עיבוד RNA בזמן שעתוק
בשיעורים הקודמים הוגדרו שלושה פולימראזות עיקריים בגרעין האאוקריוטי: RNA Polymerase I, RNA Polymerase II ו־RNA Polymerase III. בשיעור הנוכחי נתמקד במה שקורה ל־RNA לאחר שהוא מתחיל להיווצר, ובפרט בעיבודו עד לקבלת המבנה הבוגר והפעיל שלו.
כל סוג RNA עובר עיבוד אחר:
| סוג RNA | פולימראז עיקרי | עיבוד עיקרי |
|---|---|---|
| mRNA | Pol II | 5′ capping, polyadenylation, splicing, מודיפיקציות ובקרת איכות |
| rRNA | בעיקר Pol I, ובנוסף Pol III עבור 5S rRNA | חיתוך של precursor ארוך, מודיפיקציות והרכבה עם חלבונים ריבוזומליים |
| tRNA | Pol III | חיתוך קצוות, הוספת CCA, מודיפיקציות רבות וקיפול למבנה תלתני |
ב־mRNA, העיבוד קשור מאוד לשעתוק עצמו. הזנב CTD של Pol II מגייס חלבונים שמבצעים capping, splicing ו־polyadenylation בזמן שה־RNA עדיין יוצא מהפולימראז. לכן השעתוק והעיבוד מתרחשים במקביל ומשפיעים זה על זה.
עיבוד mRNA: שלושת השלבים העיקריים
תעתיק ראשוני של גן המקודד לחלבון נקרא pre-mRNA. הוא כולל אקסונים, אינטרונים וקצוות שעדיין צריכים לעבור עיבוד. כדי להפוך ל־mRNA בוגר, הוא עובר שלושה תהליכים עיקריים:
5′ capping → splicing → polyadenylation
| תהליך | מה קורה בו | למה הוא נדרש |
|---|---|---|
| 5′ capping | הוספת cap של 7-methylguanosine בקצה 5′ | יציבות, יציאה מהגרעין, התחלת תרגום |
| Splicing | הוצאת אינטרונים וחיבור אקסונים | יצירת מסגרת קריאה תקינה |
| Polyadenylation | חיתוך קצה 3′ והוספת זנב Poly-A | יציבות, יציאה מהגרעין, תרגום ובקרה על אורך חיי ה־mRNA |
בנוסף לשלושת התהליכים האלה, mRNA יכול לעבור מודיפיקציות כימיות על גבי הנוקלאוטידים עצמם. אלה שינויים בתעתיק עצמו, ולא “תוספת קצה” כמו cap או Poly-A.
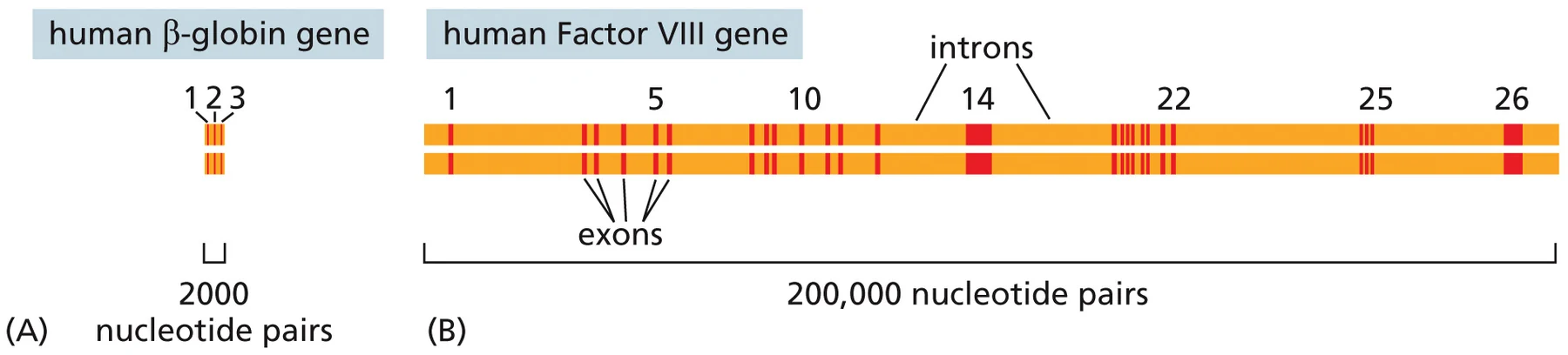
בדוגמה מהמצגת רואים את ההבדל בין גן קצר כמו β-globin, שיש בו מעט אקסונים ואינטרונים, לבין גן ארוך כמו Factor VIII, שיש בו 26 אקסונים ו־25 אינטרונים. ככל שיש יותר אינטרונים, תהליך ה־splicing נעשה חשוב יותר ליצירת mRNA מדויק.
למבחן: אם גן מכיל אינטרון, האינטרון צריך לצאת החוצה. ללא splicing תקין, מסגרת הקריאה עלולה להשתבש, עלול להיווצר stop codon מוקדם, או שעלול להיווצר mRNA שיישלח לפירוק.
5′ capping
5′ cap הוא גואנוזין שעבר מתילציה, המחובר לנוקלאוטיד הראשון של ה־RNA בקשר הפוך וייחודי: 5′-to-5′ triphosphate linkage. הקשר הזה שונה מהקשרים הרגילים בתוך RNA, שבהם נוקלאוטיד אחד מחובר לנוקלאוטיד הבא בכיוון 5′←3′ (חמש לשלוש).
ה־cap מתווסף בתחילת ההתארכות של Pol II, כאשר ה־RNA החדש כבר יצא מעט מהפולימראז. בשלב הזה חלבונים שקשורים ל־CTD ול־SPT5 מגייסים את פקטורי ה־capping.
