המעבר מקליבג׳ לגסטרולציה
בשלב הזה מסתיים הדיון ב־Cleavage ומתחילה ההכנה לגסטרולציה (gastrulation). גסטרולציה היא התארגנות מחדש של העובר: תאים מתחילים לזוז, לצאת ממקומות מסוימים, להיכנס בין שכבות, ולייצר את שלוש שכבות הנבט:
| שכבת נבט | |
|---|---|
| אקטודרם | Ectoderm |
| מזודרם | Mesoderm |
| אנדודרם | Endoderm |
לפני שאפשר להתחיל גסטרולציה, העובר צריך להגדיר צירים. בלי צירים, אי אפשר למקם נכון את התאים שייצרו ראש, זנב, גב ובטן.
שלושת הצירים המרכזיים הם:
| ציר | משמעות כללית | מתי נקבע? |
|---|---|---|
| Anterior-Posterior | ראש־זנב | לפני תחילת גסטרולציה |
| Dorsal-Ventral | גב־בטן | לפני תחילת גסטרולציה |
| Left-Right | ימין-שמאל | מאוחר יותר, במהלך גסטרולציה |
השיעור עוסק בעיקר בשני הצירים הראשונים: Anterior-Posterior ו־Dorsal-Ventral. כבר לפני שנראה מבנה ברור של ראש או זנב, העובר צריך לדעת איפה הם יהיו.
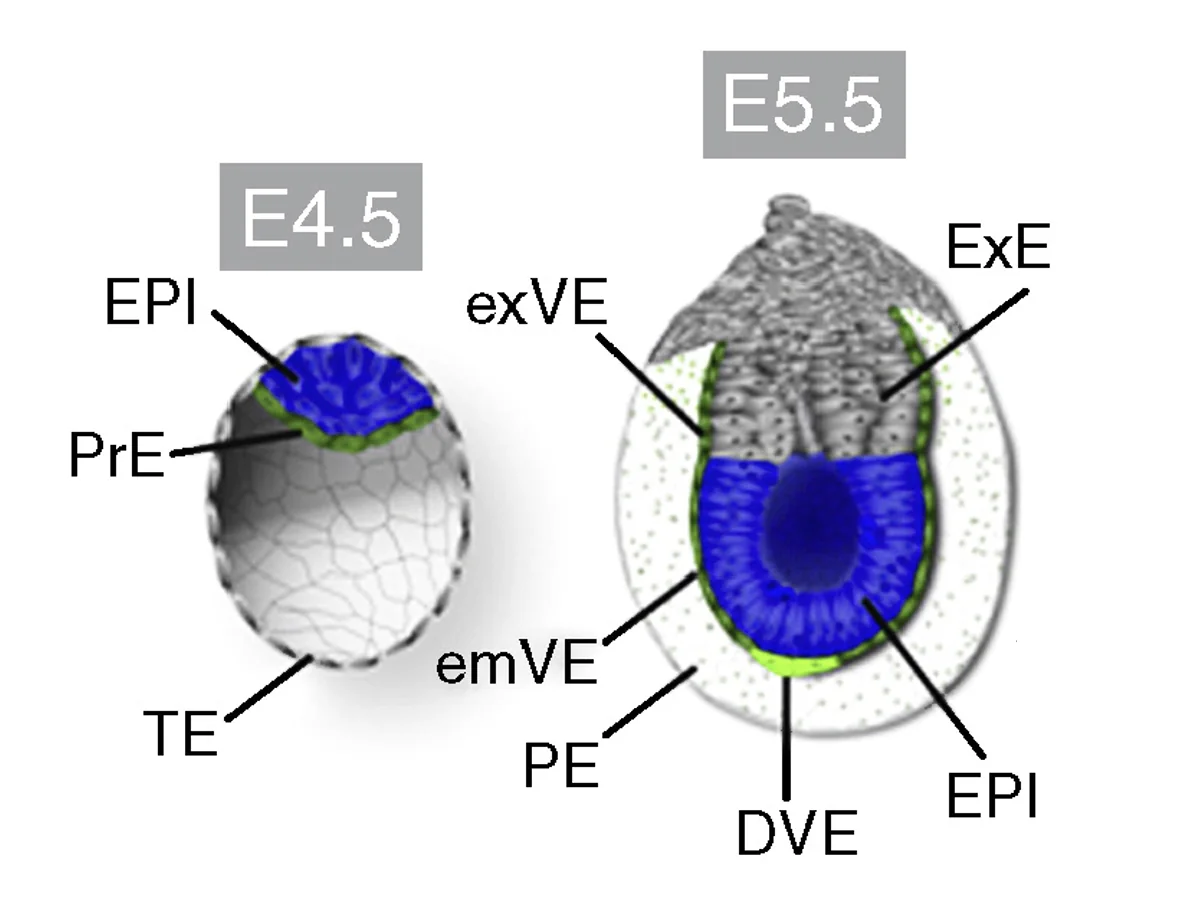
מבלסטוציסט ל־Egg cylinder
שינוי המבנה לאחר השרשה
| Blastocyst |
|---|
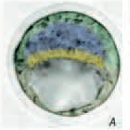 |
| \(\begin{array}{ll} {\color{#0099c8}\blacksquare} & \textit{ICM/epiblast} \\[0.55em] {\color{#00a651}\blacksquare} & \textit{Polar trophectoderm/extra-embryonic ectoderm} \\[0.55em] {\color{#ffd33d}\blacksquare} & \textit{Primitive endoderm/visceral endoderm} \end{array}\) |
בבלסטוציסט יש שלוש אוכלוסיות עיקריות:
| אוכלוסייה | המשך התפתחותי |
|---|---|
| ${\color{#0099c8}\blacksquare}$ Epiblast | ייתן את העובר עצמו |
| ${\color{#ffd33d}\blacksquare}$ Primitive endoderm | ייתן נגזרות חוץ־עובריות כמו visceral endoderm ו־parietal endoderm |
| ${\color{#00a651}\blacksquare}$ Trophectoderm | משתתף בהמשך ביצירת רקמות חוץ־עובריות ושליה (ExE) |
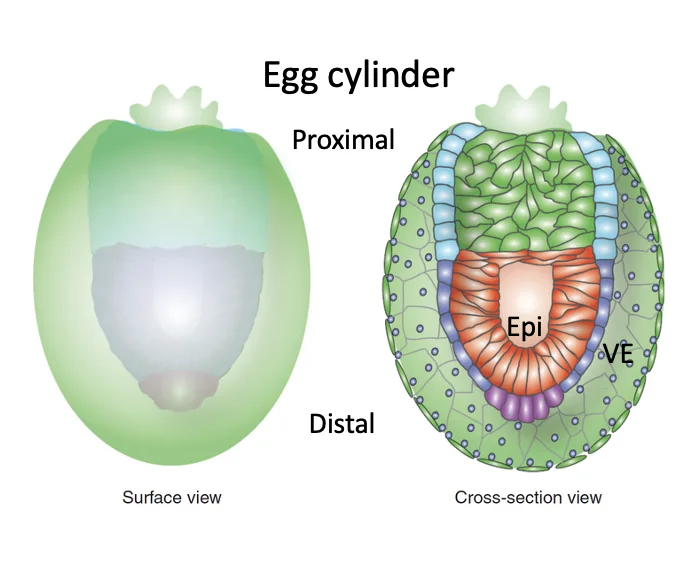
בזמן ההשרשה המבנה עובר שינוי משמעותי: ה־Epiblast עובר פולריזציה, מקבל צד בזאלי וצד אפיקלי, ויוצר חלל במרכז. גם התאים שמקורם ב־polar trophectoderm עוברים חלוקה ופולריזציה, ומהם נוצר Extra-embryonic ectoderm (ExE). בהמשך, החללים באזור האפיבלסט ובאזור ה־ExE, ונוצר מבנה של Egg cylinder.
בתשובה לשאלה של ע.ק.: החלל הזה לא נוצר כמו ה־Blastocoel. בבלסטוציסט ה־Blastocoel נוצר דרך לחץ אוסמוטי ומעבר מים. כאן החלל נוצר בעיקר בגלל פולריזציה וסידור גיאומטרי של תאי האפיבלסט.
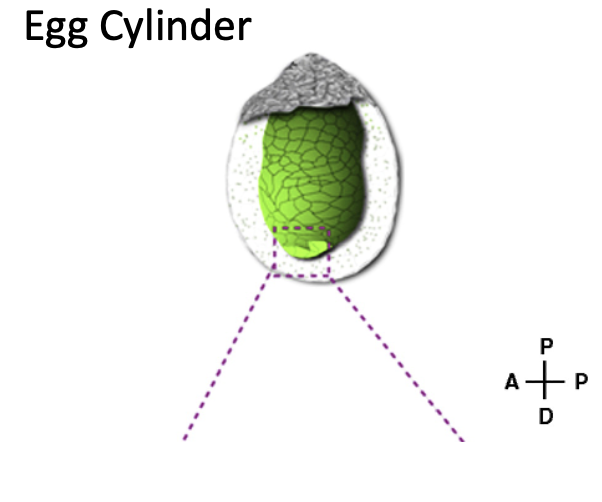
ב־Egg cylinder מופיע ציר ה־Proximal-Distal:
| צד | מיקום |
|---|---|
| Proximal | קרוב לאזור החוץ־עוברי / ExE |
| Distal | הקצה הרחוק יותר, באזור האפיבלסט |
ציר ה־Proximal-Distal מקביל בקירוב לציר שהיה קודם לכן Embryonic-Abembryonic. בשלב הזה ציר ה־Proximal-Distal גם מגדיר את ציר ה־Dorsal-Ventral: הצד האפיקלי של האפיבלסט (${\color{#0099c8}\blacksquare}$) שפונה לתוך החלל (${\color{grey}\blacksquare}$) יהיה הצד הדורסלי, והצד שפונה ל־visceral endoderm (${\color{#ffd33d}\blacksquare}$) יהיה הצד הוונטרלי.
Visceral endoderm ו־Parietal endoderm
מה־Primitive endoderm (הפעם התאים הכחולים באיור למטה, מימין) מתקבלות שתי אוכלוסיות חשובות:
| אוכלוסייה | מיקום / מאפיין |
|---|---|
| Visceral endoderm (VE) ($\color{#62d7e8}\blacksquare , \color{#5866b8}\blacksquare , \color{#a45ac0}\blacksquare$) | עוטף את ה־Egg cylinder מבחוץ ומתנהג כאפיתל |
| Parietal endoderm (${\color{#5866b8}\bullet}$) | נודד ועוטף מבפנים את אזור הטרופובלסט. הנקודות הכחולות שבתוך ה־Exe, להבנתי. |
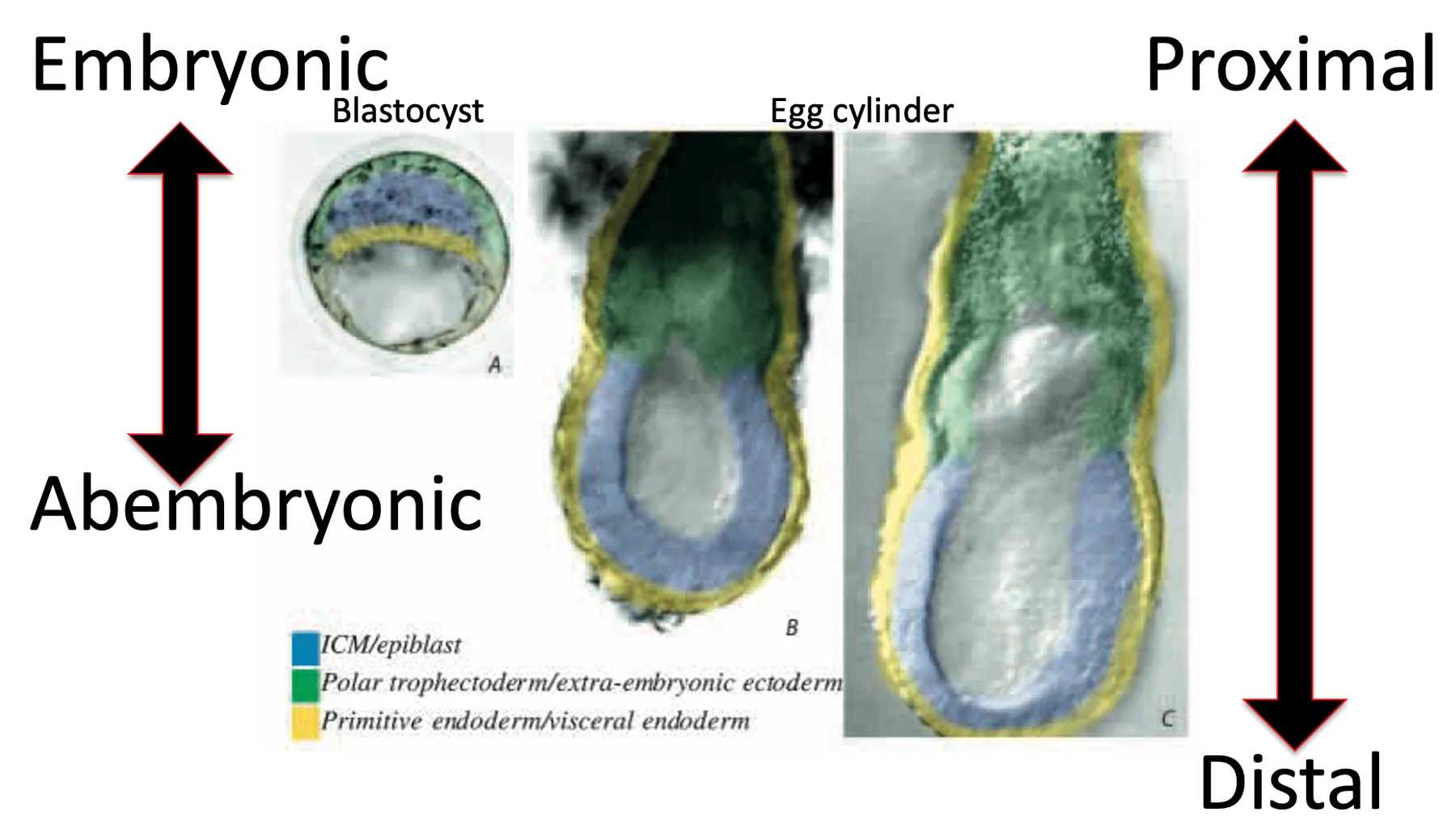
 \[\begin{array}{ll} {\color{#4fd34f}\blacksquare} & \text{Extraembryonic ectoderm} \\[0.6em] {\color{#ff4b2f}\blacksquare} & \text{Epiblast} \\[0.6em] {\color{#5fa7e8}\blacksquare} & \text{Primitive endoderm} \quad \left\{ \begin{array}{ll} {\color{#62d7e8}\blacksquare} & \text{Extraembryonic visceral endoderm} \\[0.5em] {\color{#5866b8}\blacksquare} & \text{Embryonic visceral endoderm} \\[0.5em] {\color{#a45ac0}\blacksquare} & \text{Distal visceral endoderm} \\[0.5em] {\color{#5866b8}\bullet} & \text{Parietal endoderm} \end{array} \right. \end{array}\]
\[\begin{array}{ll} {\color{#4fd34f}\blacksquare} & \text{Extraembryonic ectoderm} \\[0.6em] {\color{#ff4b2f}\blacksquare} & \text{Epiblast} \\[0.6em] {\color{#5fa7e8}\blacksquare} & \text{Primitive endoderm} \quad \left\{ \begin{array}{ll} {\color{#62d7e8}\blacksquare} & \text{Extraembryonic visceral endoderm} \\[0.5em] {\color{#5866b8}\blacksquare} & \text{Embryonic visceral endoderm} \\[0.5em] {\color{#a45ac0}\blacksquare} & \text{Distal visceral endoderm} \\[0.5em] {\color{#5866b8}\bullet} & \text{Parietal endoderm} \end{array} \right. \end{array}\] בשרטוטים של ה־Egg cylinder חשוב להבין אם מסתכלים על העובר מבחוץ או בחתך: מבחוץ, רואים בעיקר את שכבת ה־Visceral endoderm שעוטפת את המבנה; בחתך, רואים גם את ה־Epiblast, ה־ExE והשכבות החוץ־עובריות (הצד הימני של התמונה למעלה).
