השאלות והתשובות שלהלן מבוססות על תוכן השיעורים, אך הן בגדר הצעה בלבד. שימו לב שבכל פעם שתרעננו את העמוד, סדר האפשרויות בכל שאלה ישתנה (התשובה הנכונה תישאר נכונה, אך המיקום שלה בין האפשרויות עשוי להשתנות).
שיעור 1: מבוא, DNA, RNA וטופולוגיה
נושאים:
- הדוגמה המרכזית
- מבנה DNA ו־dNTP
- פורינים ופירימידינים
- זיווג בסיסים וקשרי מימן
- כיווניות 5’←3’
- דו־גדיל אנטי-פרללי
- Major Groove ו־Minor Groove
- הכוחות המייצבים את ה־DNA
- Tm והיברידיזציה
- RNA לעומת DNA
- אנזימי רסטריקציה וליגאז
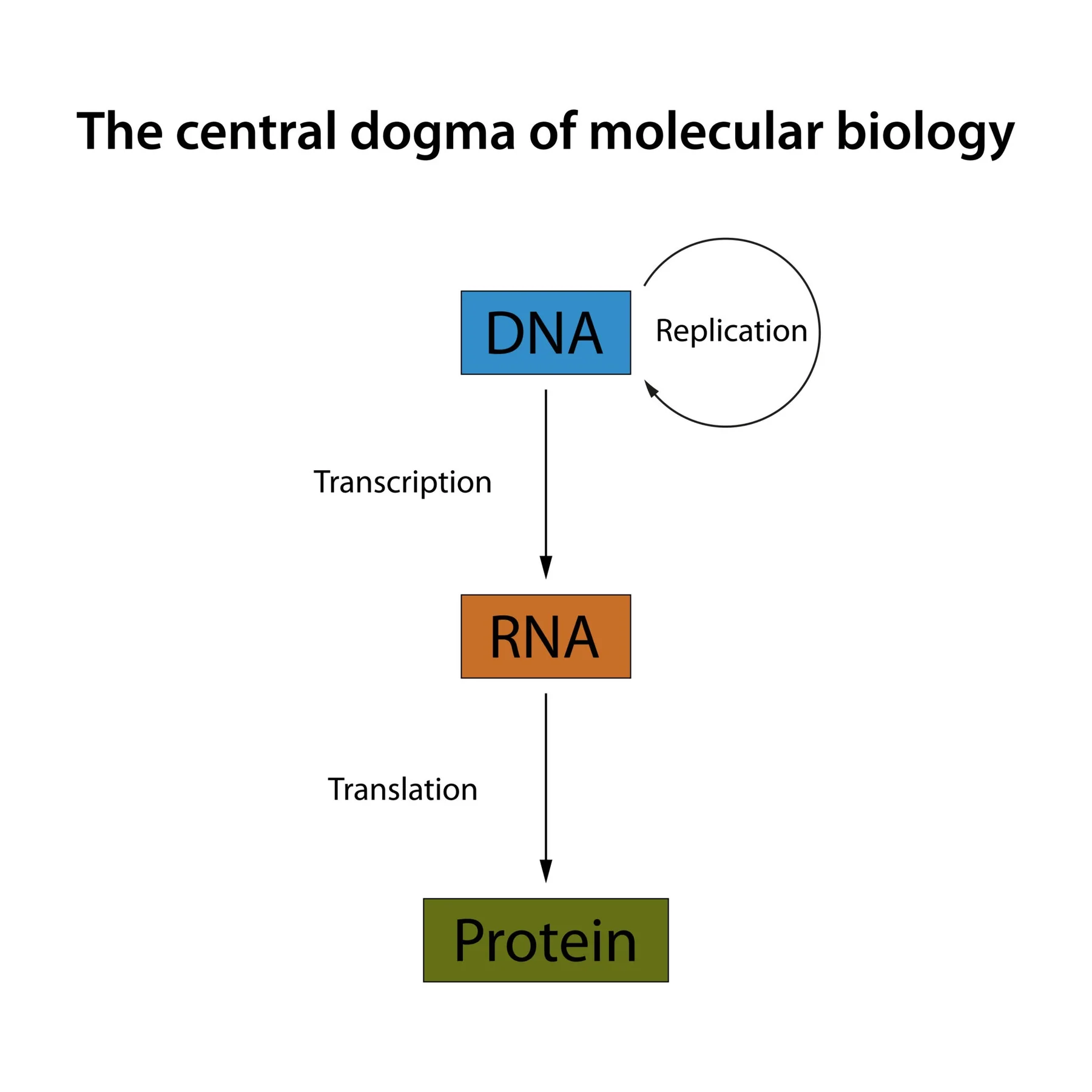
שאלה 1: הדוגמה המרכזית
איזה מהמשפטים הבאים מתאר בצורה הטובה ביותר את הדוגמה המרכזית, כפי שהוצגה בשיעור?