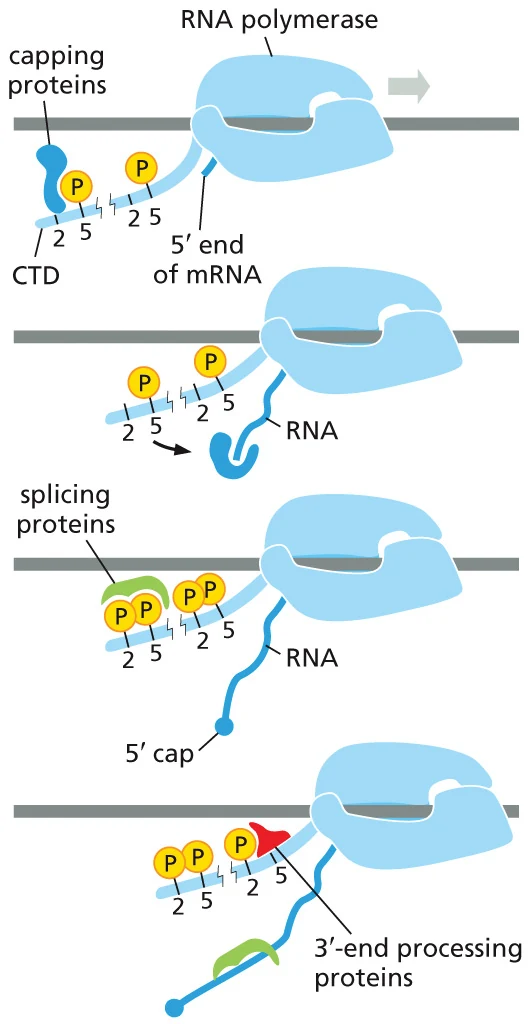
שלושת האנזימים של capping
| שלב | אנזים | פעולה |
|---|---|---|
| 1 | RNA 5′ triphosphatase | מוריד פוספט אחד מקצה 5′ של ה־pre-mRNA |
| 2 | Guanylyltransferase (GT) | מחבר GMP לקצה ה־RNA בקשר 5′-to-5′ |
| 3 | Methyltransferase (MT) | מוסיף מתיל ל־G שנוסף, ולעיתים גם לריבוז של הנוקלאוטיד הראשון |
בשלב השני, האנזים משתמש ב־GTP. הוא מסיר ממנו שני פוספטים, כך שנשאר GMP, ומחבר אותו ל־RNA. לאחר מכן מתווספות קבוצות מתיל, ולכן ה־cap הבוגר נקרא בדרך כלל m⁷G cap.
תפקידי ה־cap
| תפקיד | הסבר |
|---|---|
| הגנה מפירוק | קצה RNA חשוף יכול לעבור דגרדציה; ה־cap מגן על קצה 5′ |
| יציאה מהגרעין | חלבוני cap-binding מזהים RNA שעבר capping ועוזרים לו לעבור דרך ה־nuclear pore |
| תחילת תרגום | בציטופלזמה, פקטורי התחלת תרגום מזהים את ה־cap ומגייסים את הריבוזום |
למבחן: cap הוא לא רק סימון דקורטיבי בקצה ה־RNA. הוא משתתף ביציבות, export ותרגום. mRNA ללא cap תקין צפוי להיות פחות יציב ולעבור בקרה/פירוק.
מודיפיקציות על mRNA
מעבר ל־cap, התעתיק עצמו יכול לעבור מודיפיקציות כימיות. חלק מהן מתווספות תוך כדי שעתוק וחלק לאחר השעתוק. בשיעור לא נדרש לזכור את כל סוגי המודיפיקציות בעל פה, אבל כן חשוב להכיר את העיקרון: שינוי קטן בבסיס או בריבוז יכול להשפיע על יציבות RNA, splicing, export ותרגום.
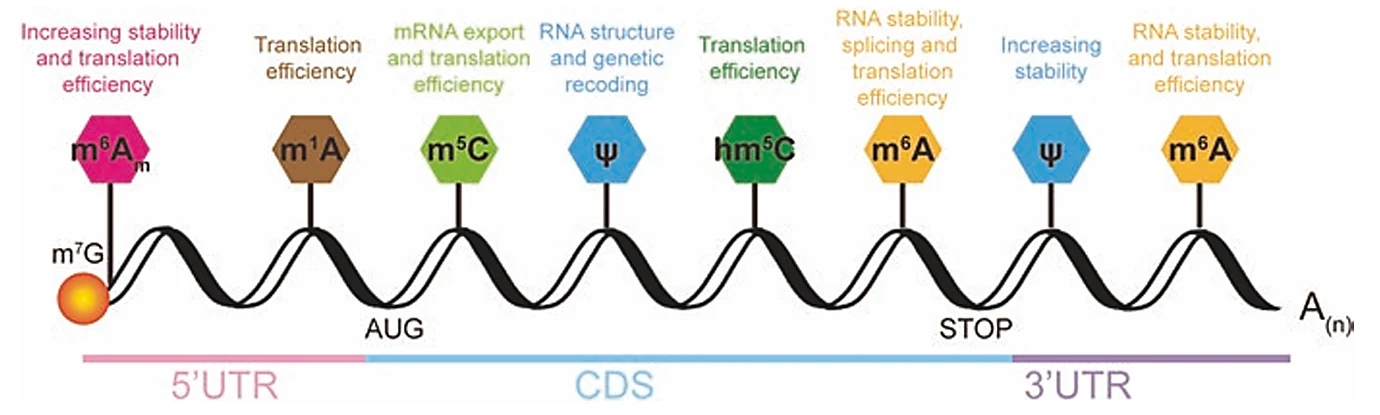
דוגמאות:
| מודיפיקציה | מה משתנה | מאפיינים כלליים |
|---|---|---|
| m⁶A | מתילציה בעמדה 6 של אדנין | אחת המודיפיקציות הידועות ב־mRNA; קשורה ליציבות, splicing, export ותרגום |
| m⁶Am | מודיפיקציה דומה, לעיתים סמוכה ל־cap | קשורה בין היתר ליציבות ותרגום |
| Pseudouridine (Ψ) | שינוי בקישור של uridine לריבוז, כך שהקשר עובר דרך פחמן | מופיעה גם ב־rRNA וב־tRNA; קשורה ליציבות וקיפול RNA |
| m⁵C, m¹A ועוד | מתילציות בעמדות שונות | חלק ממערך רגולציה רחב של RNA |
המיקום של מודיפיקציה והאם היא תתווסף לתעתיק מסוים תלויים בהקשר התאי. אותו mRNA יכול לעבור מודיפיקציה בתנאי תא אחד ולא לעבור אותה בתנאי אחר. כך התא יכול להשפיע על אורך חיי ה־RNA, על יציאתו מהגרעין ועל יעילות התרגום שלו.
Polyadenylation וזנב Poly-A
Polyadenylation הוא תהליך עיבוד קצה 3′ של mRNA. בניגוד ל־cap, שנוסף לקצה 5′, כאן התא קובע היכן לחתוך את ה־RNA בקצה 3′ ומוסיף אחר כך זנב ארוך של אדנינים.
האותות שמגדירים את אזור החיתוך וה־polyadenylation מקודדים ב־DNA, אבל הם מזוהים לאחר שהם כבר שועתקו ל־RNA. הרצף המרכזי הוא PAS - Polyadenylation Signal, ובדרך כלל מופיע בו המוטיב AAUAAA.