ה־Visceral endoderm היא לא שכבה אחידה לגמרי; יש בה כמה אזורים עם מורפולוגיה ותפקידים שונים:
| אזור ב־Visceral endoderm | מאפיין |
|---|---|
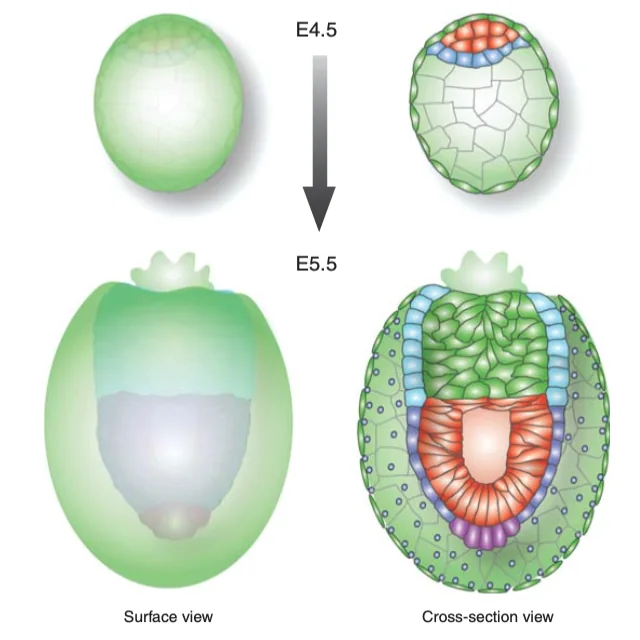
| ${\color{#62d7e8}\blacksquare} \text{Extraembryonic visceral endoderm}$ VE שמקיף את ה־ExE | תאים אפיתליאליים קובואידיים (דמויי קובייה)  |
| ${\color{#5866b8}\blacksquare} \text{Embryonic visceral endoderm}$ VE שמקיף את ה־Epiblast | תאים אפיתליאליים שטוחים יותר, squamous  |
| Distal visceral endoderm (DVE) | תאים בקצה הדיסטלי, בעלי תכונות ייחודיות לעומת שאר ה־VE 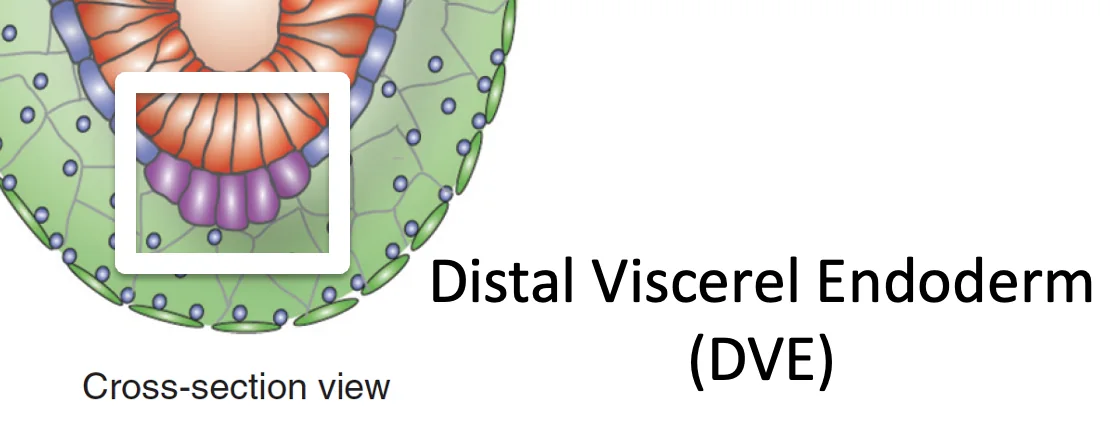 |
השם DVE מגיע מהמיקום: אלה תאי visceral endoderm שנמצאים בנקודה הדיסטלית ביותר. הם אוכלוסייה עם תכונות מולקולריות מיוחדות (לא רק “VE שנמצא בקצה”).
 |  |
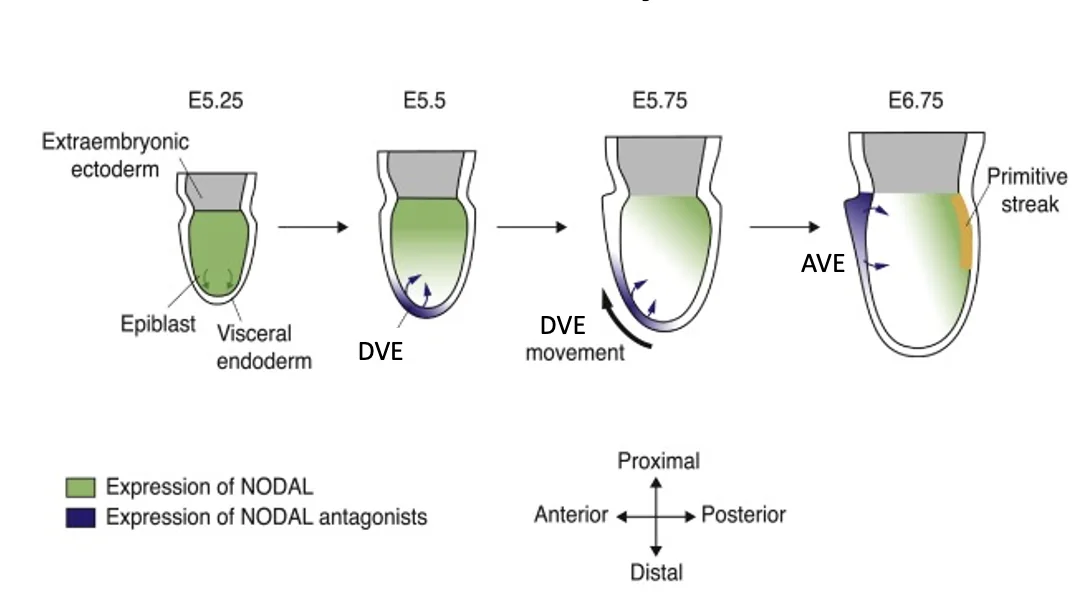
נדידת DVE ויצירת AVE
תאי ה־DVE מתחילים לנדוד מהנקודה הדיסטלית לכיוון פרוקסימלי. הם נעים כקבוצה, ומגיעים עד לקו הגבול בין ה־Epiblast ל־ExE. שם הם נעצרים.
לאחר הנדידה הם נקראים Anterior visceral endoderm (AVE). המקום שבו הם נעצרים מגדיר את הצד האנטריורי של העובר (הצד שבו יהיה הראש).
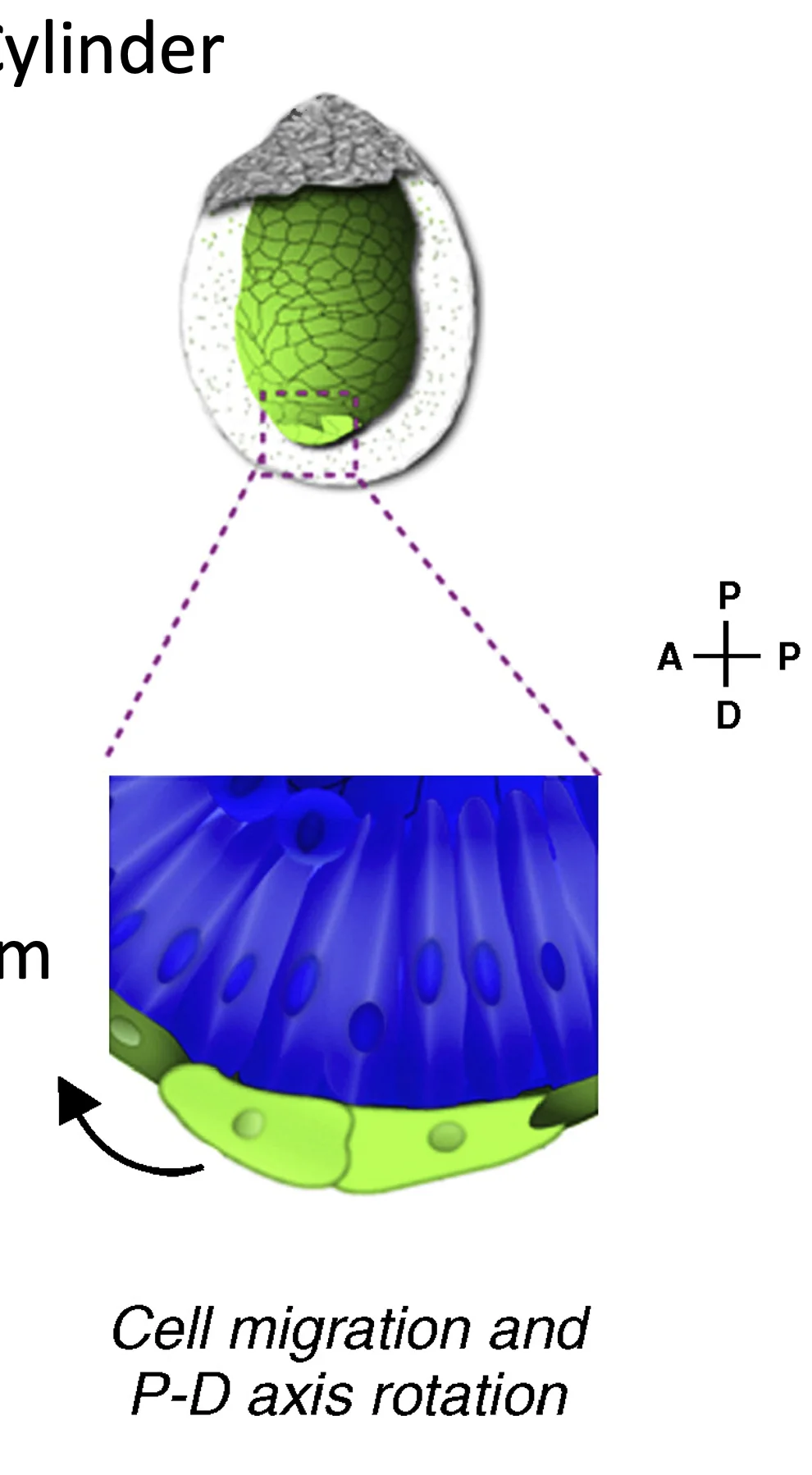
הנדידה של תאי ה־DVE מהקצה הדיסטלי לכיוון פרוקסימלי והפיכתם ל־AVE נקראת Proximal-Distal axis rotation. הציר שהיה קודם לכן Proximal-Distal “מסתובב” והופך לציר ה־Anterior-Posterior.
BEFORE EVENT AFTER
P-D axis only DVE migrates A-P axis is set
Proximal Proximal Proximal
↑ ↑ ↑
┌─────┐ ┌─────┐ ┌─────┐
│ ExE │ AVE │ ExE │ │ ExE │
├─────┤ stops →├─────┤ ANTERIOR←├─────┤→POSTERIOR
│ EPI │ here │ EPI │ │ EPI │
│ │ │ │ │ │
└──●──┘ └──↖──┘ └─────┘
↓ DVE moves
Distal from distal
בשלב שבו תאי ה־DVE נמצאים בקצה הדיסטלי, אי אפשר לדעת לפי המיקום בלבד לאיזה כיוון הם ינדדו. הקצה הדיסטלי הוא נקודה עם סימטריה רדיאלית: אפשר לנוע ממנו לכיוונים שונים. אחרי שהתאים נודדים ונעצרים, המקום שאליו הגיעו הופך ל־Anterior.
RNA in situ hybridization וביטוי Nodal
מה בודקים ב־RNA in situ hybridization?
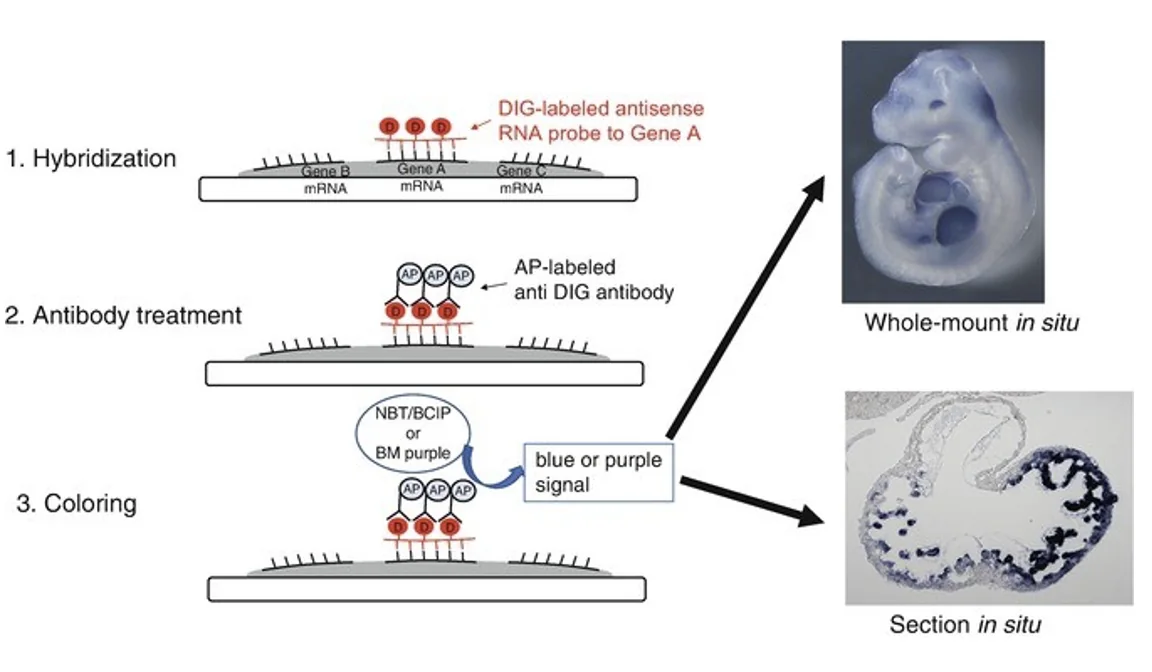
RNA in situ hybridization (ISH) היא שיטה שמראה איפה גן מסוים מתבטא בעובר שלם או ברקמה. היא מזהה את ה־mRNA של הגן, וכך מאפשרת לראות באילו תאים מתרחש שעתוק שלו.
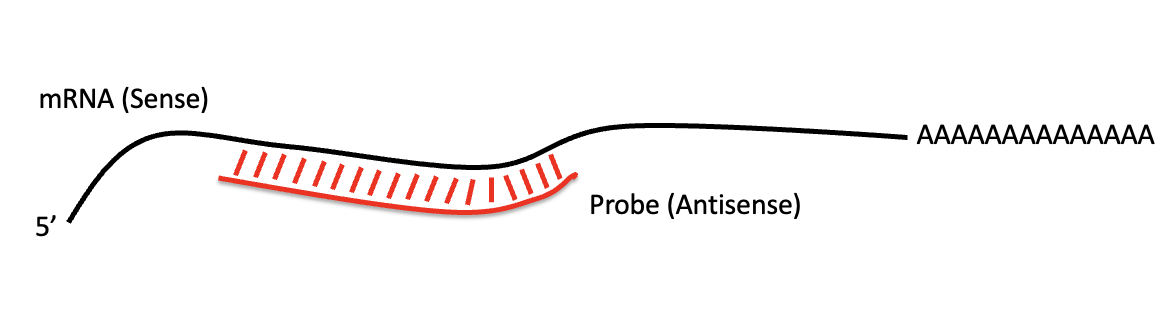
העיקרון:
- מייצרים RNA probe קצר, בדרך כלל באורך של כמה מאות בסיסים.
- ה־probe הוא antisense, ולכן הוא יכול להיקשר ספציפית ל־mRNA של הגן הרצוי.