- זרימת המידע הביולוגי היא וקטורית וחד־סטרית, מגרעין התא אל הציטופלזמה, ומתבצעת אך ורק דרך תעתוק של DNA ל־mRNA ותרגומו לחלבון.
- זרימת המידע היא מ־DNA ל־RNA לחלבון, אך קיימים גם חריגים (כמו שעתוק מהופך) והרחבות (כמו RNA שאינו מקודד).
- הדוגמה המרכזית קובעת כי המידע הגנטי מאוחסן ב־DNA, עובר שכפול עצמי, ומתבטא תמיד כחלבון פונקציונלי באמצעות מתווך של RNA שליח.
- זרימת המידע היא תהליך מעגלי שבו חלבונים יכולים להכתיב את רצף ה־RNA, אשר בתורו משמש כתבנית ליצירת גדילי DNA חדשים בתא.
פתרון
התשובה הנכונה היא (2).
הדוגמה המרכזית מתארת מעבר מידע מ־DNA ל־RNA לחלבון. יחד עם זאת, השיעור הדגיש שהתמונה כיום מורכבת יותר:
- רטרווירוסים מבצעים רוורס טרנסקריפציה מ־RNA ל־DNA
- RNA יכול לפעול גם כמולקולה קטליטית (ריבוזים)
- RNA יכול להשפיע על DNA, למשל דרך XIST
לכן לא נכון להציג את הדוגמה המרכזית ככלל קשיח ללא חריגים.
לגבי החלופות (ייתכן שמופיעות אצלכם בסדר שונה):
- חלופה 1: נשמעת “מדעית” מאוד ומשתמשת במונחים נכונים (וקטורית, גרעין, ציטופלזמה), אך היא שגויה כי היא טוענת שהתהליך הוא אך ורק כזה, תוך התעלמות מחריגים כמו רטרו־וירוסים או RNA שלא הופך לחלבון.
- חלופה 3: זו חלופה “חצי נכונה”. היא מתארת נכון את אחסון המידע והשכפול, אך הטעות שלה היא במילה “תמיד”. לא כל RNA הופך לחלבון (יש tRNA, rRNA וכו’), ולכן היא פחות מדויקת מהתשובה הנכונה.
- חלופה 4: משתמשת במונח “תהליך מעגלי” שיכול לבלבל סטודנטים שחושבים על אינטראקציות בין חלבונים ל־DNA (כמו פקטורי תעתוק), אך היא שגויה מהיסוד לגבי כיוון זרימת המידע הגנטי עצמו.
שאלה 2: מבנה ה־dNTP
איזה צירוף מתאר נכון את מרכיבי היחידה הבסיסית של DNA?
- סוכר, שתי קבוצות פוספט, ובסיס חנקני
- דאוקסיריבוז, שלוש קבוצות פוספט, ובסיס חנקני
- ריבוז, שלוש קבוצות פוספט, ובסיס חנקני
- דאוקסיריבוז, חומצת אמינו, וקבוצת פוספט
פתרון
התשובה הנכונה היא (2).
אבן הבניין הבסיסית שהוצגה בשיעור היא dNTP - deoxyribonucleotide triphosphate. היא כוללת:
- דאוקסיריבוז
- שלוש קבוצות פוספט
- בסיס חנקני
לא מדובר בריבוז, ולא בחומצת אמינו.
מקור: שיעור 1 - מבנה ה־DNA
שאלה 3: פורינים ופירימידינים
איזה מהזוגות הבאים כולל שני פורינים?
- Cytosine ו־Thymine
- Adenine ו־Guanine
- Adenine ו־Thymine
- Cytosine ו־Uracil
פתרון
התשובה הנכונה היא (2).
לפי החלוקה שהוצגה בשיעור:
- פורינים: Adenine, Guanine
- פירימידינים: Cytosine, Thymine
זה בסיסי. לבלבל כאן במבחן זה הפסד נקודות מיותר.
מקור: שיעור 1 - מבנה ה־DNA
שאלה 4: זיווג בסיסים
איזה משפט נכון לגבי זיווג הבסיסים ב־DNA?
- A מזדווג עם G באמצעות שני קשרי מימן
- C מזדווג עם T באמצעות שלושה קשרי מימן
- A מזדווג עם T באמצעות שני קשרי מימן
- כל זוג בסיסים יוצר אותו מספר קשרי מימן
פתרון
התשובה הנכונה היא (3).
ב־DNA הזיווג הוא ספציפי:
- A-T עם שני קשרי מימן
- G-C עם שלושה קשרי מימן
זו אחת הסיבות לכך שאזורים עשירים ב־GC יציבים יותר.
שאלה 5: כיווניות הסינתזה
מהו התיאור הנכון של בניית הפולימר הגרעיני?