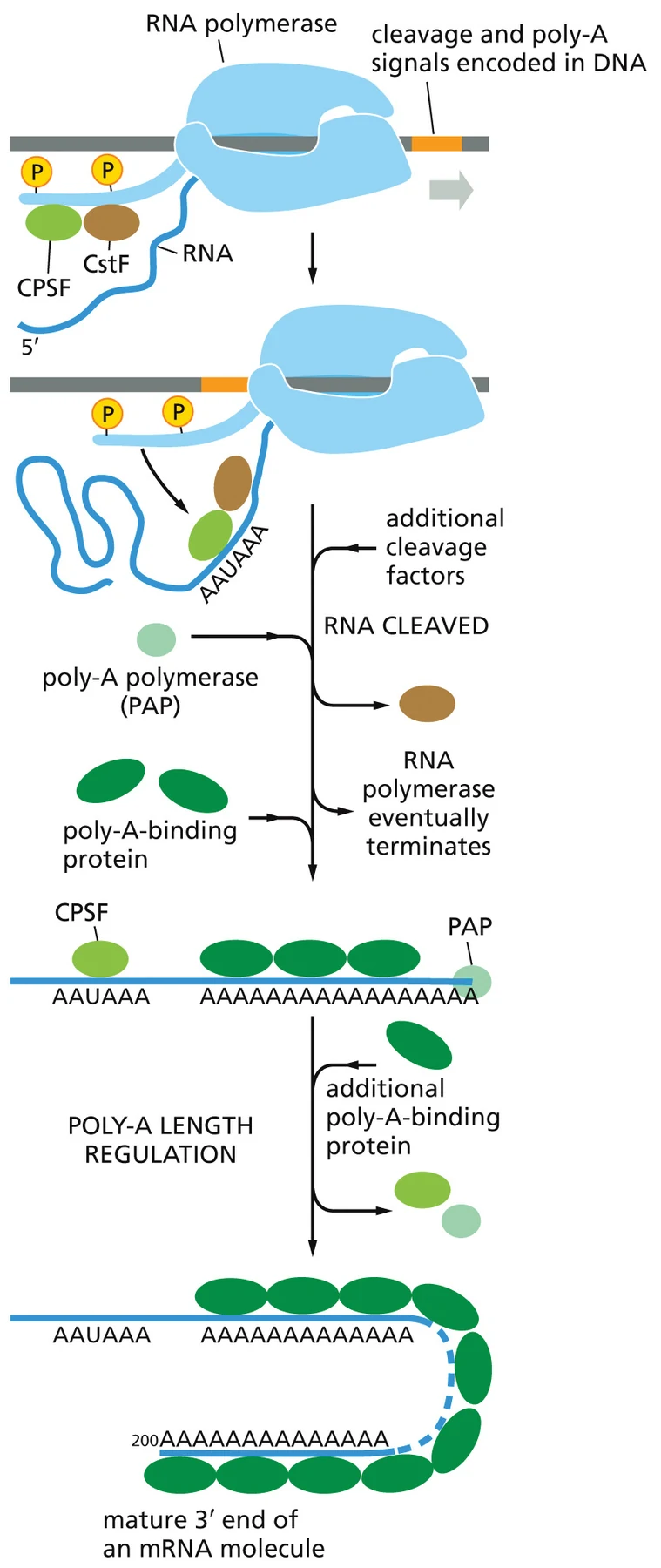
רצף האירועים
| שלב | מה מתרחש |
|---|---|
| 1 | Pol II עובר על אזור PAS בקצה 3′ של הגן |
| 2 | חלבוני CPSF ו־CstF, כחלק מ־CPA complex, עוברים מה־CTD אל ה־RNA |
| 3 | קומפלקס העיבוד מגייס חלבונים נוספים ומבצע cleavage של ה־RNA |
| 4 | Poly-A Polymerase (PAP) מוסיף נוקלאוטידי A לקצה החדש |
| 5 | Poly-A-binding proteins (PABPs) נקשרים לזנב ומסייעים לקבוע את אורכו |
PAP אינו משתמש בתבנית DNA. לכן זנב ה־Poly-A עצמו אינו מקודד ישירות בגנום. הוא מתווסף לאחר החיתוך, בעזרת ATP, ובדרך כלל מגיע לאורך של בערך 200 נוקלאוטידים.
תפקידי Poly-A
| תפקיד | הסבר |
|---|---|
| יציבות | ככל שהזנב ארוך יותר, לרוב ה־mRNA יציב יותר; בציטופלזמה הזנב מתקצר בהדרגה |
| יציאה מהגרעין | PABPs וחלבונים נוספים עוזרים ל־mRNA לעבור export |
| תרגום | בציטופלזמה זנב Poly-A משתתף בהתחלת תרגום דרך אינטראקציות עם פקטורי תרגום |
למבחן: רצף ה־PAS מקודד ב־DNA ומופיע ב־RNA, אבל זנב ה־Poly-A עצמו נוסף אחרי cleavage ואינו מועתק מתבנית DNA.
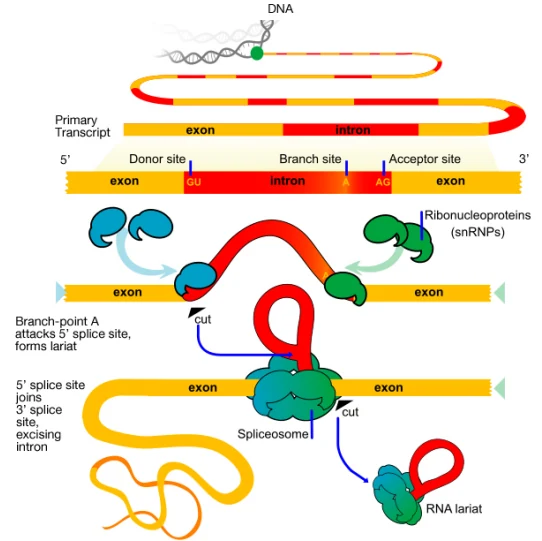
Splicing: הוצאת אינטרונים וחיבור אקסונים
Splicing הוא התהליך שבו אינטרונים מוסרים מתוך pre-mRNA, ואקסונים סמוכים מחוברים זה לזה. התוצר הוא mRNA בוגר שבו רצפי האקסונים מסודרים ברצף המתאים למסגרת הקריאה.
