק חופשי או בקומפלקס).
פירוק $\ce{ES}$:
\[\text{Breakdown rate} = [k_{-1} + k_2](ES)\]במצב העמיד קצב היצירה שווה לקצב הפירוק (ראו את ההנחה השלישית לעיל), מכאן:
\[k_1\left[ \text{E} \right]\left[ \text{S} \right] = [k_{-1} + k_2](ES)\]נגדיר את קבוע מיכאליס-מנטן:
\[K_{\text{m}} = \frac{k_{-1} + k_2}{k_1}\]זהו יחס בין קבועי הקצב של פירוק $\ce{ES}$ (לשני הכיוונים) לקבוע הקצב של יצירת $\ce{ES}$.
מכאן:
\[K_{\text{m}} = \frac{[\text{E}][\text{S}]}{[\text{ES}]}\]כאשר ריכוז האנזים הכולל:
\[[\text{E}]_{total} = [\text{E}] + [\text{ES}]\]נציב ונקבל:
\[[\text{ES}] = \frac{[\text{E}]_{total} \cdot [\text{S}]}{K_{\text{m}} + [\text{S}]}\]המהירות ההתחלתית:
\[V_0 = k_2\left[ \text{ES} \right] = \frac{k_2[\text{E}]_{total} \cdot [\text{S}]}{K_{\text{m}} + [\text{S}]}\]כאשר כל האנזים רווי בסובסטרט:
\[V_{\text{max}} = k_2\left[ \text{E} \right]_{total}\]לכן משוואת מיכאליס-מנטן:
\[\boxed{V_0 = \frac{V_{\text{max}} \cdot [\text{S}]}{K_{\text{m}} + [\text{S}]}}\]המשמעות של KM
ההגדרה המתמטית: כאשר $[\text{S}] = K_{\text{m}}$:
\[V_0 = \frac{V_{\text{max}} \cdot K_{\text{m}}}{K_{\text{m}} + K_{\text{m}}} = \frac{V_{\text{max}}}{2}\]לכן: $K_{\text{m}}$ הוא ריכוז הסובסטרט שבו מהירות הריאקציה היא מחצית המהירות המקסימלית.
מקרה פרטי - אפיניות: כאשר $k_2 \ll k_{-1}$ (השלב הקובע-קצב הוא יצירת התוצר):
\[K_{\text{m}} \approx \frac{k_{-1}}{k_1} = K_d\]במקרה זה בלבד, $K_{\text{m}}$ משקף את האפיניות בין האנזים לסובסטרט.
פרמטרים קינטיים חשובים
מספר התחלופה (Turnover Number)
\[K_{\text{cat}} = \frac{V_{\text{max}}}{\left[ \text{E} \right]_{total}}\]מספר מולקולות הסובסטרט שאנזים בודד הופך לתוצר ביחידת זמן (בתנאי רוויה).
דוגמאות:
- קרבוניק אנהידרז: $K_{\text{cat}} = 600,000 \, \mathrm{s}^{-1}$
- ליזוזים: $K_{\text{cat}} = 0.5 \, \mathrm{s}^{-1}$
השפעת תנאים סביבתיים
pH: כל אנזים פועל באופן אופטימלי ב־pH מסוים:
- פפסין (קיבה): pH אופטימלי = 2
- גלוקוז-6-פוספטאז (רקמות): pH אופטימלי = 6-7
טמפרטורה: השפעה דרמטית על קצב הריאקציה - אנזים שחותך חלבון ב־3 שעות ב־4°C יבצע את אותה ריאקציה ב־10 דקות ב־25°C.