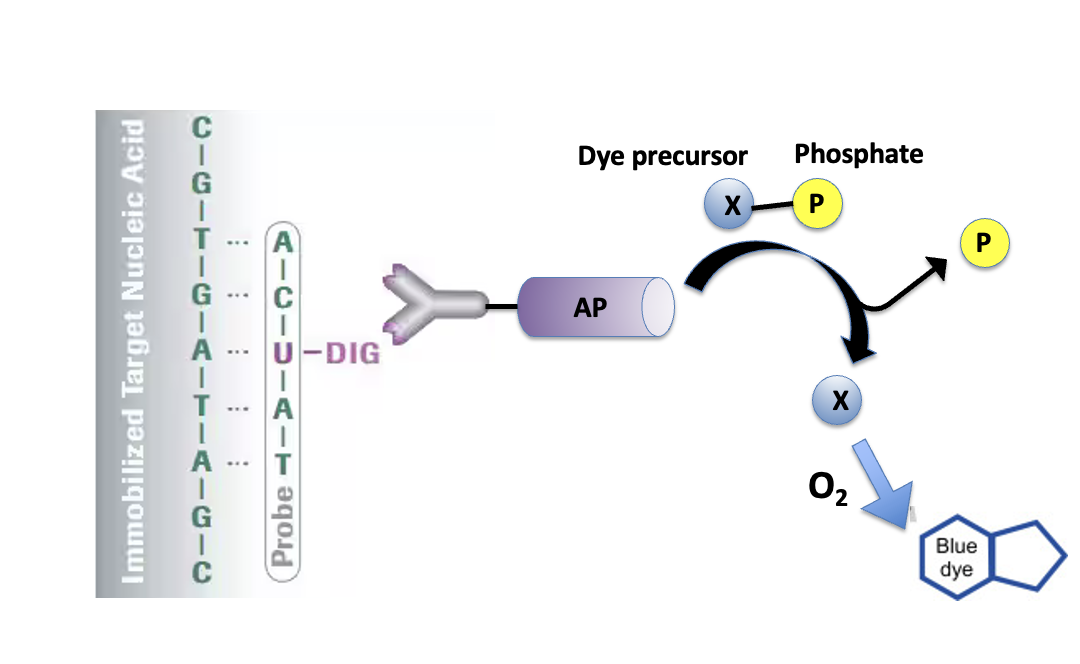
- מסמנים את ה־probe במולקולה שניתן לזהות, למשל DIG.
- מכניסים את ה־probe לעובר מקובע.
- איפה שה־probe נקשר ל־RNA המטרה, מתקבל אות צבעוני.
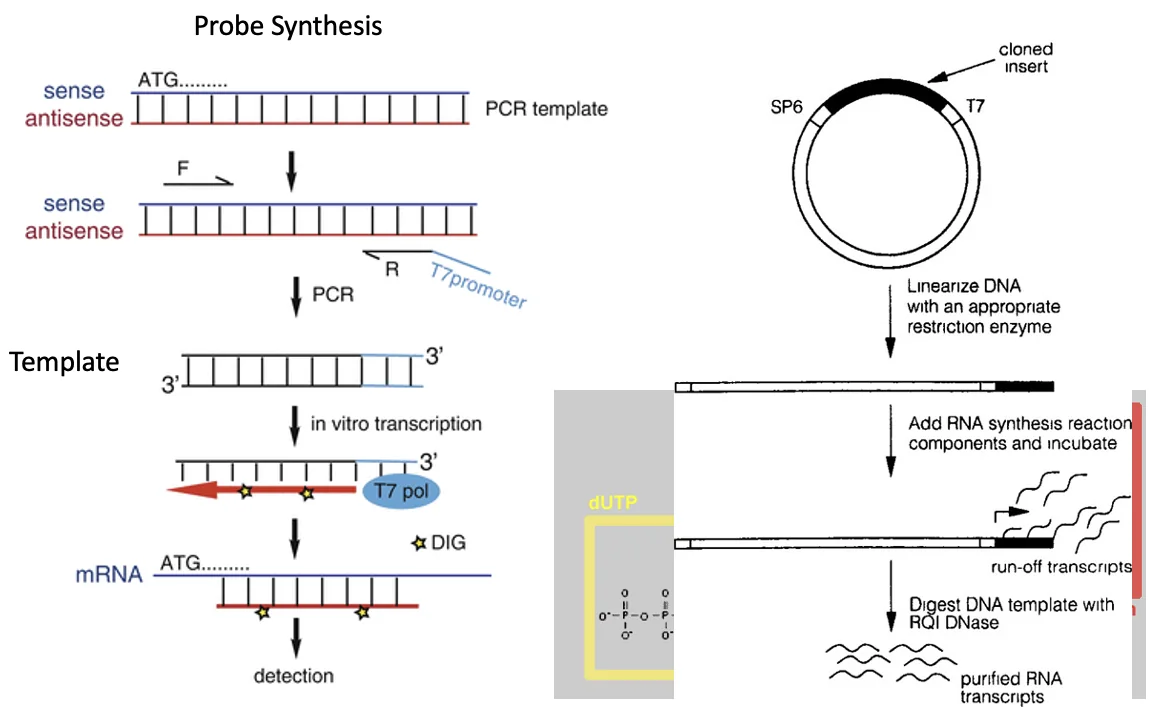
אחת הדרכים לייצר את ה־probe היא להשתמש בפריימר שמכיל רצף של T7 promoter. אחרי PCR מתקבל מקטע DNA עם T7 promoter, ואז משתמשים ב־T7 polymerase כדי לבצע in vitro transcription ולייצר ממנו RNA מסומן.
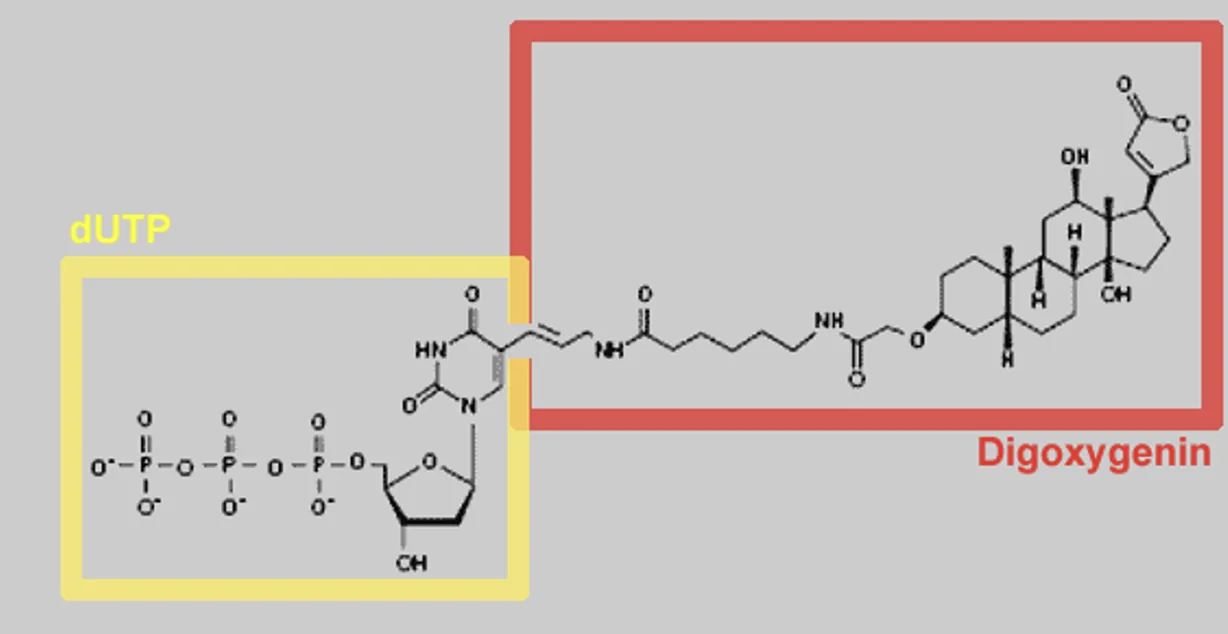
במהלך הסינתזה משלבים נוקלאוטיד מסומן, למשל UTP שמחובר אליו DIG. לאחר מכן משתמשים בנוגדן שמזהה DIG. הנוגדן מחובר לאנזים, למשל Alkaline phosphatase, שממיר סובסטרט מסיס לתוצר כחול ובלתי מסיס.
ב־whole mount in situ hybridization עובדים עם העובר השלם. צריך לקבע (פיקסציה) ולחורר אותו כדי שה־probe ייכנס לתאים. העובר לא נשאר בחיים לאחר הפעולה. האזור הכחול בתוצאה מסמן את המקום שבו ה־RNA של הגן מתבטא.
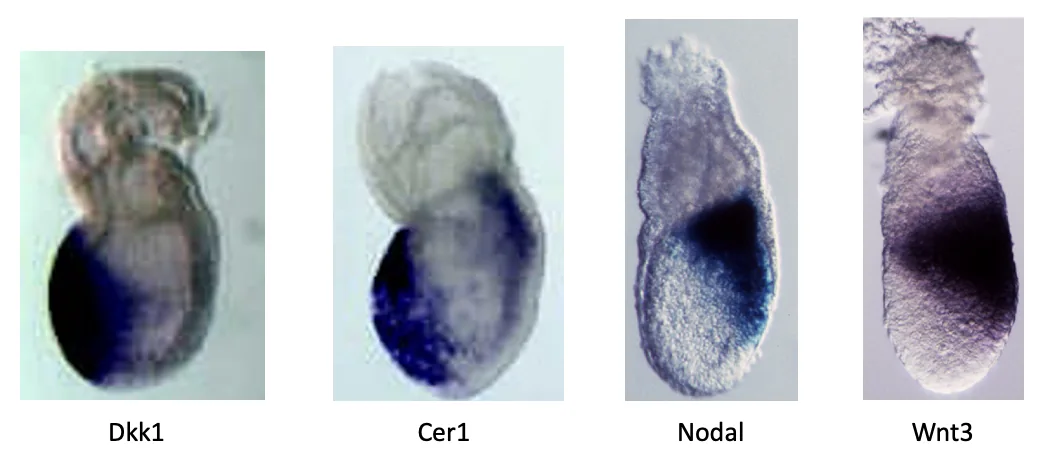
ביטוי Nodal ב־Egg cylinder
בדוגמה הראשונה משתמשים ב־ISH כדי לבדוק את הביטוי של Nodal.
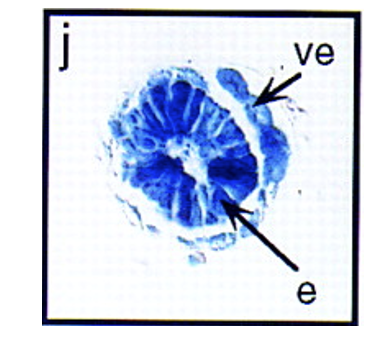
ב־whole mount רואים ש־Nodal מתבטא בצד העוברי, ולא ב־ExE (הצד הימני של התמונה למטה). כדי להבין טוב יותר באילו תאים הביטוי נמצא, עושים חתך (צד שמאל של התמונה). בחתך רואים ביטוי גבוה באפיבלסט, וגם ביטוי ב־VE.
 |  |
Nodal כחלק ממשפחת TGFβ
Nodal שייך ל־TGFβ superfamily. בעץ הפילוגנטי הוא נמצא באזור שקרוב יותר לענף של Activin מאשר ל־BMP.
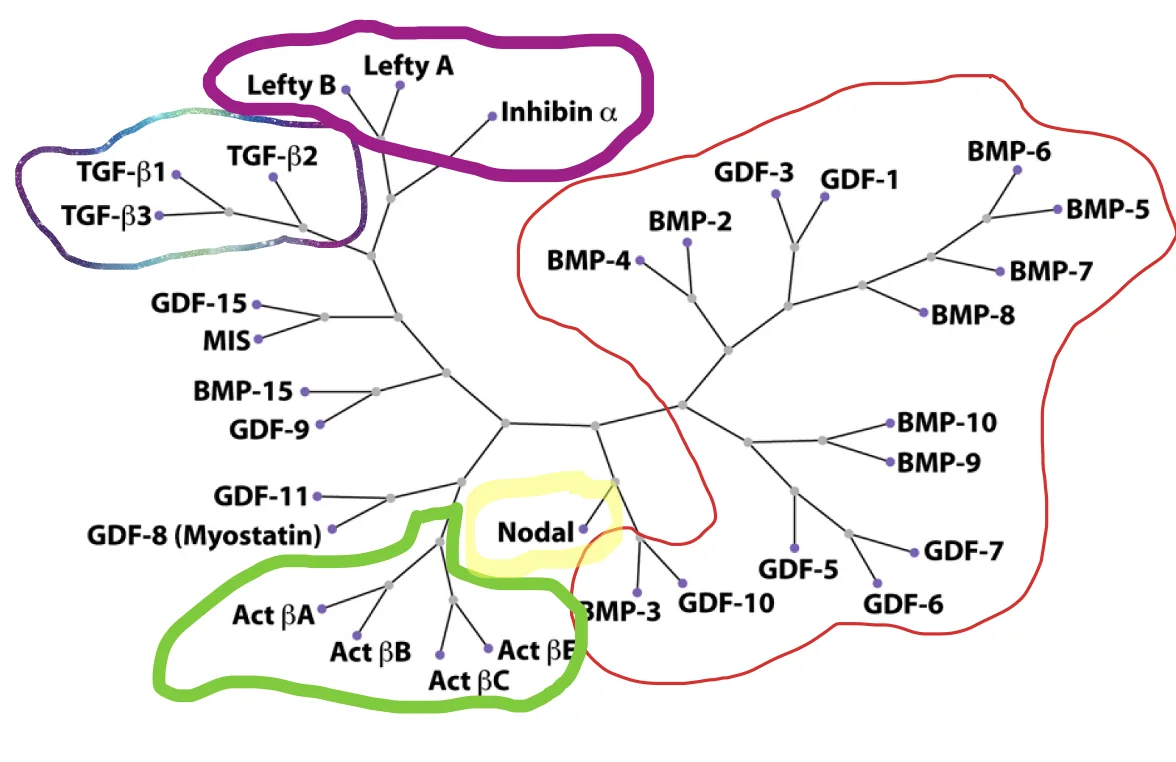
המשמעות המעשית היא ש־Nodal משתמש ברצפטורים של מסלול Activin/Nodal:
| ליגנד | רצפטורים | Smads עיקריים |
|---|---|---|
| BMP / GDF | Type I + Type II של BMP | Smad1/5/8 |
| Activin / Nodal | Type I + Type II של Activin/Nodal | Smad2/3 |
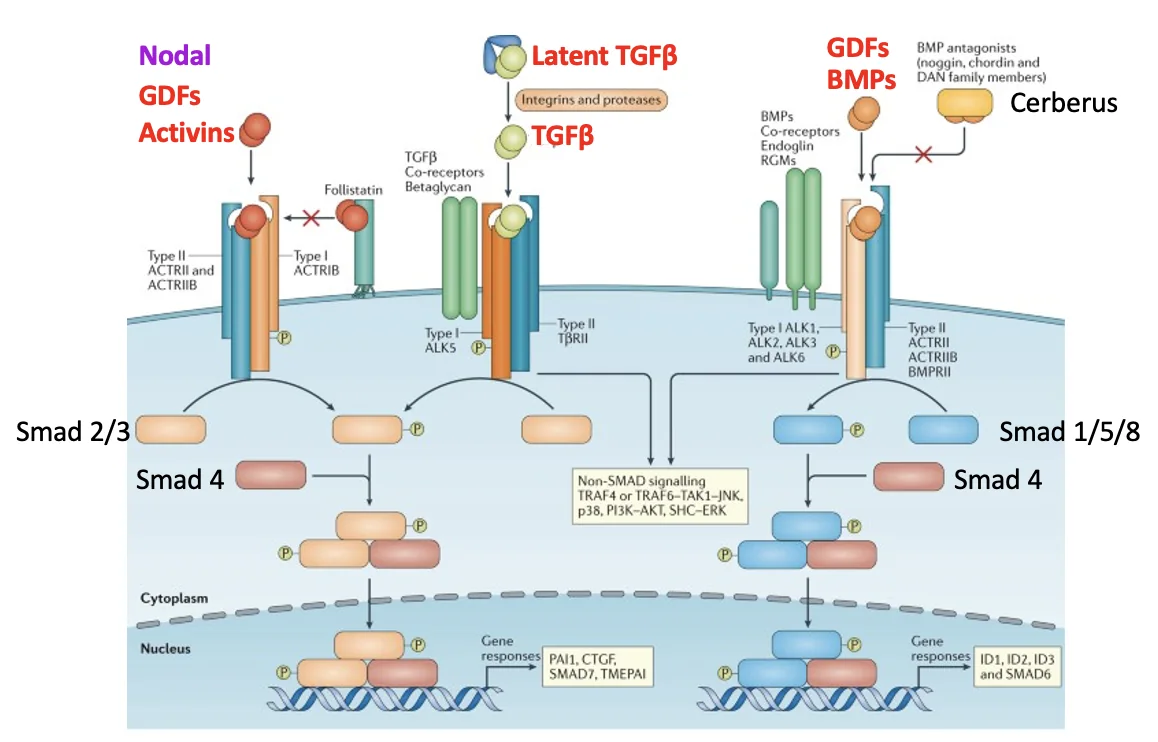
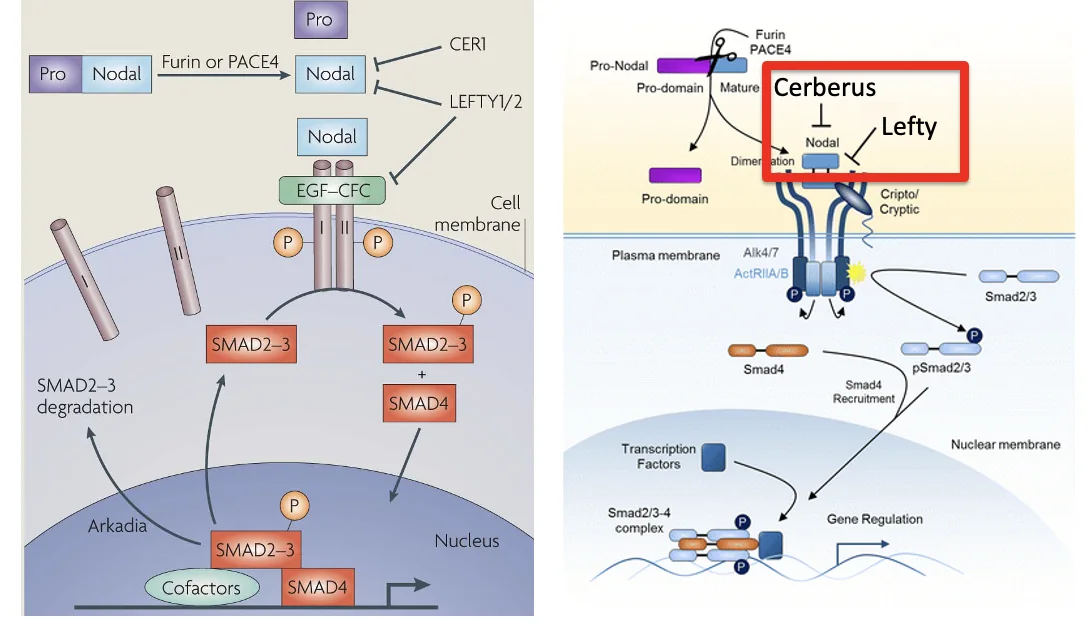
בניגוד לאקטיבינים, Nodal צריך גם co-receptor בשם Cripto בשביל להפעיל את הרצפטורים בצורה תקינה.