האנזים-קומפלקס שמבצע את הספלייסינג נקרא spliceosome. הוא מורכב מ־RNA וחלבונים, ולכן מזכיר מבחינה רעיונית קומפלקסים כמו הריבוזום: לא מדובר רק בחלבון אחד שמבצע תגובה, אלא במכונה מולקולרית גדולה ודינמית.
ה־spliceosome מבצע שלוש פעולות עיקריות:
- מזהה את גבולות האינטרון.
- מקרב את האקסון שלפני האינטרון לאקסון שאחריו.
- מוציא את האינטרון ומחבר את האקסונים.
התהליך מתרחש במקרים רבים תוך כדי שעתוק. Pol II ממשיך לסנתז RNA, ובמקביל חלבוני splicing מגויסים לתעתיק החדש, בין היתר דרך ה־CTD.
רצפי ההכרה של הספלייסינג
כדי להוציא את האינטרון במקום הנכון, התא צריך לזהות שלושה אזורים ב־pre-mRNA:
| אזור | מיקום | מאפיין רצפי |
|---|---|---|
| 5′ splice site | תחילת האינטרון | לרוב מתחיל ב־GU |
| Branch point | בתוך האינטרון, קרוב יותר לקצה 3′ | כולל אדנוזין חשוב (A) |
| 3′ splice site | סוף האינטרון | לרוב מסתיים ב־AG; לפניו אזור עשיר בפירימידינים, בעיקר C ו־U |
הרצף הכללי:
AG|GUAAGU.............CU(A)AC........YYYYYY....AG|G
הסימון Y מציין פירימידין, כלומר C או U. האדנוזין ב־branch point הוא זה שייצור בהמשך את מבנה ה־lariat.
למבחן: GU בתחילת האינטרון, AG בסופו, ו־A ב־branch point. אלה שלושת האזורים שמכוונים את ה־spliceosome למקום הנכון.
הרכבת ה־spliceosome
ה־spliceosome בנוי מ־snRNPs - small nuclear ribonucleoproteins. כל snRNP כולל snRNA וחלבונים. חמשת ה־snRNPs העיקריים הם:
| snRNP | תפקיד |
|---|---|
| U1 | נקשר ל־5′ splice site |
| U2 | נקשר ל־branch point לאחר BBP ו־U2AF |
| U4 | מעכב זמנית את U6, כדי שהפעילות הקטליטית לא תתרחש מוקדם מדי |
| U5 | מקרב וממקם את שני האקסונים לקראת החיבור |
| U6 | משתתף בליבה הקטליטית של ה־spliceosome, יחד עם U2 ו־NTC/NTR |
בנוסף ל־snRNPs יש חלבונים נוספים:
| חלבון / קומפלקס | תפקיד |
|---|---|
| BBP - Branch-point Binding Protein | מזהה את ה־branch point בתחילת ההרכבה |
| U2AF - U2 Auxiliary Factor | נקשר לאזור ה־polypyrimidine tract ול־3′ splice site, ועוזר לגייס U2 |
| NTC/NTR | משתתפים ביצירת המצב הקטליטי הפעיל של ה־spliceosome |
| EJC - Exon Junction Complex | מתווסף לאחר splicing באזור חיבור אקסון-אקסון ומשמש לבקרה בהמשך |
סדר ההרכבה
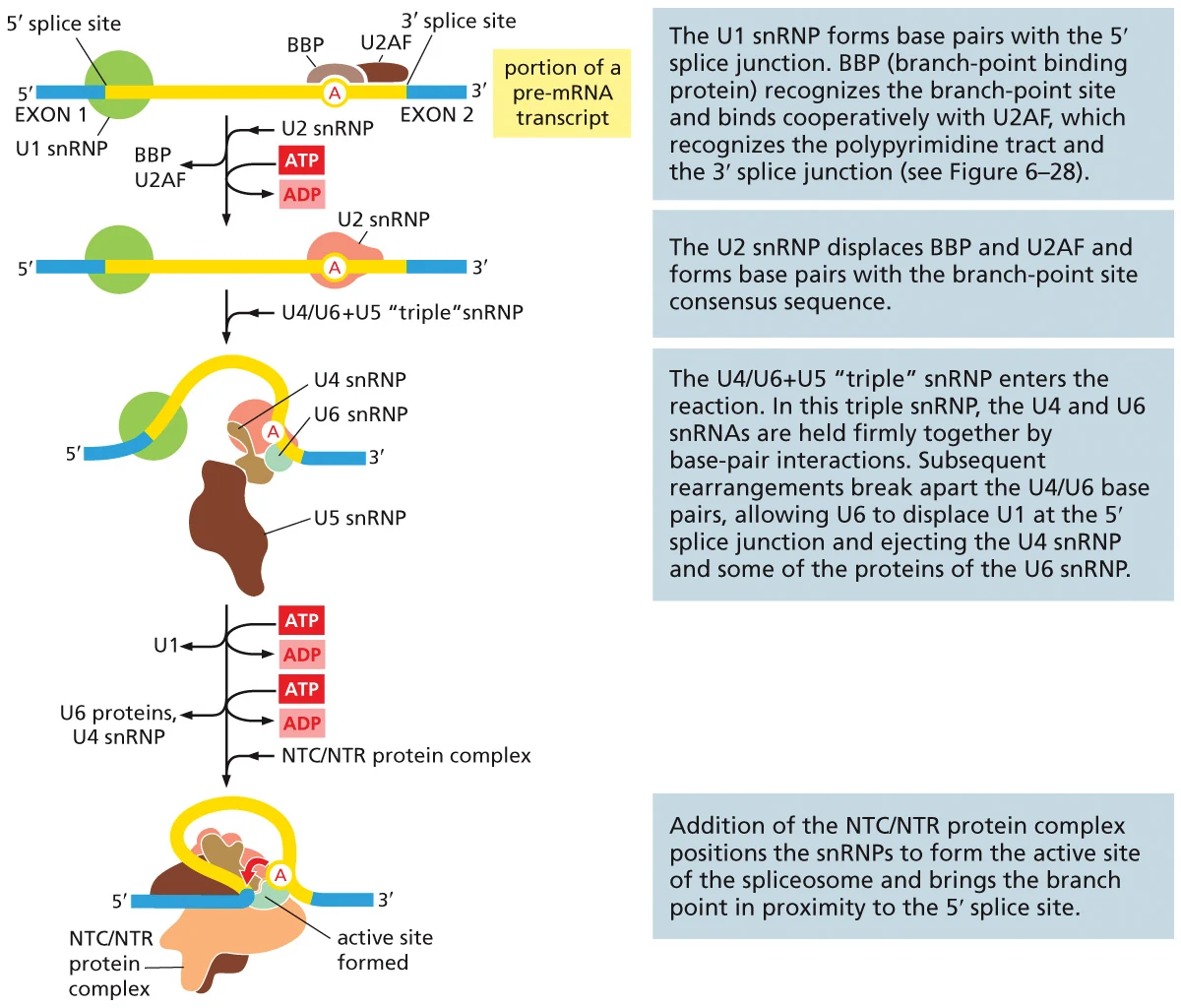
- U1 נקשר ל־5′ splice site.