גרף ליינוויבר-בורק
כדי לקבוע במדויק את $V_{\text{max}}$ ו־$K_{\text{m}}$ (שלעיתים קשה להגיע אליהם אמפירית), משתמשים בלינאריזציה:
\[\boxed{\frac{1}{V_0} = \frac{K_{\text{m}}}{V_{\text{max}}} \cdot \frac{1}{\left[ \text{S} \right]} + \frac{1}{V_{\text{max}}}}\]זאת משוואת קו ישר מהצורה $y = ax + b$ כאשר:
- ציר Y: $1/V_0$
- ציר X: $1/\left[ \text{S} \right]$
- שיפוע: $K_{\text{m}}/V_{\text{max}}$
- חיתוך עם ציר Y: $1/V_{\text{max}}$
- חיתוך עם ציר X: $-1/K_{\text{m}}$
איזוזימים - משמעות פיזיולוגית
הגדרה
איזוזימים הם אנזימים שונים שמקטלזים את אותה ריאקציה אך נבדלים ב:
- רצף חומצות אמינו
- ערכי $K_{\text{m}}$
- ערכי $V_{\text{max}}$
דוגמה: מטבוליזם של אלכוהול
אצטאלדהיד דהידרוגנז (ALDH):
- איזוזים מיטוכונדריאלי: $K_{\text{m}}$ נמוך - פועל ביעילות בריכוזי אצטאלדהיד נמוכים
- איזוזים ציטופלזמטי: $K_{\text{m}}$ גבוה - נכנס לפעולה רק בריכוזים גבוהים
מוטציה ב־ALDH המיטוכונדריאלי גורמת לתסמונת רגישות לאלכוהול (הסמקה, בחילה, טכיקרדיה).
דוגמה: הקסוקינאז מול גלוקוקינאז
שניהם מזרחנים גלוקוז ל־גלוקוז-6-פוספט:
הקסוקינאז (רוב הרקמות):
- $K_{\text{m}} = 0.1 \, \text{mM}$
- פועל ביעילות בריכוזי גלוקוז נמוכים
- מבטיח אספקת גלוקוז לתאים
גלוקוקינאז (כבד):
- $K_{\text{m}} = 10 \, \text{mM}$ (גבוה פי 100!)
- פועל רק בריכוזי גלוקוז גבוהים
- מאחסן עודפי גלוקוז כגליקוגן