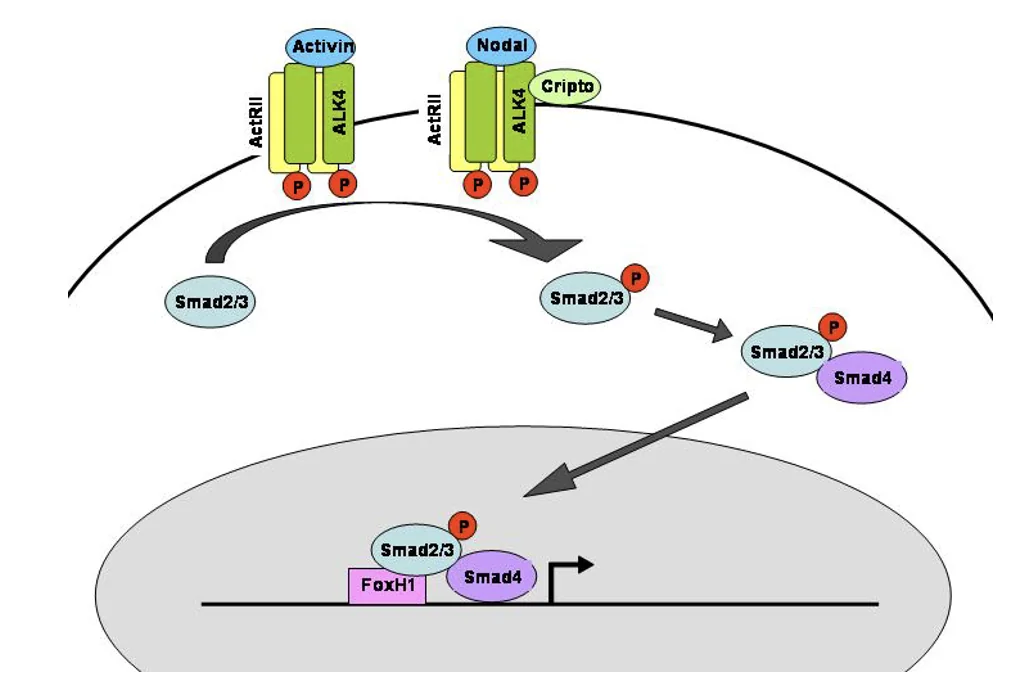
Activin/Nodal signaling ו־Cripto
כאמור, ב־Activin signaling, הליגנד יכול להיקשר ישירות לרצפטורי Type I ו־Type II. ב־Nodal signaling צריך בנוסף את Cripto.
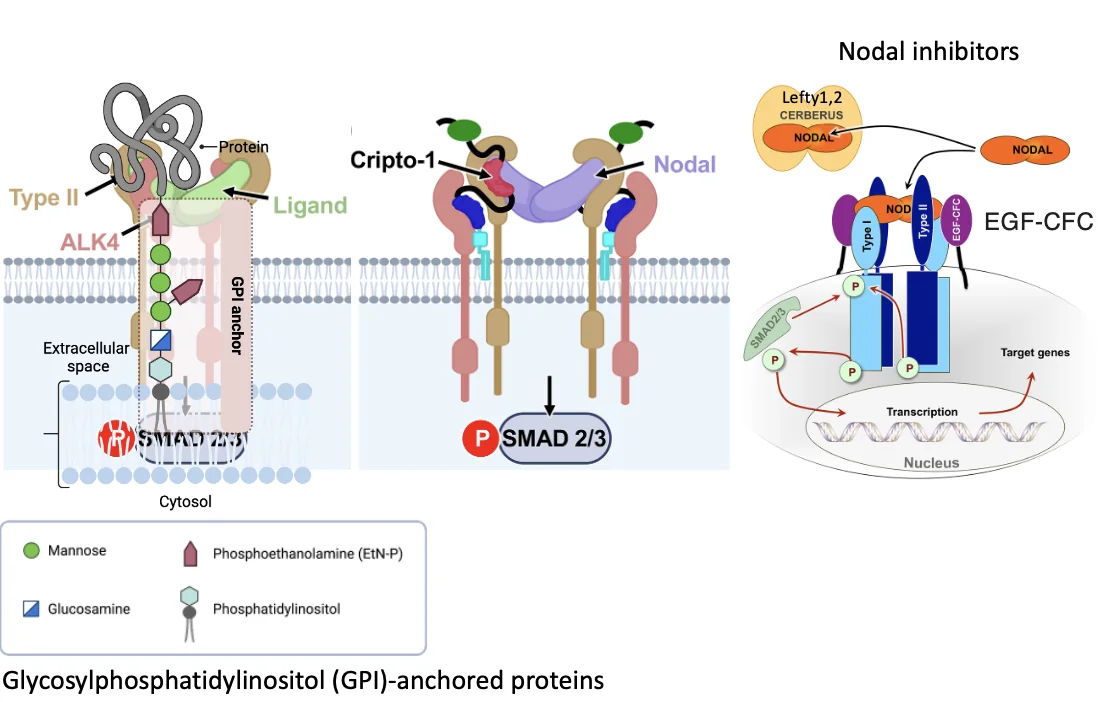
Cripto הוא חלבון שמחובר לממברנה מבחוץ דרך עוגן GPI. הוא לא חוצה את הממברנה ולא מעביר אות בעצמו. תפקידו לאפשר ל־Nodal להפעיל את הקומפלקס הרצפטורי.
למסלול Nodal יש גם מעכבים:
| מעכב | פעולה |
|---|---|
| Lefty1/2 | ממשפחת TGFB; נקשרים ל־Nodal ומונעים ממנו להפעיל את הרצפטורים |
| Cerberus | נקשר ל־Nodal ומונע את פעילותו; בהמשך השיעור הוא מתואר גם כאנטגוניסט רחב יותר |
Lefty מלשון Left-right determination factor.
עיבוד הליגנד Nodal
ליגנדים ממשפחת TGFβ מיוצרים לרוב כ־Pro-ligands: חלבון גדול שמכיל pro-domain וצורה בוגרת של הליגנד. כדי לקבל ליגנד פעיל, נדרש ביקוע על ידי קונברטאזות (convertases).
ברוב בני המשפחה, ה־Pro-ligand לא פעיל. Nodal יוצא דופן: גם ה־Pro-Nodal יכול להפעיל את המסלול, אבל בעוצמה נמוכה.
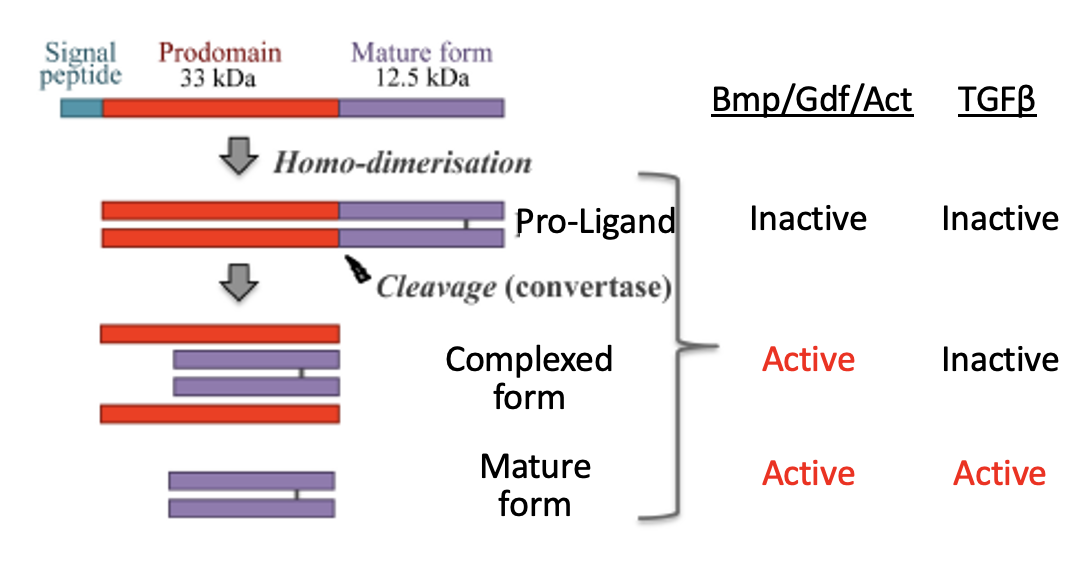 | 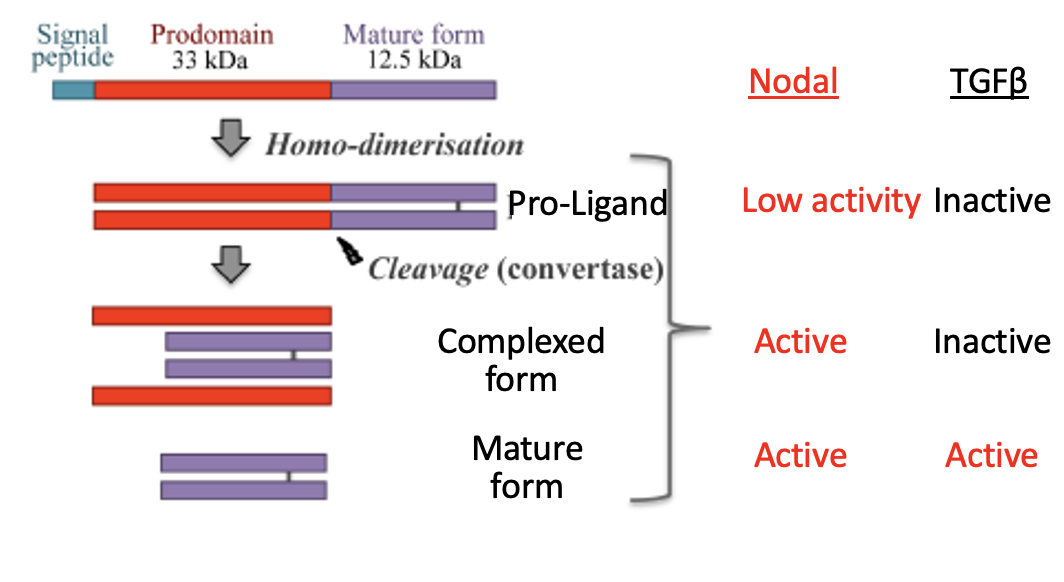 |
הקונברטאזות החשובים כאן הם Furin ו־PACE4. הם בדרך כלל פועלים בתוך התא, למשל באזור ה־ER/Golgi, והם אינם ספציפיים רק ל־Nodal. בסיטואציה המתוארת בשיעור הם יכולים גם להיות מופרשים ולפעול מחוץ לתא.