- BBP ו־U2AF נקשרים באזור ה־branch point וה־3′ splice site.
- U2 מחליף את BBP/U2AF ונקשר ל־branch point.
- קומפלקס U4/U6 ו־U5 מצטרף.
- U5 מקרב את האקסונים.
- U1 ו־U4 עוזבים. U4 מפסיק לעכב את U6.
- U6 נקשר מחדש באזור 5′ ויוצר יחד עם U2 את הליבה הקטליטית.
- NTC/NTR מייצבים את המצב הפעיל ומאפשרים את תגובות החיתוך.
ה־ATP משמש בעיקר לשינויי קונפורמציה, לפירוק והרכבה מחדש של אינטראקציות RNA-RNA ו־RNA-חלבון, ולמיחזור רכיבי ה־spliceosome.
שתי תגובות החיתוך וה־lariat
לאחר שה־spliceosome פעיל, מתרחשות שתי תגובות עיקריות:
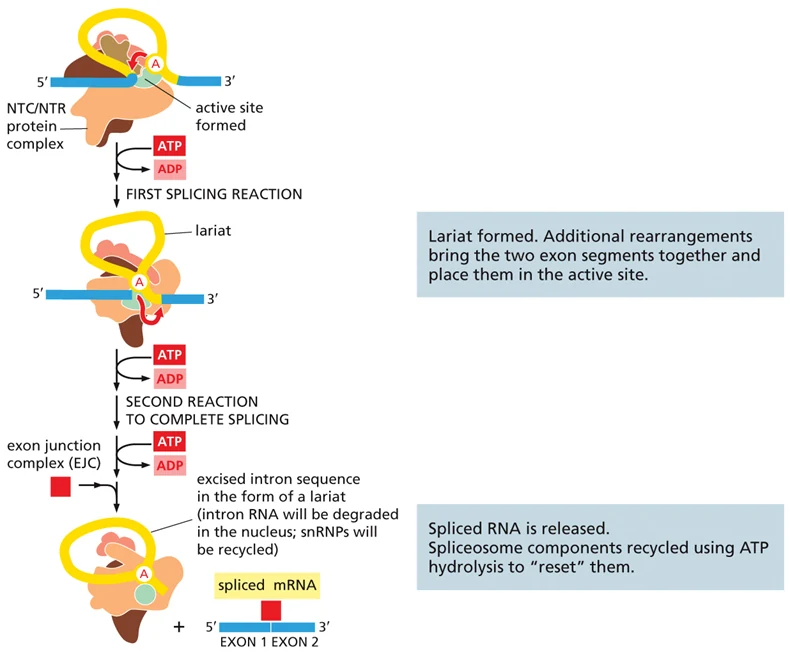
| תגובה | מה קורה |
|---|---|
| תגובה ראשונה | קצה 5′ של האינטרון נחתך. האדנוזין ב־branch point נקשר לקצה האינטרון ויוצר מבנה lariat |
| תגובה שנייה | קצה 3′ של האינטרון נחתך, שני האקסונים מחוברים, והאינטרון יוצא כ־lariat |
לאחר החיבור, האינטרון בצורת lariat יפורק בגרעין. רכיבי ה־spliceosome ממוחזרים לשימוש נוסף. באזור החיבור בין שני האקסונים מתווסף EJC, שישמש בהמשך לבקרה על mRNA שעבר splicing.
למבחן: ה־spliceosome אינו רק “מקרב אקסונים”. הוא מזהה splice sites, יוצר ליבה קטליטית עם U2/U6, מבצע שתי תגובות חיתוך, מוציא lariat ומסמן את חיבור האקסונים בעזרת EJC.
Alternative splicing
ב־alternative splicing, אותו גן יכול ליצור כמה mRNAs שונים. ההבדל בין ה־mRNAs נובע מבחירה שונה של אקסונים שייכללו בתעתיק הבוגר.
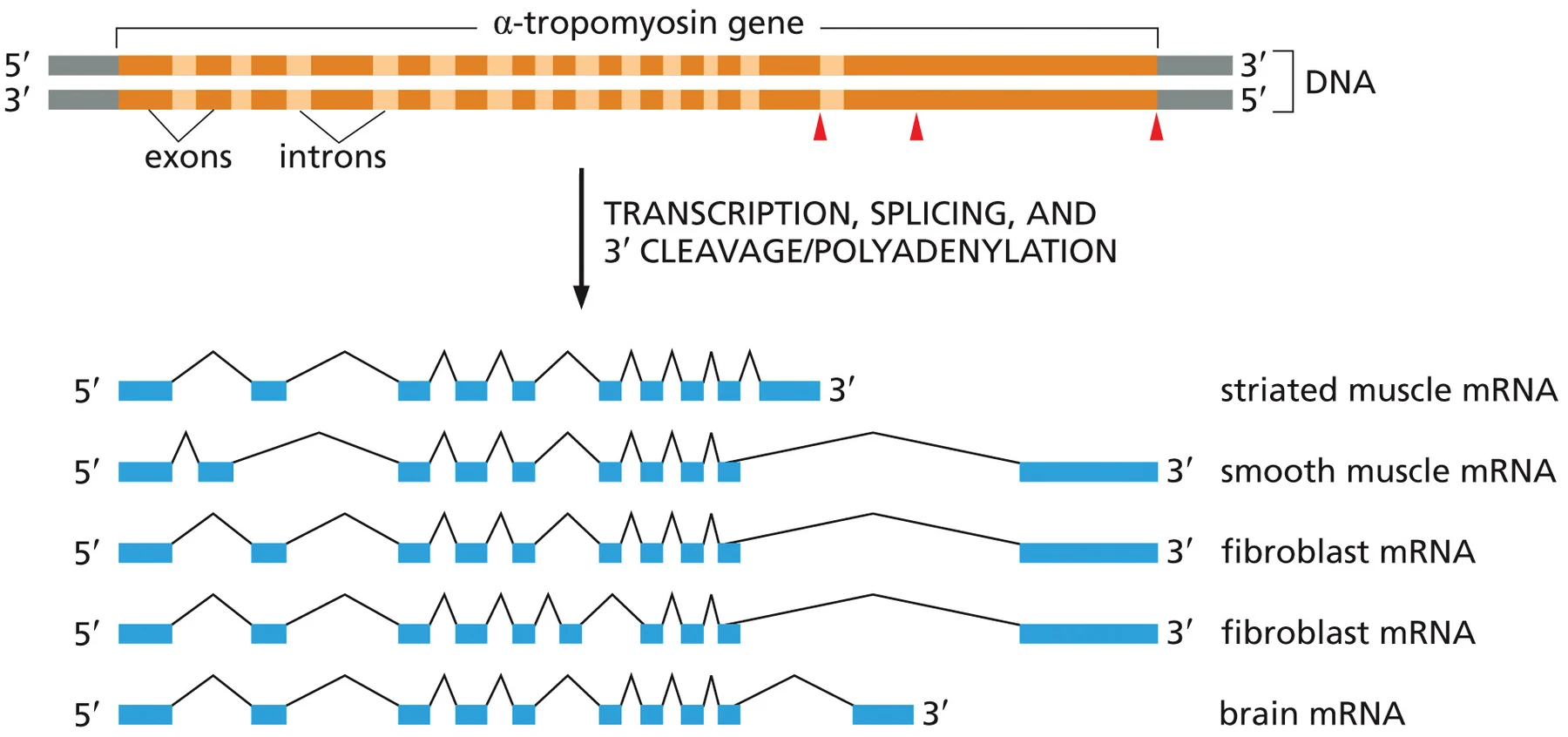
בדוגמה של α-tropomyosin, אותו גן עובר splicing בצורות שונות בתאים שונים:
- תעתיק אחד מתאים לסוגי שריר מסוימים.