המשמעות: הכבד “לוקח” גלוקוז רק אחרי שהרקמות החיוניות קיבלו את צרכיהן.
עיכוב אנזימים
עיכוב בלתי הפיך
מעכבים הנקשרים קוולנטית לאנזים ומוציאים אותו מכלל פעולה לצמיתות.
דוגמאות:
DIPF (גז עצבים):
- נקשר קוולנטית לסרין באתר הפעיל של אצטילכולין אסטראז
- משבש העברה עצבית בסינפסות
פניצילין:
- נקשר קוולנטית ל־Glycopeptide Transpeptidase
- מונע יצירת קשרי צילוב בדופן התא החיידקי
- גורם למות החיידק
עיכוב הפיך
קיימים מספר סוגים של עיכוב הפיך, כאשר שני הסוגים העיקריים הם עיכוב תחרותי ועיכוב אל־תחרותי (Uncompetitive).
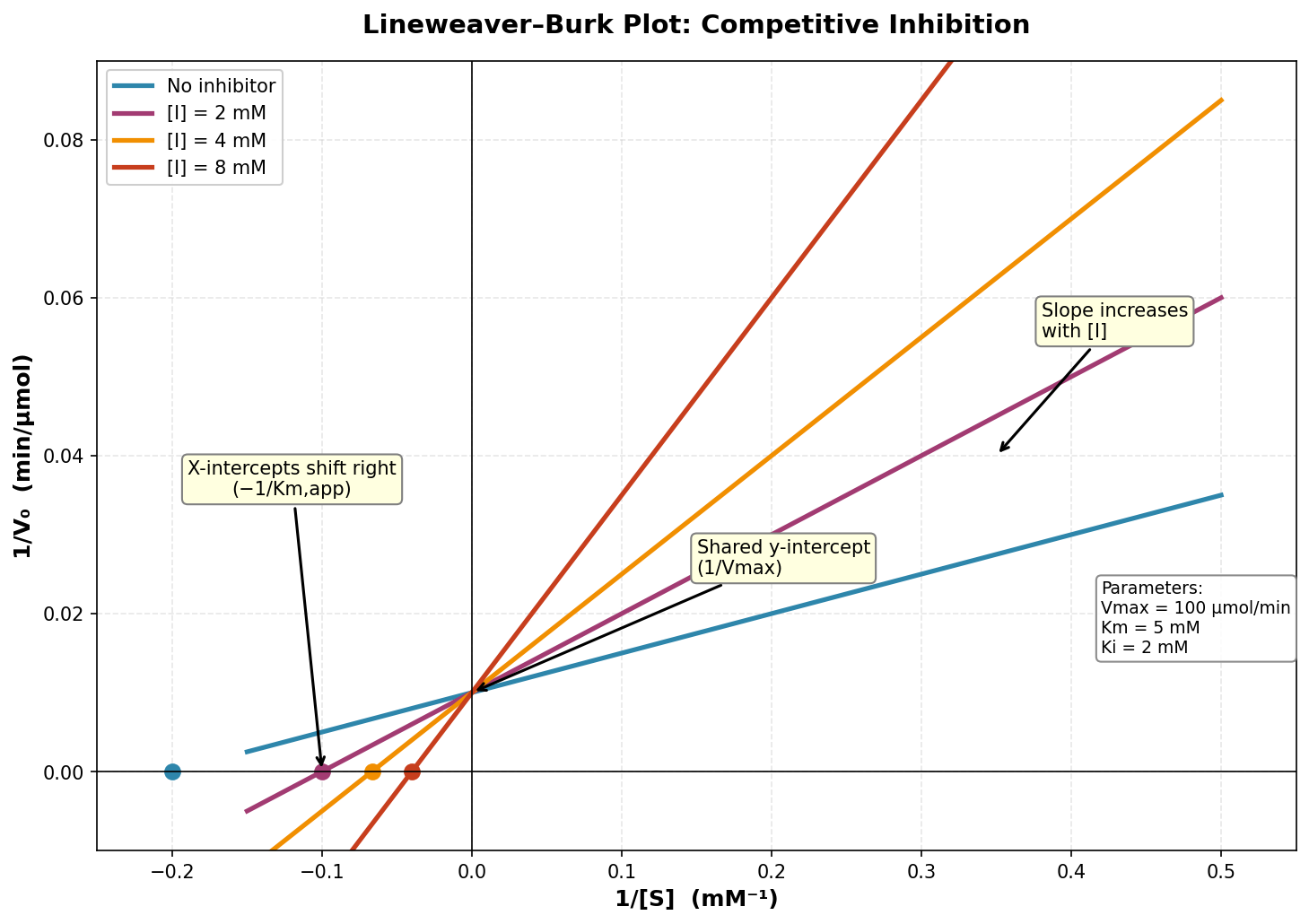
עיכוב תחרותי (Competitive)
המעכב מתחרה עם הסובסטרט על האתר הפעיל.
מאפיינים קינטיים:
- $V_{\text{max}}$ לא משתנה (ניתן להתגבר על העיכוב בריכוזי סובסטרט גבוהים)
- $K_{\text{m}}^{apparent} = K_{\text{m}} \cdot \alpha$ כאשר $\alpha = 1 + \frac{[I]}{K_i}$
בגרף ליינוויבר-בורק:
- כל הקווים נחתכים על ציר Y (אותו $1/V_{\text{max}}$)
- שיפוע עולה עם ריכוז המעכב - משקף את העלייה ב־$K_{\text{m}}$ שהמשמעות שלה היא לכאורה ירידה באפיניות בין האנזים לסובסטרט. בפועל, המעכב תופס את האתר הפעיל ומונע מהסובסטרט להיקשר, ולכן נדרש ריכוז סובסטרט גבוה יותר כדי להגיע לאותה מהירות (כזכור, $K_{\text{m}}$ הוא הריכוז שבו המהירות היא חצי מ־$V_{\text{max}}$).
דוגמה: מתוטרקסט - מעכב של דיהידרופולאט רדוקטאז, משמש בכימותרפיה.