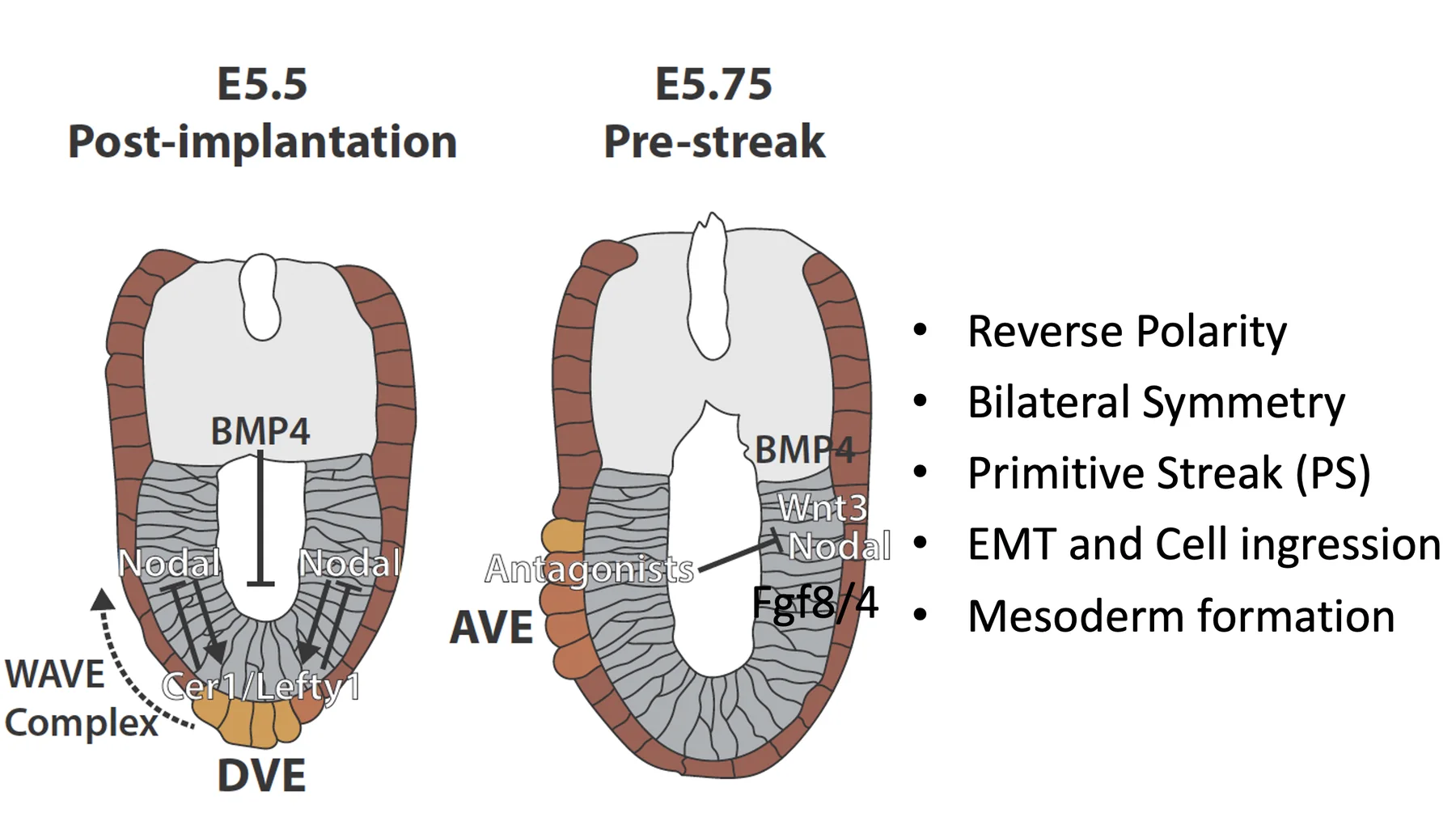
Nodal, BMP, Wnt והיווצרות DVE/AVE
Pro-Nodal באפיבלסט
האפיבלסט מבטא Nodal, אבל הוא לא מבטא Furin ו־PACE4 (הקונברטאזות). לכן הוא מפריש בעיקר Pro-Nodal, עם פעילות נמוכה.
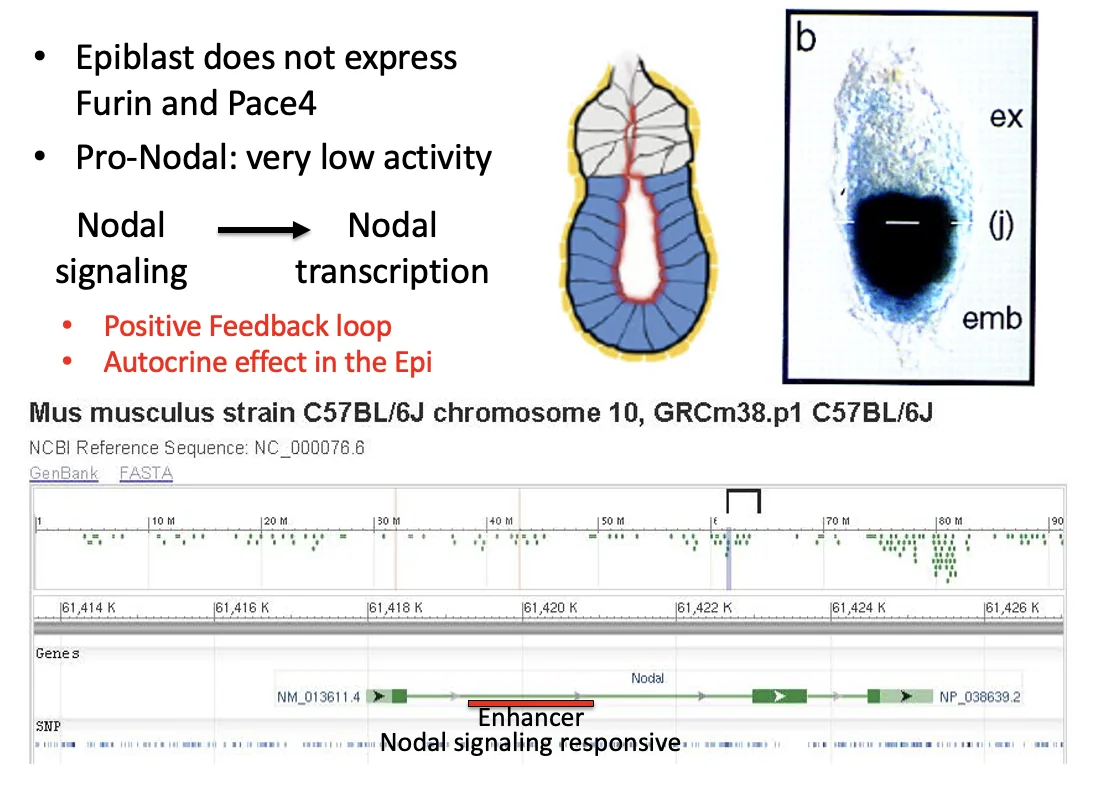
בעובר קיימים רצפטורים ל־Nodal. לכן גם Pro-Nodal יכול להפעיל את המסלול (ברמה נמוכה). בתוך הגן של Nodal יש enhancer שמגיב ל־Nodal signaling. ברגע שהמסלול מופעל, הוא מגביר שעתוק של Nodal עצמו.
זה יוצר positive feedback loop:
Pro-Nodal מהאפיבלסט
↓ פעילות נמוכה של Nodal signaling
הפעלת enhancer של Nodal
↓
עוד שעתוק של Nodal
↓
עוד Pro-Nodal
כך האפיבלסט יכול לשמור על ביטוי Nodal.
תגובת ה־ExE: ה־BMP4
Pro-Nodal פועל גם על תאים מסביב. כאשר ה־ExE נחשף לרמות נמוכות של Pro-Nodal, הוא מתחיל לשעתק Bmp4.
BMP4 מתבטא ב־ExE ולא באפיבלסט. החלבון BMP4 מופרש ומתפשט לכיוון האפיבלסט, ולכן נוצר גרדיאנט על ציר ה־Proximal-Distal: ריכוז גבוה יותר באזור הפרוקסימלי, וריכוז נמוך יותר בקצה הדיסטלי (ה־DVE).
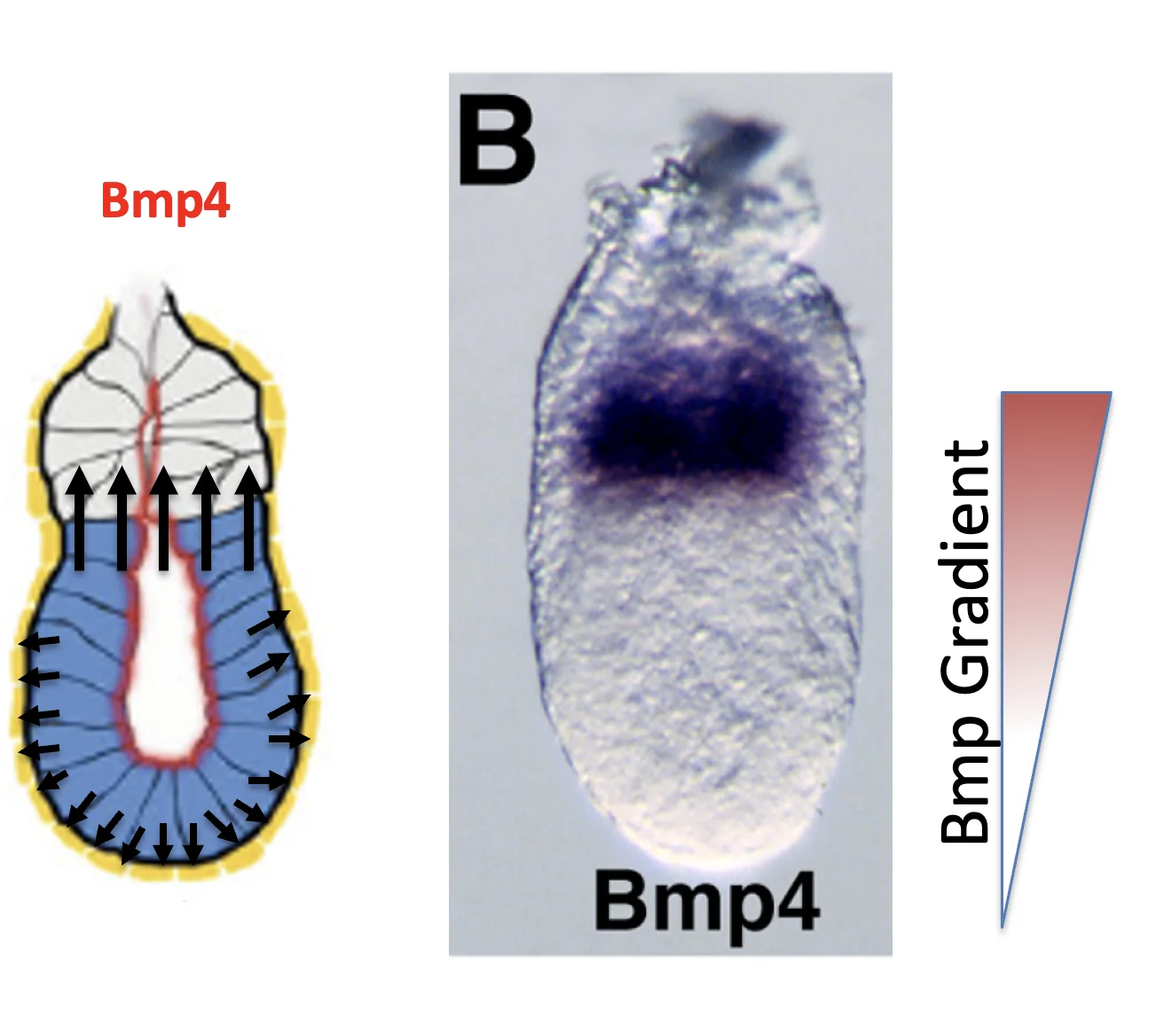
Nodal activity gradient
ה־ExE מגיב ל־Nodal signaling גם באמצעות ביטוי והפרשה של Furin ו־PACE4. הקונברטאזות האלה מבקעים את Pro-Nodal והופכים אותו ל־Nodal פעיל יותר.
כך נוצר גרדיאנט נוסף, הפעם של פעילות Nodal ולא של Nodal RNA. ה־RNA של Nodal עדיין נמצא בעיקר באפיבלסט, אבל הפעילות של Nodal גבוהה יותר בצד הפרוקסימלי, קרוב ל־ExE, ונמוכה יותר בצד הדיסטלי.
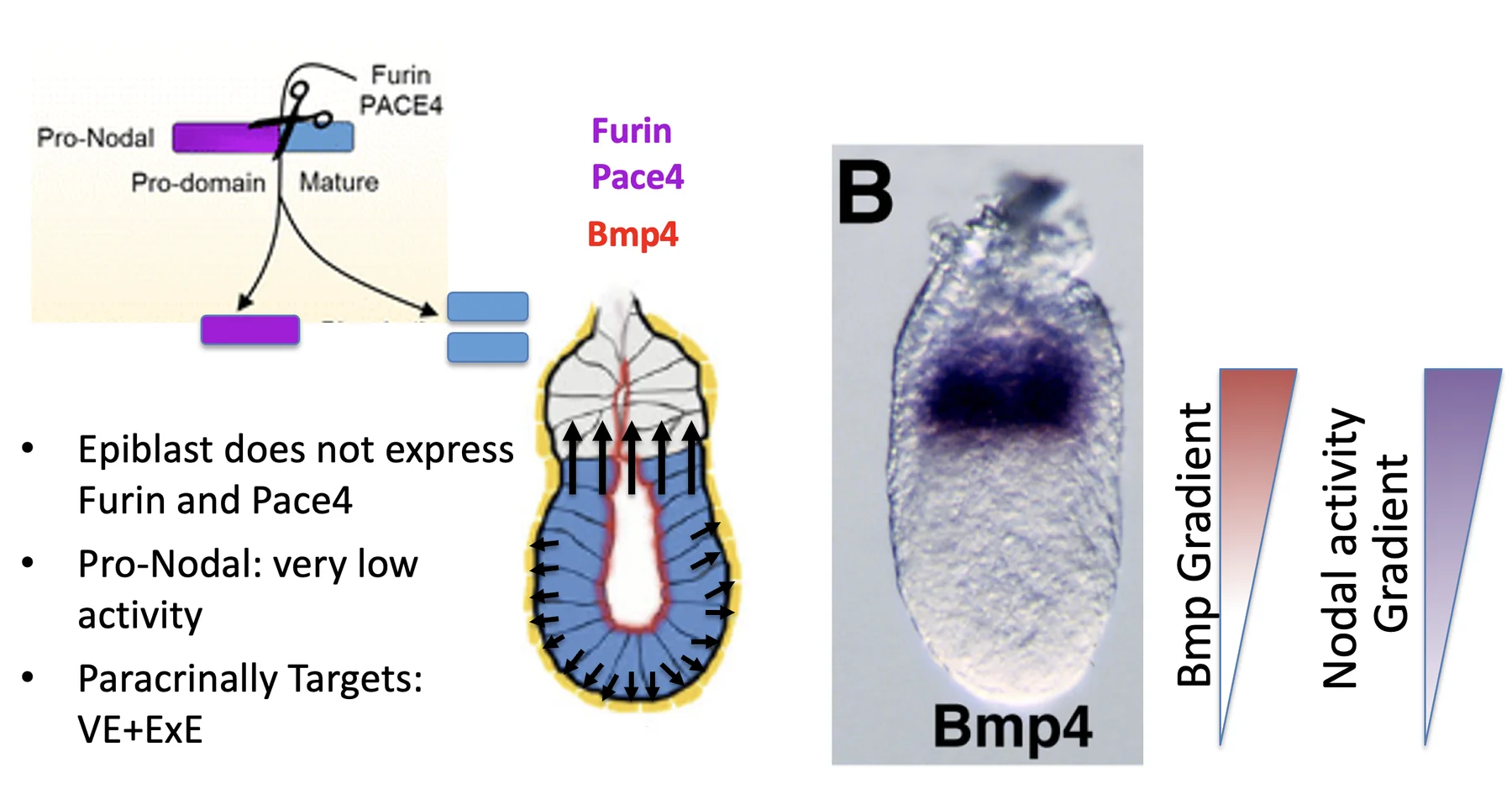
Wnt3 gradient
BMP signaling באפיבלסט גורם לשעתוק של Wnt3. מכיוון ש־BMP מופיע בגרדיאנט, גם Wnt3 נוצר בהדרגה לאורך אותו כיוון.