- תעתיק אחר מתאים לשריר חלק.
- תעתיקים אחרים נוצרים בפיברובלסטים או בתאי מוח.
כך התא יכול להשתמש באותו גן כדי לייצר איזופורמים שונים של חלבון. זה חוסך צורך בגן נפרד לכל וריאציה של חלבון, ומאפשר התאמה לרקמה, לשלב התפתחותי או למצב פיזיולוגי.
Alternative splicing יכול לכלול:
| סוג שינוי | תיאור |
|---|---|
| Exon skipping | אקסון מסוים מדולג ולא נכנס ל־mRNA הבוגר |
| Alternative 5′ splice site | שימוש בנקודת חיתוך אחרת בתחילת אינטרון |
| Alternative 3′ splice site | שימוש בנקודת חיתוך אחרת בסוף אינטרון |
| Alternative polyadenylation | שימוש באתר poly-A שונה, ולכן קצה 3′ שונה |
הבחירה בין אפשרויות splicing מושפעת מרצפי RNA ומחלבוני בקרה שנקשרים אליהם. רצפי splice חזקים מזוהים ביעילות גבוהה יותר, ורצפים חלשים תלויים יותר בפקטורי בקרה.
בקרת איכות, יציאה מהגרעין ו־NMD
mRNA בוגר אינו יוצא מהגרעין רק מפני שהוא סונתז. עליו לשאת סימנים שמראים שהוא עבר עיבוד תקין: cap, זנב Poly-A עם PABPs, חלבונים שנקשרים בזמן splicing, וחלבוני export.
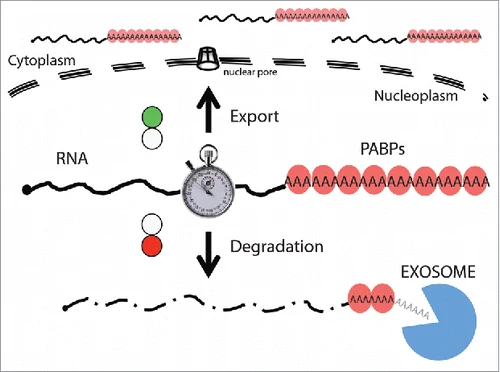
יציאה מהגרעין או פירוק
| מצב ה־RNA | גורל |
|---|---|
| mRNA עם cap, Poly-A, PABPs וחלבוני עיבוד מתאימים | עובר export לציטוזול |
| RNA לא מעובד או פגום | נשמר בגרעין ונשלח לפירוק |
| שברי RNA, אינטרונים, תעתיקים לא בשלים | מפורקים בגרעין, בין היתר על ידי nuclear RNA exosome |
Nuclear RNA exosome
ה־nuclear RNA exosome הוא קומפלקס חלבוני גדול שמפרק RNA בגרעין. הוא בנוי כמעין תעלה/חבית חלבונית: RNA נכנס אל הקומפלקס, מגיע לתת־יחידות קטליטיות בעלות פעילות RNase, ומתפרק לנוקלאוטידים.
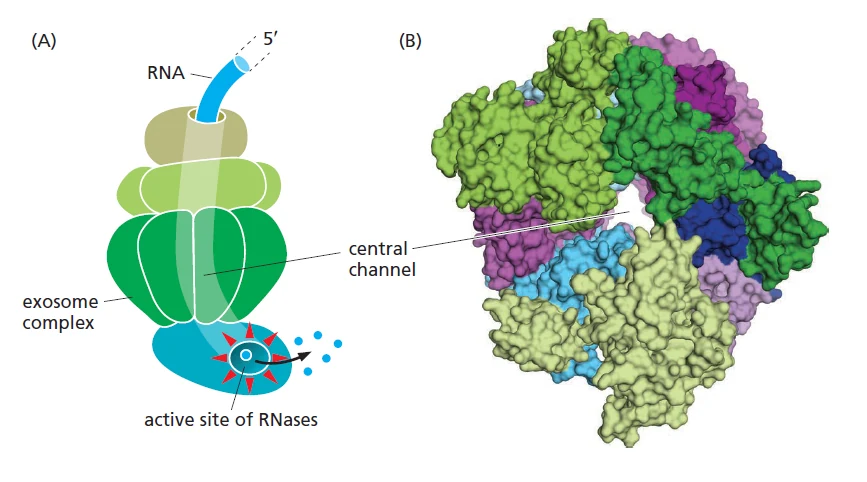
ה־exosome מפרק בין היתר:
- אינטרונים שנחתכו החוצה.
- RNAs לא בשלים.
- שברי RNA.
- תעתיקים שלא עברו עיבוד תקין.
Nonsense-mediated decay (NMD)
גם אחרי export לציטוזול יש בקרת איכות. אחת הבקרות שנלמדו היא Nonsense-mediated decay, או NMD. היא מזהה mRNA שבו יש stop codon מוקדם, מצב שעלול ליצור חלבון קצר ולא תקין.