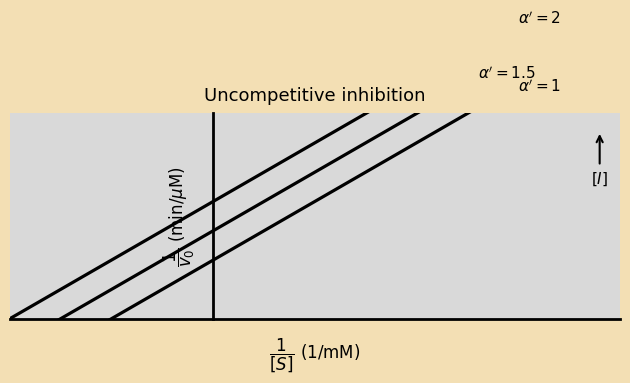
עיכוב אל־תחרותי (Uncompetitive)
המעכב נקשר רק לקומפלקס ES, לא לאנזים החופשי.
מאפיינים קינטיים:
- גם $V_{\text{max}}$ וגם $K_{\text{m}}$ יורדים באותו יחס
- היחס $V_{\text{max}}/K_{\text{m}}$ נשאר קבוע
בגרף ליינוויבר-בורק:
- קווים מקבילים (אותו שיפוע)
- חיתוך שונה עם שני הצירים
הקווים בנוכחות מעכב אל־תחרותי תמיד יופיעו בגרף ליינוויבר-בורק מעל הקו של האנזים ללא המעכב, כתוצאה מהירידה גם ב־$V_{\text{max}}$ וגם ב־$K_{\text{m}}$ (נקודות החיתוך זזות שמאלה על ציר X וגבוהות יותר על ציר Y).
בגרף רגיל ($V_0$ מול $[\text{S}]$), הקווים בנוכחות מעכב אל־תחרותי יופיעו מתחת לקו של האנזים ללא המעכב, כי המהירות בכל ריכוז סובסטרט תהיה נמוכה יותר. האפיניות דווקא עולה (כי $K_{\text{m}}$ יורד) אבל המהירות בכל ריכוז סובסטרט נמוכה יותר בגלל הירידה ב־$V_{\text{max}}$.
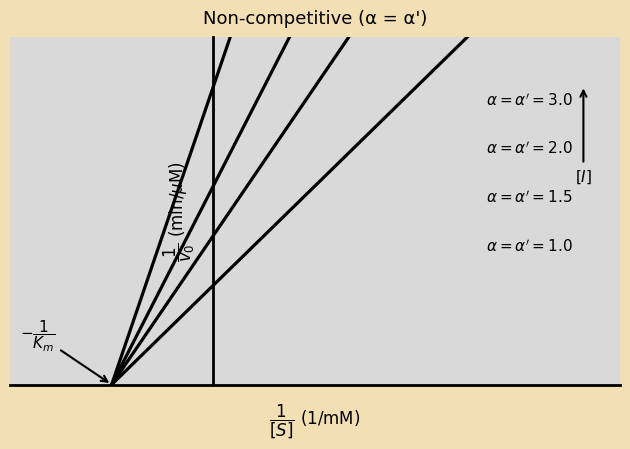
עיכוב לא תחרותי (Non-competitive)
לא הרחבנו על זה בשיעור למיטב זכרוני אבל כן הופיע בתרגול.
מדובר במקרה פרטי של Mixed Inhibition (מעכב שיכול להיקשר גם לאנזים החופשי E וגם לקומפלקס ES), שבו הנטייה של המעכב להיקשר לאנזים החופשי שווה לנטייה שלו להיקשר לקומפלקס ES. במצב כזה:
- $V_{\text{max}}$ יורד
- $K_{\text{m}}$ נשאר קבוע
המשוואה הכללית לעיכוב
עבור עיכוב תחרותי:
\[V_0 = \frac{V_{\text{max}} \cdot \left[ \text{S} \right]}{\alpha K_{\text{m}} + \left[ \text{S} \right]}\]כאשר $\alpha$ הוא מקדם העיכוב התלוי בריכוז המעכב ובקבוע העיכוב $K_i$.