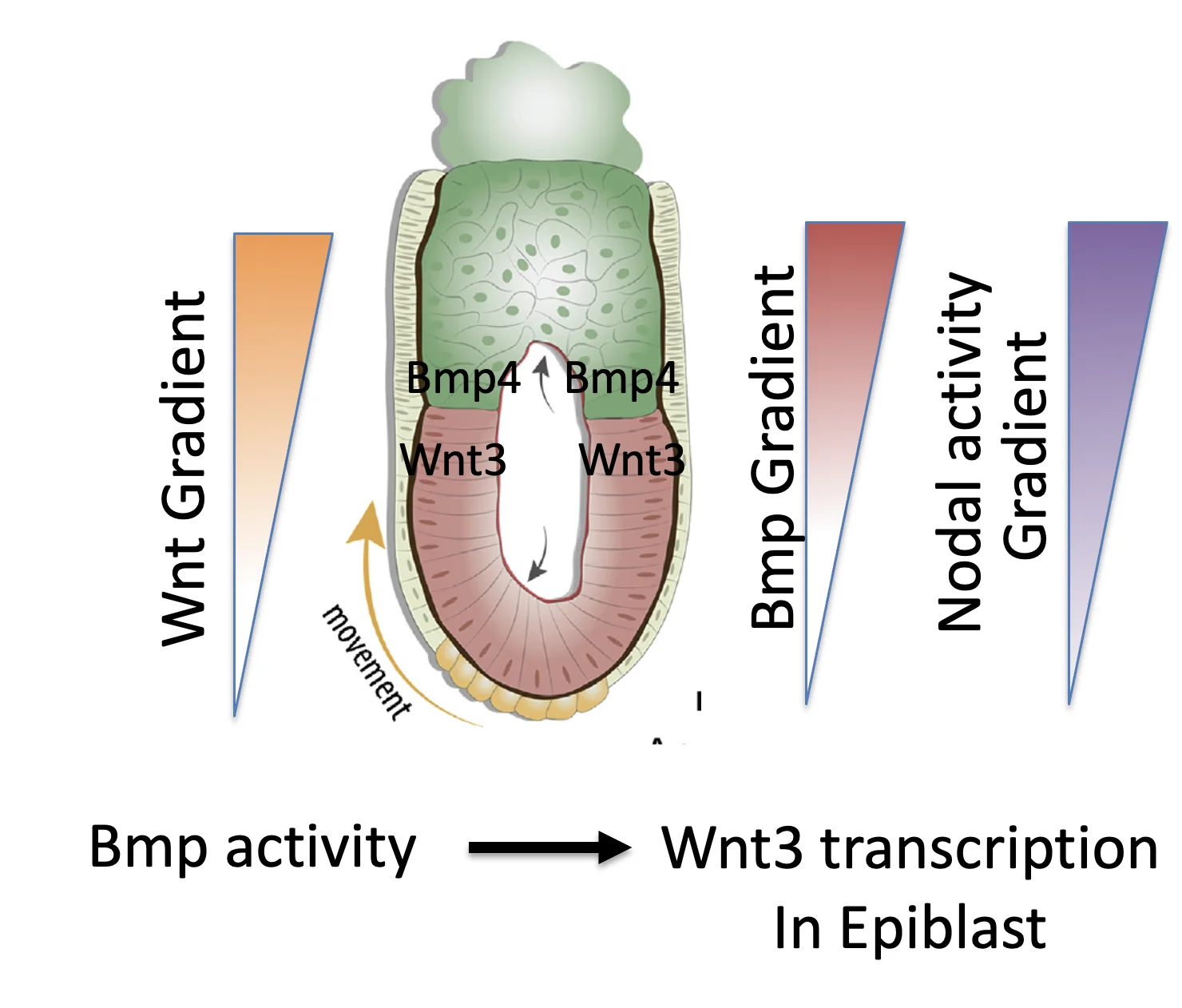
בשלב הזה קיימים שלושה גרדיאנטים בכיוון Proximal-Distal:
| גרדיאנט | מקור / מנגנון |
|---|---|
| BMP4 | ExE מייצר BMP4 בתגובה ל־Pro-Nodal |
| Nodal activity | ExE מייצר/מפריש Furin ו־PACE4 שמפעילים Pro-Nodal (הופכים אותו ל־Nodal בוגר ופעיל יותר) |
| Wnt3 | BMP signaling באפיבלסט גורם לשעתוק Wnt3 |
יצירת DVE בקצה הדיסטלי
כדי ליצור DVE צריך שילוב מסוים:
| תנאי | משמעות |
|---|---|
| פעילות של Nodal | דרושה לאינדוקציה של DVE |
| רמת BMP נמוכה מאוד | מאפשרת יצירת DVE (BMP מעכב יצירת DVE) |
השילוב הזה מופיע בקצה הדיסטלי: יש שם מעט פעילות של Nodal (דרך Pro-Nodal), ורמות BMP נמוכות מאוד. לכן ה־DVE נוצר דווקא בנקודה הדיסטלית.
לאחר שנוצר, ה־DVE מתחיל לבטא אנטגוניסטים:
| גן / חלבון | תפקיד |
|---|---|
| Lefty1 | מעכב Nodal |
| Cerberus1 / Cer1 | אנטגוניסט של Nodal; בהמשך מתואר גם כמעכב של BMP ו־Wnt |
| Dkk1 | אנטגוניסט של Wnt signaling |
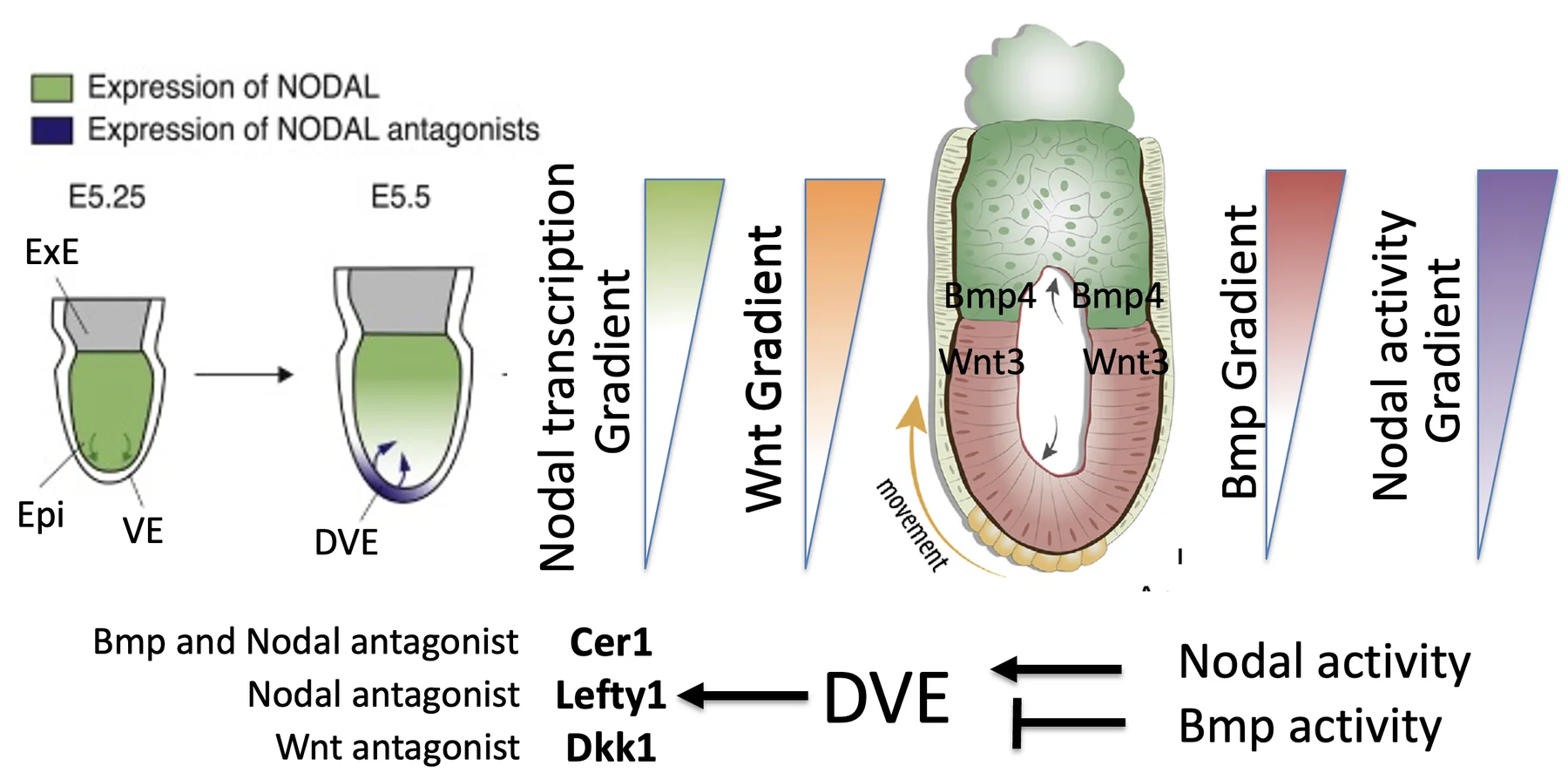
האנטגוניסטים שמופרשים מה־DVE מתחילים לדחוק את הפעילות של Nodal. בגלל ה־positive feedback של Nodal, כאשר פעילות Nodal נדחקת, גם ביטוי ה־RNA של Nodal נדחק לכיוון הפרוקסימלי.
מ־DVE ל־AVE: הפיכת הציר
תאי ה־DVE נודדים לכיוון פרוקסימלי והופכים ל־AVE. בזמן הנדידה הם ממשיכים לבטא אנטגוניסטים. כך הם קובעים את הצד האנטריורי ומרחיקים את הפעילות של Nodal, BMP ו־Wnt לצד הפוסטריורי.
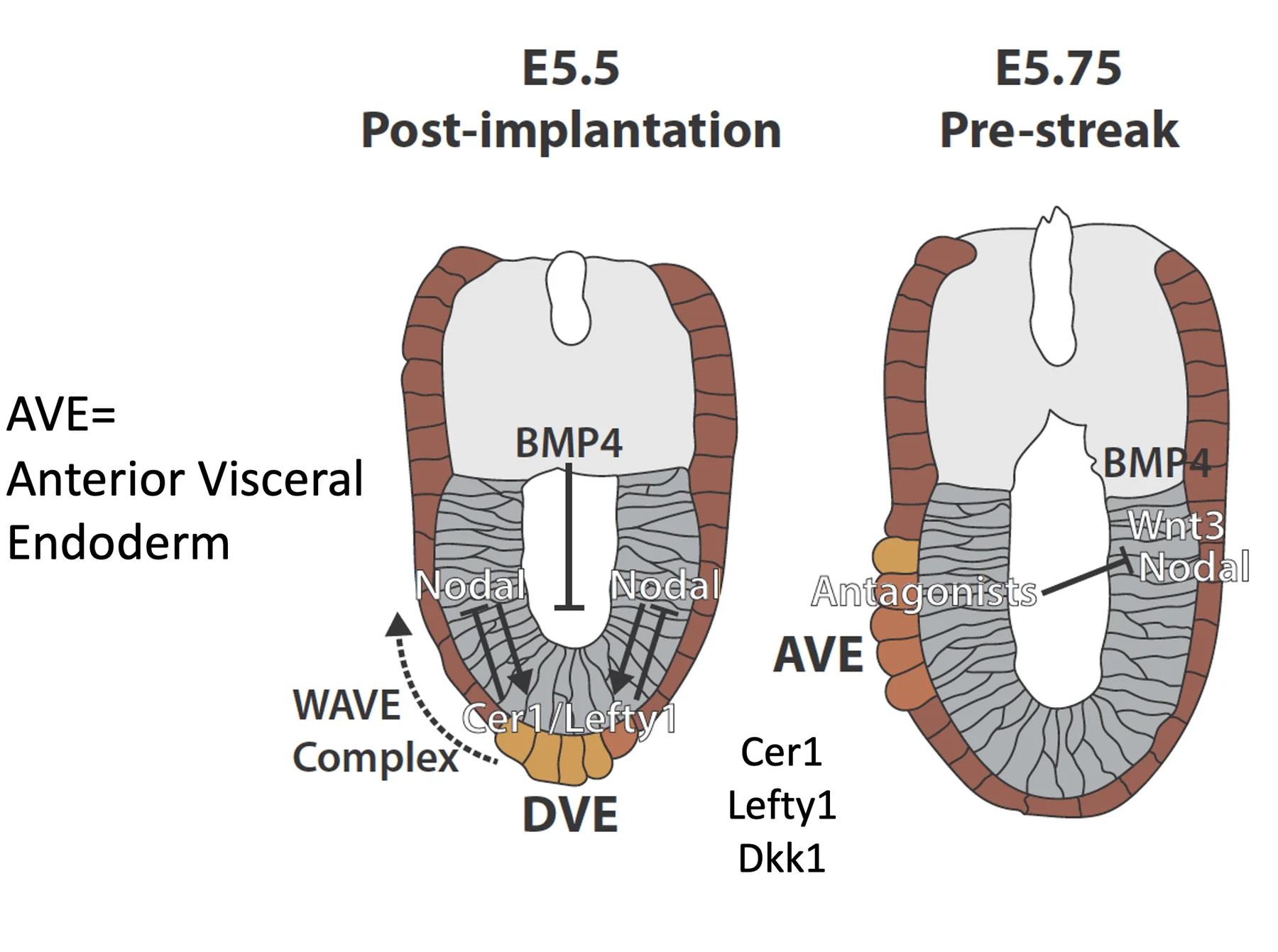
הדחיקה הזאת מייצרת הפרדה ברורה:
| צד | מאפיין |
|---|---|
| Anterior | AVE, ביטוי אנטגוניסטים כמו Dkk1 ו־Cer1 |
| Posterior | רמות גבוהות יותר של Nodal, BMP4 ו־Wnt3 |
 | 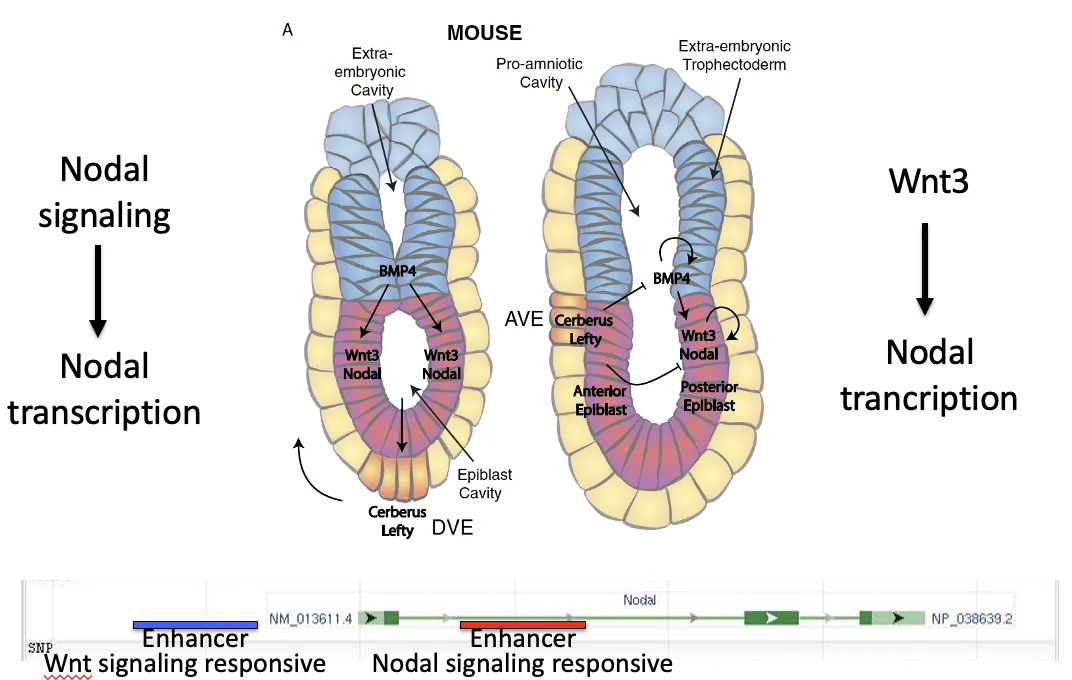 |
יש כאן נקודה חשובה לגבי Nodal: בהתחלה, Nodal שומר על הביטוי שלו דרך enhancer שמגיב ל־Nodal signaling. אחרי שה־AVE נודד ומתקרב לצד הפוסטריורי, האנטגוניסטים שלו עלולים להשתיק את Nodal. לכן נכנס מנגנון נוסף: enhancer של Nodal שמגיב ל־Wnt signaling. כך הביטוי של Nodal נשמר בצד הפוסטריורי גם כשהאזור האנטריורי מפריש אנטגוניסטים.