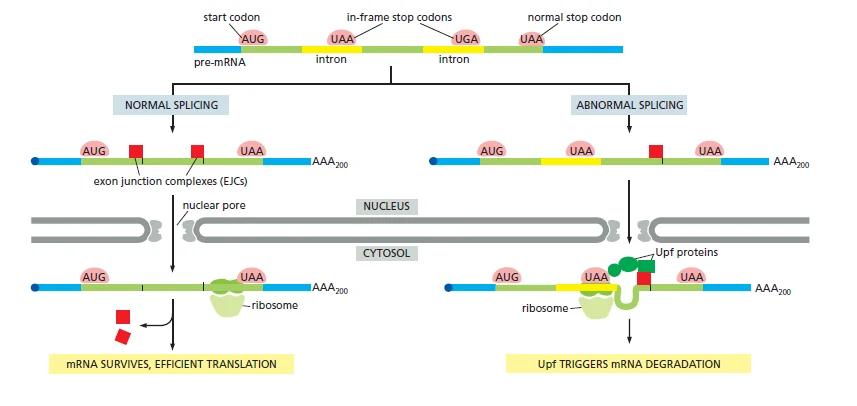
הרעיון:
- לאחר splicing, אזורי חיבור אקסון-אקסון מסומנים על ידי EJC.
- לאחר export, ריבוזום מבצע סריקה/תרגום ראשון של ה־mRNA.
- אם מסגרת הקריאה תקינה, הריבוזום עובר לאורך ה־mRNA ומסיר את ה־EJCs.
- אם יש stop codon מוקדם, הריבוזום נעצר לפני שהוא הסיר EJC שנמצא downstream.
- EJC שנשאר על mRNA אחרי עצירה מוקדמת מגייס חלבוני Upf ומפעיל פירוק של ה־mRNA.
למבחן: EJC הוא סימן לכך שהתרחש splicing באזור מסוים. ב־NMD, EJC שנשאר אחרי stop codon מוקדם משמש כסימן ל־mRNA בעייתי.
עיבוד rRNA והרכבת ריבוזומים
החלק השני של השיעור עובר ל־rRNA. בשיעור הקודם נלמד ש־Pol I מסנתז precursor ארוך של rRNA בגרעינון. בשיעור הזה הושלם מה קורה ל־precursor הזה לאחר השעתוק.
מה־47S precursor ל־rRNA בוגר
Pol I מסנתז 47S pre-rRNA. התעתיק הזה כולל את הרצפים שמהם ייווצרו:
- 18S rRNA - חלק מהתת־יחידה הקטנה של הריבוזום.
- 5.8S rRNA - חלק מהתת־יחידה הגדולה.
- 28S rRNA - חלק מהתת־יחידה הגדולה.
בין הרצפים האלה נמצאים spacers:
| Spacer | מיקום | מה קורה לו |
|---|---|---|
| 5′ ETS | בתחילת התעתיק | מוסר בשלבי העיבוד הראשונים |
| ITS1 | בין 18S לבין 5.8S | מוסר בחיתוכים פנימיים |
| ITS2 | בין 5.8S לבין 28S | מוסר בחיתוכים פנימיים |
| 3′ ETS | בסוף התעתיק | מוסר בשלבי העיבוד |
חשוב להבדיל בין spacers ב־rRNA לבין אינטרונים ב־mRNA. כאן מדובר באזורים שנמצאים בתוך precursor של rRNA ומוסרים בתהליך עיבוד ריבוזומלי.
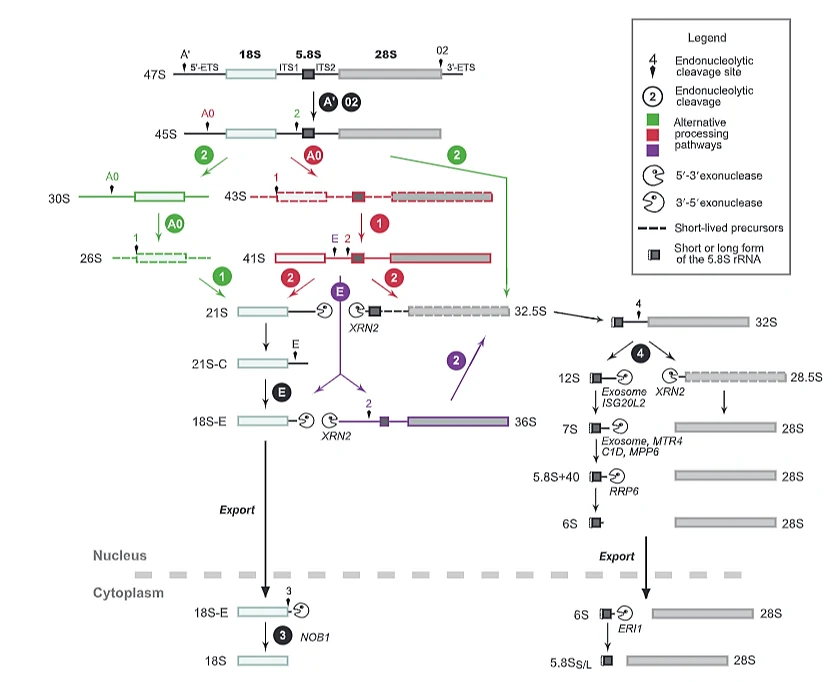
רצף העיבוד:
47S pre-rRNA45S לאחר הסרת ETSחיתוך בין 18S ל־5.8S/28S18S לתתי-יחידה קטנה5.8S + 28S לתתי-יחידה גדולה
ה־18S עובר מסלול עיבוד שמוביל לתת־היחידה הקטנה 40S. ה־5.8S וה־28S מעובדים יחד עם 5S rRNA, שמיוצר על ידי Pol III, ויחד הם משתתפים ביצירת התת־יחידה הגדולה 60S.
מודיפיקציות rRNA
rRNA מכיל הרבה מודיפיקציות. שתי מודיפיקציות שהוזכרו במיוחד:
| מודיפיקציה | מה משתנה | תפקיד כללי |
|---|---|---|
| 2′-O-methylation | מתילציה על הריבוז | ייצוב מבנה RNA וסיוע בבניית ריבוזום תקין |
| Pseudouridylation | uridine הופך ל־pseudouridine | קיפול נכון ויציבות rRNA |
מודיפיקציות rRNA חשובות לקיפול, יציבות והרכבת ריבוזום פעיל. הן מתרחשות כחלק מתהליך הביוגנזה של הריבוזום בגרעינון.