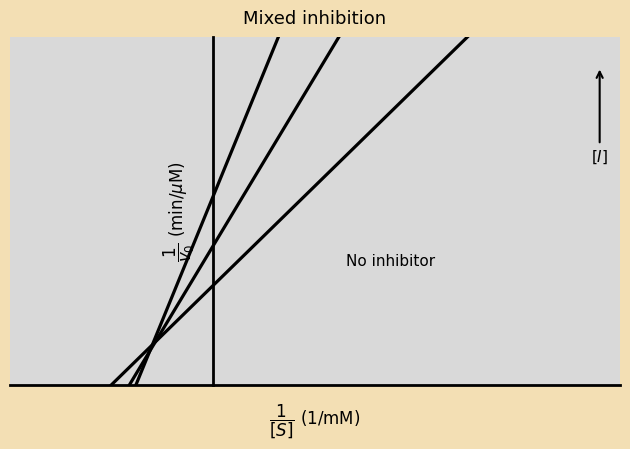
כאשר בעיכוב מסוג Mixed הנטייה של המעכב להיקשר ל־E גבוהה מזו של הקישור ל־ES (כלומר $\alpha > \alpha’$),
- ערך $K_{\text{m}}$ עולה (ירידה באפיניות),
- השיפוע עולה (כי $\alpha K_m$ גדל),
- החיתוך עם ציר X זז ימינה (כלומר ערך $-1/K_m$ מתקרב לאפס).
חישוב דוגמה
נתון: ריאקציה אנזימטית שמתנהגת לפי מיכאליס מנטן עם הנתונים הבאים:
$\left[ \text{S} \right] \, (\mathrm{M})$ $V \, (\mathrm{M/s})$ $1\times10^{-4}$ $1\times10^{-4}$ $4\times10^{-3}$ $6\times10^{-4}$ $2\times10^{-3}$ $6\times10^{-4}$ מה יהיו $K_{\text{m}}$ ו־$V$ אם ריכוז הסובסטרט יהיה $5\times 10^{-4} \, \mathrm{M}$?
אפשר ללמוד מהטבלה על המהירות המקסימלית (הוספת ריכוז לא מעלה את המהירות מעבר לה):
\[V_{\text{max}} = 6\times10^{-4} \, (\mathrm{M/s})\]$K_{\text{m}}$ הוא הריכוז שבו המהירות היא בדיוק חצי. נציב במשוואה כדי למצוא אותו:
\[\frac{6\times10^{-4}}{2} = \frac{ 6\times10^{-4} \times 5\times 10^{-4}}{K_{\text{m}} + 5\times 10^{-4}}\] \[K_{\text{m}} + 5\times 10^{-4} = 10 \times 10^{-4}\] \[\boxed{K_{\text{m}} = 5\times 10^{-4}}\]כעת נציב שוב בנוסחה כדי למצוא את המהירות בנתונים ששאלו עליהם:
\[V = \frac{ 6\times10^{-4} \times 5\times 10^{-4}}{ 5\times 10^{-4} + 5\times 10^{-4}} = \boxed{3\times10^{-4}}\]עקרונית היה ניתן להגיע למהירות גם בדרך אחרת - בריכוז של $k_{\text{m}}$ המהירות היא חצי מהמהירות המקסימלית.
סיכום
קינטיקה אנזימטית מספקת כלים כמותיים להבנת פעילות אנזימים ולפיתוח תרופות. משוואת מיכאליס-מנטן ונגזרותיה מאפשרות:
- אפיון כמותי של אנזימים ($K_{\text{m}}$, $V_{\text{max}}$, $K_{\text{cat}}$)
- הבנת מנגנוני עיכוב
- תכנון רציונלי של תרופות
- הבנת המשמעות הפיזיולוגית של איזוזימים
העקרונות הללו הם הבסיס להבנת רגולציה מטבולית ולפיתוח טיפולים תרופתיים רבים.
שאלות ממוצאות לתרגול
שאלה 1: המשמעות של Km
מהי המשמעות המתמטית של $K_m$ במשוואת מיכאליס-מנטן, ומתי הוא משקף את האפיניות בין האנזים לסובסטרט?
- $K_m$ הוא תמיד שווה לקבוע הדיסוציאציה ($K_d$) ומשקף את האפיניות בכל מקרה.
- $K_m$ הוא ריכוז הסובסטרט שבו המהירות מקסימלית, ומשקף אפיניות רק כש־$k_2 \gg k_{-1}$.
- $K_m$ הוא ריכוז הסובסטרט שבו המהירות היא מחצית המהירות המקסימלית, ומשקף אפיניות רק כש־$k_2 \ll k_{-1}$.