רצף האירועים:
Nodal באפיבלסט
↓
ExE מייצר BMP4, Furin ו־PACE4
↓
נוצרים גרדיאנטים של BMP, Nodal activity ו־Wnt3 על ציר Proximal-Distal
↓
בקצה הדיסטלי: Nodal activity + BMP נמוך
↓
DVE נוצר
↓
DVE מבטא Lefty1, Cer1 ו־Dkk1
↓
DVE נודד והופך ל־AVE
↓
Anterior מוגדר באזור ה־AVE
↓
Nodal/BMP/Wnt נדחקים לצד Posterior
↓
נוצרים התנאים לתחילת Gastrulation
המצב לפני גסטרולציה
לפני תחילת הגסטרולציה כבר רואים ביטוי מרחבי ברור של הגנים המרכזיים:
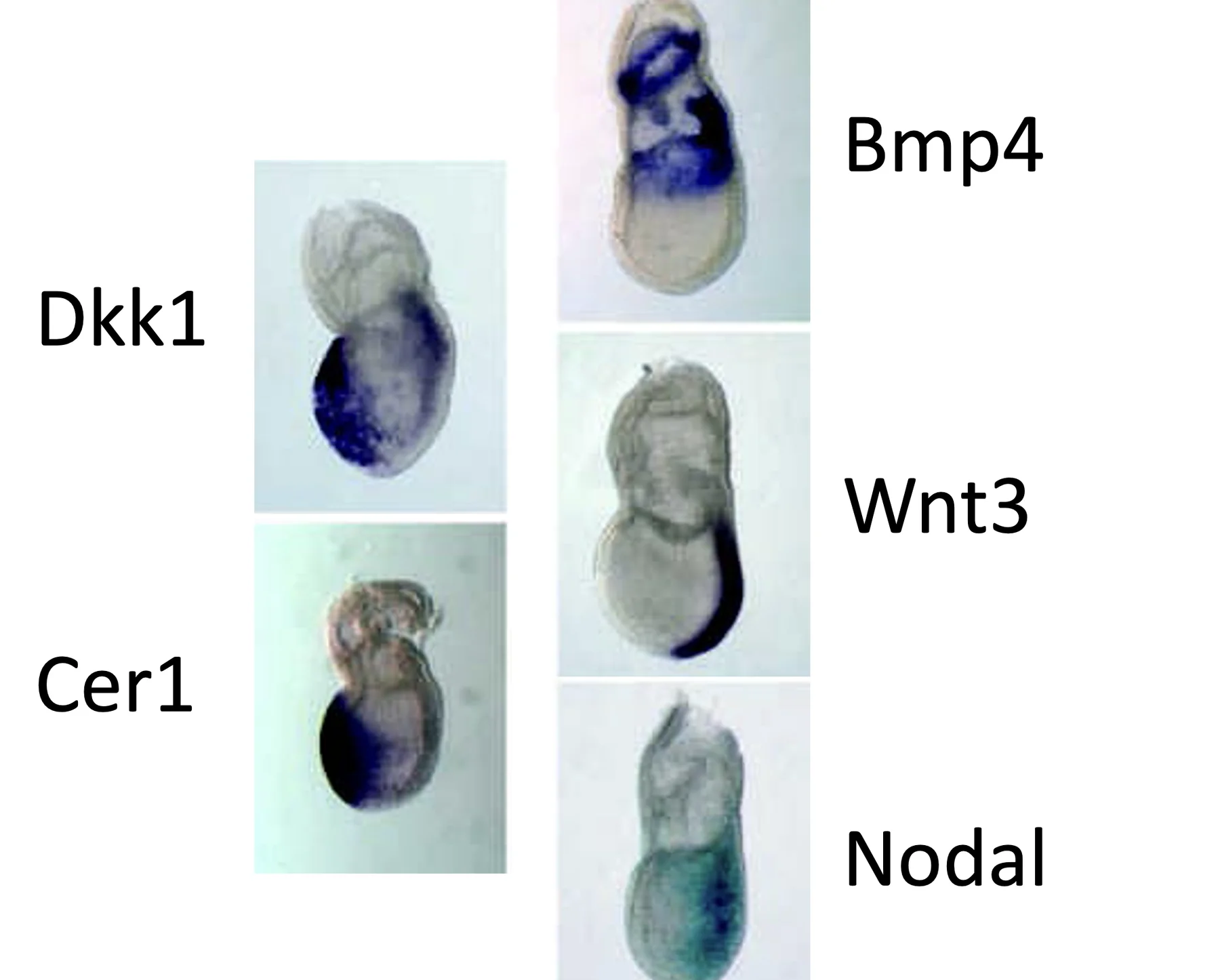
בשלב הזה Dkk1 ו־Cer1 נמצאים בצד, לא בנקודה הדיסטלית. לכן אלה כבר תאי AVE ולא DVE. בצד הפוסטריורי נמצאים Nodal, Wnt3 ו־BMP4, שמכינים את האזור שבו יתחיל ה־Primitive streak.
עכבר לעומת אדם
בעכבר נוצר Egg cylinder עם Epiblast ו־ExE. באדם המבנה שונה: האפיבלסט מסודר כדיסק, ולא כ־cup/egg cylinder. באדם יש גם שכבה שממנה יתפתח ה־Amnion, ומקורה באפיבלסט.
למרות ההבדלים במבנה, העיקרון דומה: יש אזור שמפריש אנטגוניסטים בצד האנטריורי, ויש פעילות Nodal/Wnt בצד הפוסטריורי, ושם מתחילה הגסטרולציה. בפרטים המוקדמים באדם יש פחות ודאות, בעיקר משום שהשלבים שלאחר השרשה קשים יותר למחקר.
לקראת גסטרולציה: אפיתל, EMT ו־Primitive streak
אפיתל ותאים מזנכימליים
לפני שמתחילים את הגסטרולציה צריך להבחין בין תאים אפיתליאליים לתאים מזנכימליים.
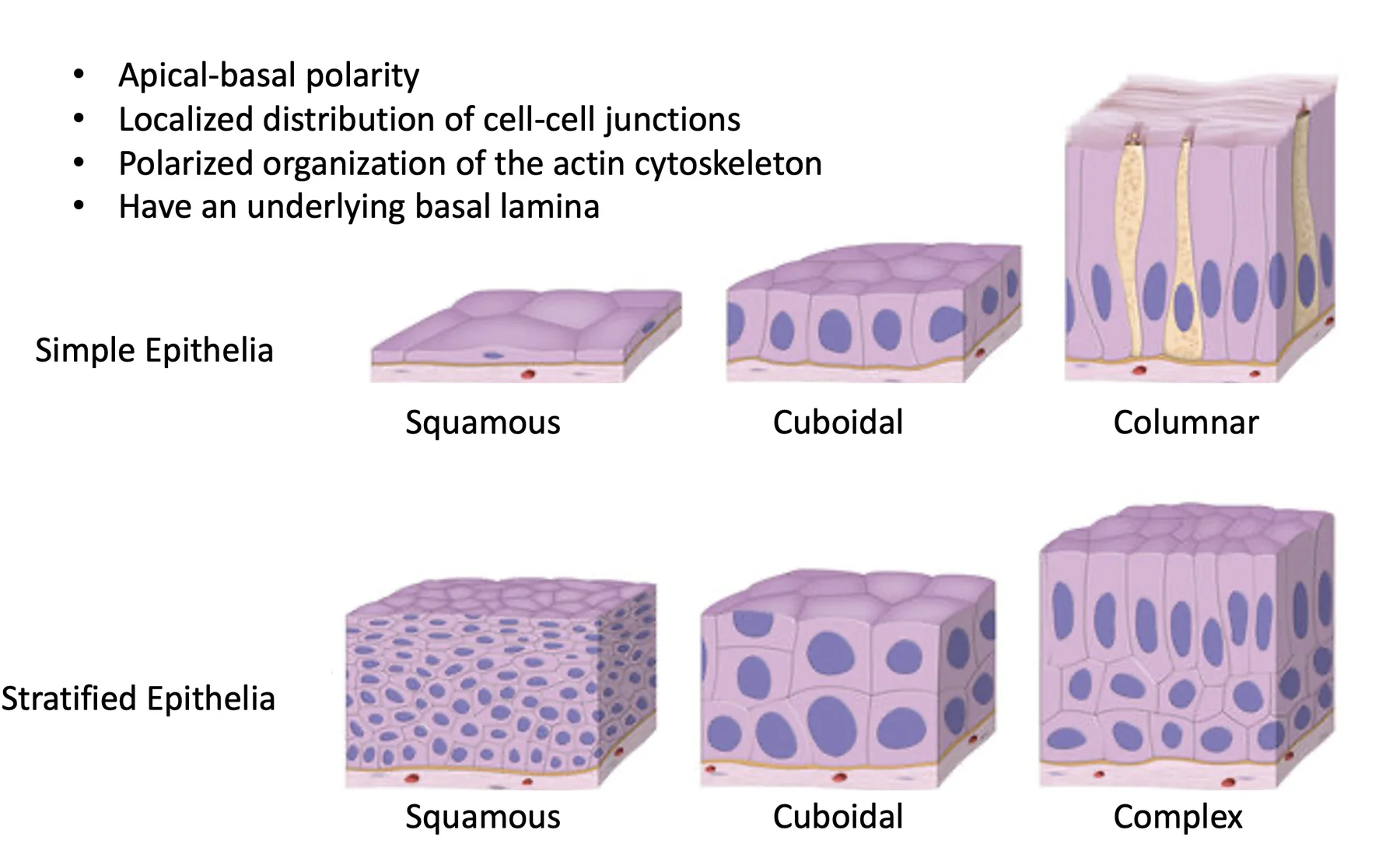
אפיתל הוא שכבת תאים שמפרידה בין שני חללים או שני אזורים. לתאים אפיתליאליים יש כמה מאפיינים קבועים:
- Apical-basal polarity
- סידור מקומי של junctions בין תאים
- ארגון מקוטב של ציטוסקלטון האקטין
- שכבת basal lamina שעליה התאים יושבים
| סוג אפיתל | מאפיין |
|---|---|
| Simple epithelium | שכבת תאים אחת, למשל באזורים רבים במערכת העיכול |
| Stratified epithelium | כמה שכבות תאים, למשל בעור ובחלקים של חלל הפה |
תאים מזנכימליים שונים מתאי אפיתל. אין להם מבנה אפיקלי-בזאלי מסודר, הם מופיעים בצורות מגוונות, והקשרים ביניהם נקודתיים יותר.
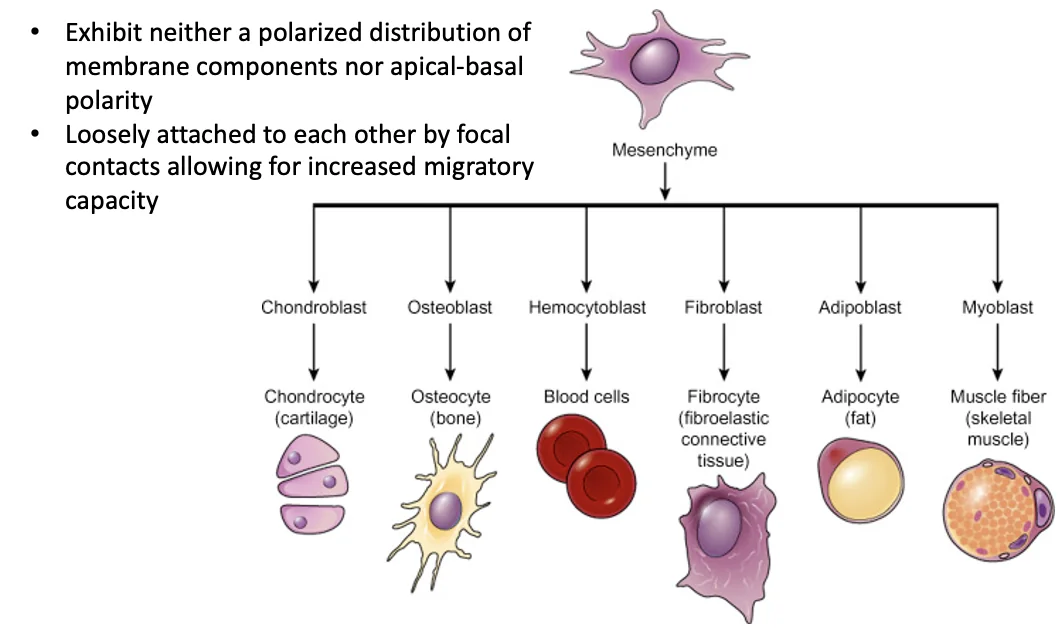
בהקשר העוברי, תאים מזנכימליים הם תאים שמקורם במזודרם. בהמשך הם יכולים לתת מגוון סוגי תאים, למשל תאי סחוס, תאי עצם, תאי דם, פיברובלסטים, תאי שומן ותאי שריר.
EMT ו־MET
שני תהליכים חשובים בגסטרולציה:
| תהליך | פירוש | מה קורה לתאים? |
|---|---|---|
| EMT | Epithelial to Mesenchymal Transition | תא אפיתליאלי הופך לתא מזנכימלי |
| MET | Mesenchymal to Epithelial Transition | תא מזנכימלי הופך לתא אפיתליאלי |
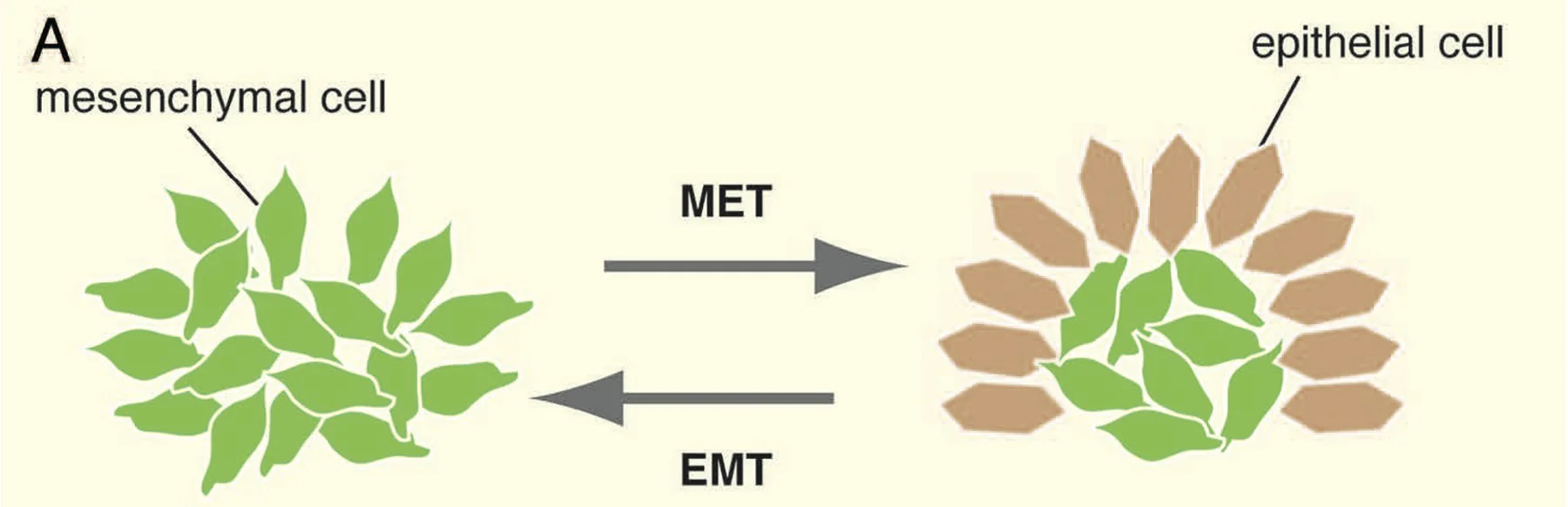
ב־EMT, תא מאבד מאפיינים אפיתליאליים, מקבל מאפיינים מזנכימליים, ויכול לצאת מתוך שכבת האפיתל. היציאה הזאת נקראת ingression. בגסטרולציה זה קריטי, כי תאי אפיבלסט צריכים לעזוב את שכבת האפיבלסט ולהיכנס בין שכבות כדי ליצור מזודרם.
הוזכר שתהליכי EMT מופיעים גם בהקשר סרטני: תאי אפיתל סרטניים יכולים “לרכוב” על תוכנית EMT כדי לפלוש וליצור גרורות. בהקשר של השיעור, החשיבות היא עוברית: EMT הוא מנגנון שמאפשר תנועה והיווצרות של שכבות חדשות.