הרכבת הריבוזום דורשת את שלוש הפולימראזות
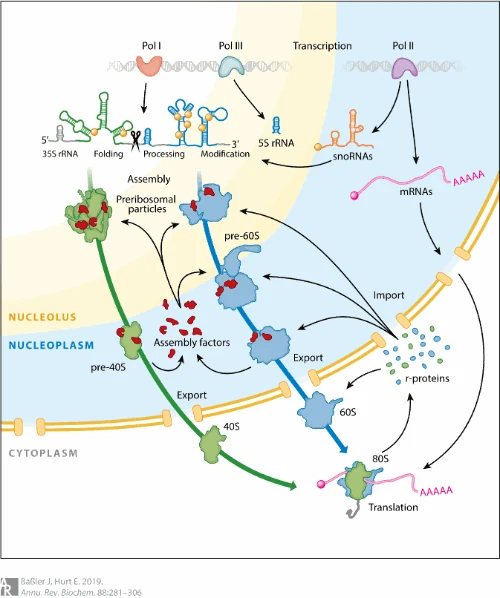
הרכבת הריבוזום משלבת כמה מקורות:
| מקור | תרומה לריבוזום |
|---|---|
| Pol I | מסנתז את 47S pre-rRNA, שממנו נוצרים 18S, 5.8S ו־28S |
| Pol III | מסנתז 5S rRNA |
| Pol II | מסנתז mRNAs לחלבוני הריבוזום וגם RNAs רגולטוריים כמו snoRNAs |
חלבוני הריבוזום מסונתזים בציטופלזמה, כי הם תוצרי תרגום של mRNAs. לאחר מכן הם חוזרים לגרעין בעזרת NLS - Nuclear Localization Signal, מגיעים לגרעינון, ונקשרים ל־rRNA תוך כדי עיבודו.
הריבוזום אינו יוצא מהגרעין כ־rRNA חופשי. תתי-היחידות נבנות בהדרגה בגרעינון, עוברות שלבי maturation נוספים, ואז יוצאות בנפרד לציטופלזמה. שם, בזמן תרגום, תת־היחידה הקטנה ותת־היחידה הגדולה מתחברות על גבי mRNA.
עיבוד tRNA
החלק האחרון של השיעור עוסק ב־tRNA. Pol III מסנתז pre-tRNA, שאינו מוכן מיד לתרגום. כדי להפוך ל־tRNA בוגר הוא צריך לעבור חיתוך קצוות, הוספת CCA ומודיפיקציות רבות.
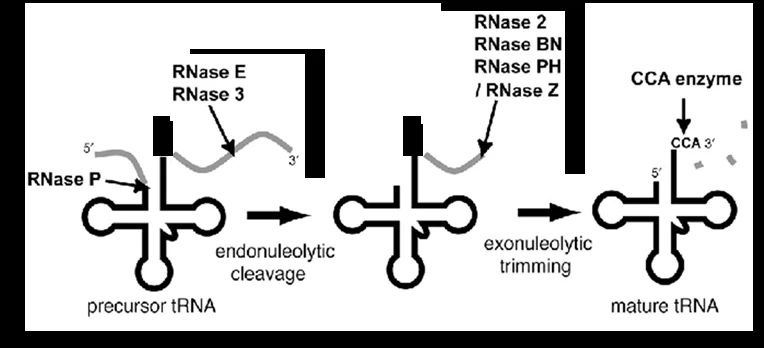
שלבי עיבוד tRNA
| שלב | מה קורה |
|---|---|
| חיתוך 5′ leader | RNase P חותך את הקצה 5′ של pre-tRNA |
| עיבוד 3′ trailer | אנדונוקלאזות ואקסונוקלאזות מסירות רצפים עודפים בקצה 3′ |
| הוספת CCA | אנזים CCA מוסיף את הרצף CCA לקצה 3′ |
| מודיפיקציות כימיות | בסיסים שונים ב־tRNA עוברים שינוי כימי, בעיקר לצורך יציבות וקריאת קודונים |
רצף CCA בקצה 3′ הוא אתר הקישור של חומצת האמינו. לכן הוא חיוני לתפקיד של tRNA בתרגום. האנזים שמוסיף אותו אינו צריך תבנית DNA; הוא מוסיף C, C ואז A בצורה אנזימטית.
למבחן: CCA הוא חלק חיוני מה־tRNA הבוגר כי אליו תיקשר חומצת האמינו. במקרים רבים, הוא נוסף אחרי השעתוק ולא מקודד ישירות ברצף הגן.
מודיפיקציות tRNA
tRNA הוא אחד מסוגי ה־RNA שעוברים הכי הרבה מודיפיקציות. חלקן עוזרות לייצב את המבנה התלתני, וחלקן משפיעות על זיהוי קודון-אנטיקודון.
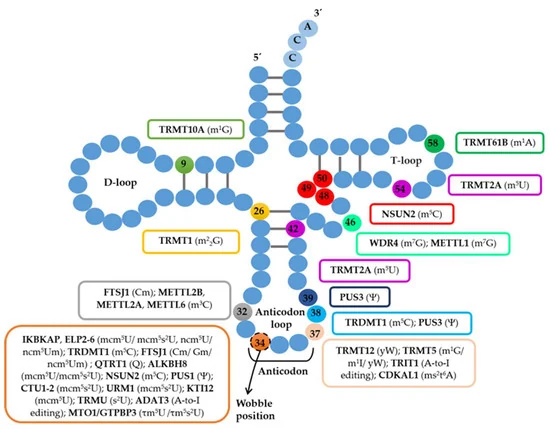
דוגמאות שהוזכרו:
| מודיפיקציה | מאפיין |
|---|---|
| Pseudouridine (Ψ) | שינוי של uridine, מופיע גם ב־rRNA וב־tRNA |
| m²G | מתילציות על guanosine |
| s⁴U | החלפת חמצן בגופרית על uridine |
| 2′-O-methylribose | מתילציה על הריבוז |
| Dihydrouridine (D) | שינוי של uridine, אופייני ל־D-loop |
| Inosine (I) | נוצר מדה־אמינציה של adenosine, נפוץ באזור ה־wobble של האנטיקודון |
Inosine חשוב במיוחד לקריאת קודונים. הוא נוצר כאשר אדנוזין עובר דה־אמינציה, כלומר הסרת קבוצת אמינו והחלפה שלה בחמצן. שינוי כזה מאפשר ל־tRNA לקרוא יותר מאפשרות אחת בקודון, בעיקר בעמדת wobble.
דור פסקל