- $K_m$ הוא ריכוז האנזים שבו המהירות היא מחצית המהירות המקסימלית.
פתרון
התשובה הנכונה היא (3).
ההגדרה המתמטית של $K_m$:
כאשר $[\text{S}] = K_m$:
\[V_0 = \frac{V_{max} \cdot K_m}{K_m + K_m} = \frac{V_{max}}{2}\]לכן: $K_m$ הוא ריכוז הסובסטרט שבו מהירות הריאקציה היא מחצית המהירות המקסימלית.
מתי $K_m$ משקף אפיניות?
\[K_m = \frac{k_{-1} + k_2}{k_1}\]רק כאשר $k_2 \ll k_{-1}$ (השלב הקובע קצב הוא יצירת התוצר):
\[K_m \approx \frac{k_{-1}}{k_1} = K_d\]במקרה זה בלבד, $K_m$ משקף את האפיניות בין האנזים לסובסטרט.
חשוב לזכור:
- $K_m$ נמוך = האנזים מגיע לחצי מהירות מקסימלית בריכוזי סובסטרט נמוכים
- $K_m$ גבוה = צריך ריכוז סובסטרט גבוה כדי להגיע לחצי מהירות מקסימלית
שאלה 2: איזוזימים - הקסוקינאז מול גלוקוקינאז
הקסוקינאז וגלוקוקינאז שניהם מזרחנים גלוקוז לגלוקוז-6-פוספט. ל־הקסוקינאז $K_m = 0.1 \text{ mM}$ ולגלוקוקינאז $K_m = 10 \text{ mM}$. מה המשמעות הפיזיולוגית של ההבדל?
- גלוקוקינאז יעיל יותר ולכן נמצא ברקמות החיוניות.
- הכבד (עם גלוקוקינאז) “לוקח” גלוקוז רק אחרי שהרקמות החיוניות קיבלו את צרכיהן.
- הקסוקינאז פועל רק בריכוזי גלוקוז גבוהים.
- אין משמעות פיזיולוגית - זו רק וריאציה גנטית.
פתרון
התשובה הנכונה היא (2).
השוואה בין האיזוזימים:
| פרמטר | הקסוקינאז | גלוקוקינאז |
|---|---|---|
| מיקום | רוב הרקמות | כבד |
| $K_m$ | $0.1 \, \mathrm{mM}$ (נמוך) | $10 \, \mathrm{mM}$ (גבוה פי 100!) |
| מתי פועל | בריכוזי גלוקוז נמוכים | רק בריכוזי גלוקוז גבוהים |
| תפקיד | מבטיח אספקת גלוקוז לתאים | מאחסן עודפי גלוקוז כגליקוגן |
המשמעות הפיזיולוגית:
- כשרמת הגלוקוז בדם נמוכה ← הקסוקינאז ברקמות פועל ביעילות
- גלוקוקינאז בכבד לא פעיל (צריך ריכוז גבוה בהרבה)
- רק כשיש עודף גלוקוז בדם ← גלוקוקינאז נכנס לפעולה ומאחסן בכבד
התוצאה: הכבד “מחכה בתור” - לוקח גלוקוז רק אחרי שהרקמות החיוניות (מוח, שרירים) קיבלו את צרכיהן.
שאלה 3: גרף ליינוויבר-בורק ועיכוב תחרותי
בניסוי מעבדה נמדדה פעילות אנזימטית עם ובלי מעכב. בגרף ליינוויבר-בורק, כל הקווים נחתכים באותה נקודה על ציר Y. מה סוג העיכוב?
- עיכוב אל־תחרותי (Uncompetitive) - המעכב נקשר רק לקומפלקס ES.
- עיכוב תחרותי (Competitive) - המעכב מתחרה עם הסובסטרט על האתר הפעיל.
- עיכוב בלתי הפיך - המעכב נקשר קוולנטית לאנזים.
- עיכוב מעורב - המעכב נקשר גם לאנזים וגם לקומפלקס ES.
פתרון
התשובה הנכונה היא (2) - עיכוב תחרותי.
עיכוב תחרותי (Competitive Inhibition):
המעכב מתחרה עם הסובסטרט על האתר הפעיל.