Reverse polarity
במבנה שלפני גסטרולציה מתקיים מצב של Reverse polarity: הצד הבזאלי של תאי האפיבלסט והצד הבזאלי של ה־VE פונים לאותו אזור (נמצאים זה מול זה).
 \[\begin{array}{c@{\qquad\qquad}ll} \begin{array}{c} \text{Prox.} \\[0.2em] \text{A}\;\;+\;\;\text{P} \\[0.2em] \text{Dist.} \end{array} & \begin{array}{ll} {\color{#6f6f6f}\circ} & \text{Extraembryonic Ectoderm} \\[0.55em] {\color{#bfbfbf}\bullet} & \text{Epiblast} \\[0.55em] {\color{#5a5a5a}\bullet} & \text{Primitive Streak} \\[0.55em] {\color{#8b1220}\bullet} & \text{Visceral Endoderm} \\[0.55em] {\color{#d79a2e}\bullet} & \text{Distal Visceral Endoderm (DVE)} \\[0.55em] {\color{#c95b32}\bullet} & \text{Anterior Visceral Endoderm (AVE)} \end{array} \end{array}\]
\[\begin{array}{c@{\qquad\qquad}ll} \begin{array}{c} \text{Prox.} \\[0.2em] \text{A}\;\;+\;\;\text{P} \\[0.2em] \text{Dist.} \end{array} & \begin{array}{ll} {\color{#6f6f6f}\circ} & \text{Extraembryonic Ectoderm} \\[0.55em] {\color{#bfbfbf}\bullet} & \text{Epiblast} \\[0.55em] {\color{#5a5a5a}\bullet} & \text{Primitive Streak} \\[0.55em] {\color{#8b1220}\bullet} & \text{Visceral Endoderm} \\[0.55em] {\color{#d79a2e}\bullet} & \text{Distal Visceral Endoderm (DVE)} \\[0.55em] {\color{#c95b32}\bullet} & \text{Anterior Visceral Endoderm (AVE)} \end{array} \end{array}\] בשלב הזה העובר כבר עבר מסימטריה רדיאלית לסימטריה בילטרלית (דו־צדדית): יש Anterior ו־Posterior, ויש Dorsal ו־Ventral. ציר ימין-שמאל טרם הוגדר.
תחילת Primitive streak
בצד הפוסטריורי מתרכזים מספר מסלולי איתות:
| מסלול | תפקיד כללי בשלב הזה |
|---|---|
| BMP | משתתף בהכנת האזור הפוסטריורי ובאינדוקציות מוקדמות |
| Wnt3 | שומר על ביטוי Nodal ותומך בתחילת גסטרולציה |
| Nodal | דרוש לתהליכים שמובילים לגסטרולציה |
| FGF signaling | מצטרף למסלולים שתומכים ביצירת Primitive streak ו־EMT |
שילובם מאפשר יצירת Primitive streak בצד הפוסטריורי של האפיבלסט.
Primitive streak הוא אירוע של האפיבלסט: תאי אפיבלסט באזור הפוסטריורי מתחילים לעבור EMT, עושים ingression, ויוצאים מתוך שכבת האפיבלסט. לאחר היציאה הם נמצאים בין האפיבלסט לבין ה־VE. התאים האלה הם תחילת יצירת ה־Mesoderm.
להלן מספר איורים שממחישים את התהליך (קרדיט לא.ס.):
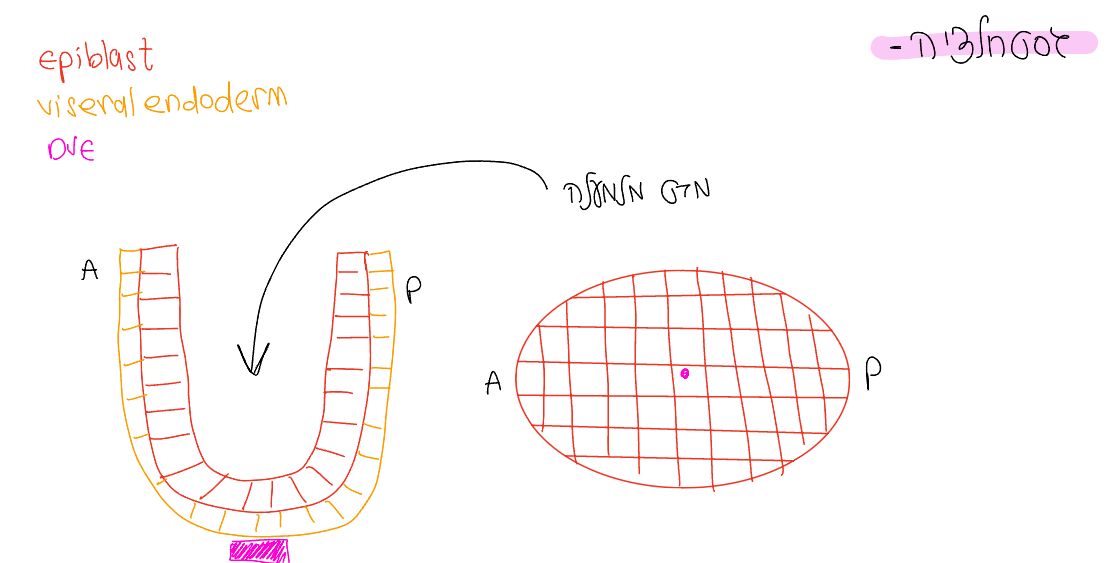
תשומת הלב שבשיעור החלק הצהוב הוצג כ־Visceral ectoderm, לעניות דעתי מדובר ב־Visceral endoderm.
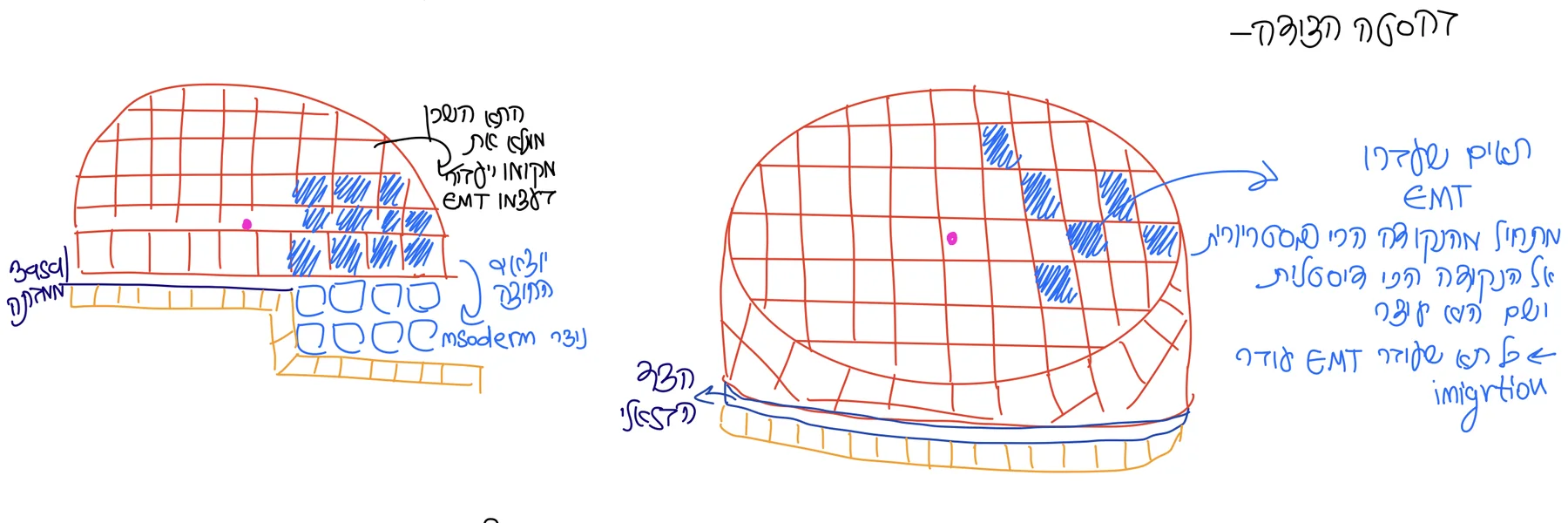
בזמן שתאים יוצאים מהאפיבלסט, לא נשאר “חור” בשכבה: תאים שכנים נכנסים למקום שהתפנה, והתאים ממשיכים להתחלק. כך נשמרת רציפות השכבה בזמן שתאים חדשים עוברים EMT ו־ingression.
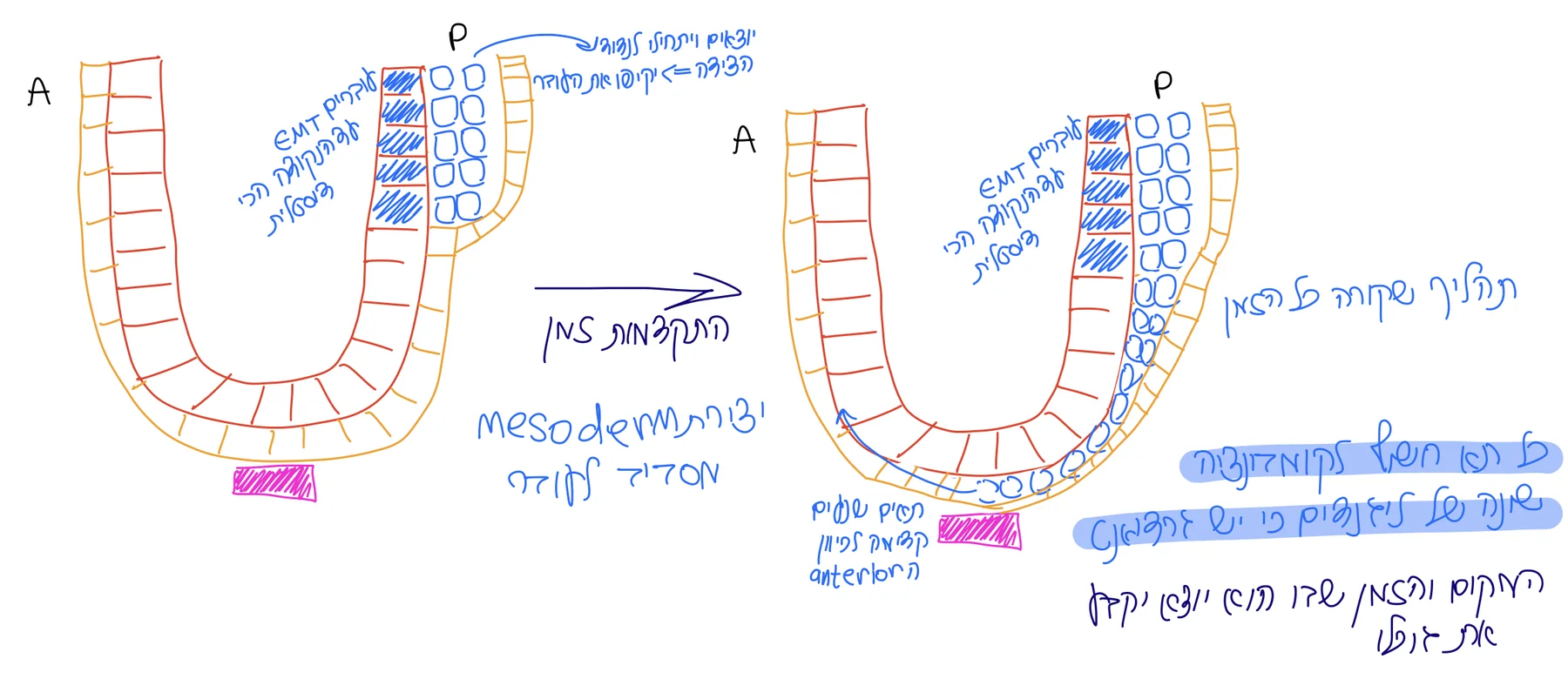
ה־Primitive streak מתחיל בנקודה הפוסטריורית ומתקדם לכיוון הדיסטלי. תאים שיוצאים דרך ה־streak נודדים לצדדים ובין השכבות, כך שהם מקיפים בהדרגה את העובר.
המקום והזמן שבהם תא יוצא דרך ה־Primitive streak משפיעים על גורלו:
| תאים שיוצאים | גורל |
|---|---|
| תאים ראשונים | PGCs - ה־primordial germ cells; הם נעים מהר לאזור החוץ־עוברי כדי להיחשף ל־BMP |
| תאים שיוצאים אחריהם באזור מתאים | תאים שיתרמו ל־cardiac fields וינועו קדימה לכיוון האזור שבו יתפתח הלב |
| תאים שיוצאים מאוחר יותר / ממיקום אחר | גורלות נוספים שיוסברו בהמשך הגסטרולציה |
בשלב הזה הגורל כבר נקבע בצורה הרבה יותר דטרמיניסטית מאשר בשלבי הקליבג׳. הסיבה היא שהתאים עוברים דרך אזורים עם שילובים שונים של Nodal, BMP, Wnt ו־FGF, וכל תא נחשף לסביבה מולקולרית אחרת לפי המיקום והזמן שבו הוא יוצא.
גם כאן העיקרון דומה בין עכבר לאדם. בעכבר קל יותר לראות את זה ב־Egg cylinder, ובאדם באפיבלסט הדיסקואידי, אבל ברמה הקונספטואלית: Primitive streak מתחיל בצד הפוסטריורי, תאי אפיבלסט עוברים EMT ו־ingression, ומתחיל להיווצר מזודרם.
רצף האירועים המרכזי ומשפטי מפתח
רצף האירועים המרכזי
Blastocyst
↓ implantation
Egg cylinder
↓
Epiblast + ExE + Visceral endoderm
↓
Proximal-Distal axis
↓
DVE forms at distal tip
↓
DVE migrates proximally and becomes AVE
↓
Anterior side is defined
↓
Nodal/BMP/Wnt/FGF activity is positioned posteriorly
↓
Primitive streak begins in posterior epiblast
↓
Epiblast cells undergo EMT + ingression
↓
Mesoderm begins to form
משפטי מפתח
- גסטרולציה דורשת צירים מוגדרים מראש: Anterior-Posterior ו־Dorsal-Ventral נקבעים לפני תחילת התהליך.
- המעבר מבלסטוציסט ל־Egg cylinder מתרחש בזמן ההשרשה, וכולל פולריזציה של האפיבלסט ויצירת חללים.
- ה־Visceral endoderm עוטף את ה־Egg cylinder, אבל הוא כולל אוכלוסיות שונות ולא שכבה אחידה לגמרי.
- ה־DVE נוצר בקצה הדיסטלי, נודד לכיוון פרוקסימלי, והופך ל־AVE.
- ה־AVE מגדיר את הצד האנטריורי של העובר.
- RNA in situ hybridization מאפשר לראות איפה גן מתבטא לפי מיקום ה־mRNA שלו.
- Nodal מתבטא בעיקר באפיבלסט, שייך למשפחת TGFβ, ופועל דרך רצפטורי Activin/Nodal עם Cripto.
- האפיבלסט מפריש Pro-Nodal, ו־Pro-Nodal מפעיל את המסלול בעוצמה נמוכה.
- ה־ExE מגיב ל־Nodal באמצעות BMP4, Furin ו־PACE4, וכך נוצרים גרדיאנטים של BMP, פעילות Nodal ו־Wnt3.
- DVE נוצר באזור שבו יש פעילות Nodal ורמת BMP נמוכה.
- DVE/AVE מפריש אנטגוניסטים כמו Lefty1, Cer1 ו־Dkk1, שממקמים את Nodal/BMP/Wnt בצד הפוסטריורי.
- Primitive streak מתחיל בצד הפוסטריורי של האפיבלסט.
- תאי אפיבלסט שעוברים EMT ו־ingression יוצאים מהאפיבלסט ומתחילים ליצור מזודרם.
- בגסטרולציה, מיקום וזמן היציאה של תא דרך ה־Primitive streak משפיעים על גורלו.
קובץ האיורים של א.ס. זמין כאן.
דור פסקל