מאפיינים קינטיים:
- $V_{max}$ לא משתנה (ניתן להתגבר על העיכוב בריכוזי סובסטרט גבוהים)
- $K_m^{apparent}$ עולה (צריך יותר סובסטרט כדי להגיע לחצי מהירות)
בגרף ליינוויבר-בורק:
- כל הקווים נחתכים על ציר Y (אותו $1/V_{max}$)
- שיפוע עולה עם ריכוז המעכב
- חיתוך עם ציר X משתנה ($-1/K_m$ הופך לפחות שלילי)
לעומת עיכוב אל־תחרותי:
- קווים מקבילים (אותו שיפוע)
- גם $V_{max}$ וגם $K_m$ יורדים
דוגמה קלינית: מתוטרקסט - מעכב תחרותי של דיהידרופולאט רדוקטאז, משמש בכימותרפיה.
שאלה 4: עיכוב בלתי הפיך
פניצילין הוא אנטיביוטיקה שפועלת כמעכב בלתי הפיך. מה מנגנון הפעולה שלו?
- מתחרה עם הסובסטרט על האתר הפעיל של אנזימי החיידק.
- נקשר קוולנטית ל־Glycopeptide Transpeptidase ומונע יצירת קשרי צילוב בדופן התא החיידקי.
- נקשר לקומפלקס ES בלבד ומונע שחרור התוצר.
- משנה את ה־pH באתר הפעיל של האנזים.
פתרון
התשובה הנכונה היא (2).
עיכוב בלתי הפיך:
מעכבים הנקשרים קוולנטית לאנזים ומוציאים אותו מכלל פעולה לצמיתות.
מנגנון הפעולה של פניצילין:
- נקשר קוולנטית ל־Glycopeptide Transpeptidase
- מונע יצירת קשרי צילוב (Cross-links) בדופן התא החיידקי
- דופן התא נחלשת ← החיידק מת
דוגמה נוספת - DIPF (גז עצבים):
- נקשר קוולנטית לסרין באתר הפעיל של אצטילכולין אסטראז
- משבש העברה עצבית בסינפסות
- גורם לשיתוק ומוות
ההבדל מעיכוב הפיך:
- עיכוב הפיך: המעכב יכול להתנתק מהאנזים
- עיכוב בלתי הפיך: קשר קוולנטי = האנזים “מת” לצמיתות
שאלה 5: הנחות מיכאליס-מנטן
איזו מההנחות הבאות היא הנחת “המצב העמיד” (Steady State) בגזירת משוואת מיכאליס-מנטן?
- ריכוז הסובסטרט שווה לריכוז האנזים.
- ריכוז הקומפלקס ES נשאר קבוע לאורך המדידה: $\frac{d[ES]}{dt} = 0$
- הריאקציה נמצאת בשיווי משקל תרמודינמי מלא.
- קצב יצירת התוצר שווה לאפס.
פתרון
התשובה הנכונה היא (2).
שלוש הנחות היסוד של מיכאליס-מנטן:
| הנחה | משמעות |
|---|---|
| 1. הזנחת הריאקציה ההפוכה | $k_2 \gg k_{-2}$ - מודדים מהירות התחלתית כשאין עדיין תוצר |
| 2. עודף סובסטרט | $[S] \gg [E]_{total}$ - הסובסטרט לא הגורם המגביל |
| 3. המצב העמיד | $\frac{d[ES]}{dt} = 0$ - ריכוז הקומפלקס ES קבוע |
מה זה המצב העמיד?
- קצב יצירת ES = קצב פירוק ES
- ריכוז ES קבוע (לא משתנה עם הזמן)
- זה לא שיווי משקל - הריאקציה ממשיכה להתקדם!
ההבדל משיווי משקל:
- שיווי משקל = כל המערכת לא משתנה
- מצב עמיד = רכיב בודד (ES) קבוע, אבל הריאקציה ממשיכה
מהנחת המצב העמיד נגזרת המשוואה:
\[k_1[E][S] = (k_{-1} + k_2)[ES]\]ומכאן מגיעים למשוואת מיכאליס-מנטן.