- השרשרת נבנית בכיוון 3’←5’, כאשר ה־OH על פחמן 5’ תוקף את הנוקלאוטיד הנכנס
- השרשרת נבנית בכיוון 5’←3’, כאשר ה־OH על פחמן 3’ של הגדיל תוקף את הפוספט של הנוקלאוטיד הנכנס
- השרשרת נבנית ללא כיווניות קבועה, בהתאם לסוג הבסיס
- הסינתזה מתבצעת על ידי קשרי מימן בלבד
פתרון
התשובה הנכונה היא (2).
הכיווניות של חומצות גרעין היא 5’←3’. ההוספה מתבצעת על ידי התקפה נוקלאופילית של קבוצת ה־OH על פחמן 3’ של הנוקלאוטיד האחרון, על קבוצת הפוספט של הנוקלאוטיד הנכנס, ונוצר קשר פוספודיאסטרי.
מי שלא שולט בזה עד הסוף, ייפול אחר כך גם בשכפול וגם בשעתוק.
מקור: שיעור 1 - בניית הפולימר
שאלה 6: הדו־גדיל
איזה מהמשפטים הבאים נכון לגבי ה־DNA הדו־גדילי?
- שני הגדילים רצים באותו כיוון ולכן נקראים מקבילים
- שני הגדילים מסודרים אנטי-פרללית, והבסיסים נמצאים כלפי חוץ
- שני הגדילים מסודרים אנטי-פרללית, והשלד הפוספוסוכרי נמצא מבחוץ
- השלד והבסיסים מסודרים באותו צד של המולקולה
פתרון
התשובה הנכונה היא (3).
ב־DNA דו־גדילי:
- שני הגדילים הם אנטי-פרלליים
- השלד הפוספוסוכרי נמצא מבחוץ
- הבסיסים נמצאים מבפנים
זה בדיוק הסידור שמאפשר מצד אחד יציבות, ומצד שני גישה של חלבונים למידע דרך הגרובים.
מקור: שיעור 1 - הדו־גדיל
שאלה 7: Major Groove ו־Minor Groove
מדוע חלבונים רבים מעדיפים להיקשר דרך ה־Major Groove?
- כי זהו האזור שבו נמצאות קבוצות הפוספט
- כי הוא צר יותר ולכן מדויק יותר
- כי הוא רחב ועמוק יותר
- כי רק שם קיימים קשרי מימן
פתרון
התשובה הנכונה היא (3).
החלבונים “קוראים” את הרצף דרך הגרובים. ה־Major Groove רחב ועמוק יותר, ולכן נותן גישה טובה יותר למידע שבפנים. יש גם חלבונים שנקשרים ל־Minor Groove, אבל זו לא ברירת המחדל העיקרית.
שאלה 8: כוחות מייצבים ו־Tm
איזה שינוי צפוי להעלות את טמפרטורת ההיתוך (Tm) של מקטע DNA?
- הגדלת שיעור זוגות A-T
- קיצור המולקולה לאזור קצר יותר
- הגדלת שיעור זוגות G-C
- החלפת DNA ב־RNA חד־גדילי
פתרון
התשובה הנכונה היא (3).
DNA עשיר ב־G/C יעבור היתוך בטמפרטורה גבוהה יותר מאשר DNA עשיר ב־A/T, משום שבין G ל־C יש שלושה קשרי מימן לעומת שניים בין A ל־T.
נכון, בפועל גם stacking energy חשוב ליציבות. לכן מי שחושב רק בסיסים ולא מבין מבנה כולל, מפספס חצי מהסיפור.
מקור: שיעור 1 - הכוחות שמחזיקים את ה־DNA ביחד; Melting Temperature
שאלה 9: היברידיזציה
מה נכון לגבי היברידיזציה של DNA?
- זהו תהליך שבו DNA דו־גדילי הופך תמיד ל־RNA
- אחרי קירור חיבור מחדש מתרחש באופן ספונטני
- היברידיזציה דורשת פירוק קשרים קוולנטיים בשלד
- היברידיזציה מתרחשת רק בתוך התא ולא במעבדה
פתרון
התשובה הנכונה היא (2).
אחרי דנטורציה והפרדת הגדילים, חד־גדילים משלימים יכולים לעבור היברידיזציה - למצוא זה את זה, ליצור גרעין זיווג, ולהיבנות מחדש לדו־גדיל. השיעור גם קישר זאת ליישומים כמו FISH, Microarray וגישות ריצוף.
מקור: שיעור 1 - היברידיזציה
שאלה 10: RNA לעומת DNA
איזה מהמשפטים הבאים נכון?
- RNA יציב יותר מ־DNA כי יש לו OH על פחמן 2’
- DNA משתמש ב־Uracil כדי שמערכות תיקון יזהו דמינציה של Cytosine
- RNA כולל בדרך כלל דאוקסיריבוז ו־T במקום U
- DNA משתמש ב־Thymine ולא ב־Uracil, בין השאר כדי לאפשר זיהוי של דמינציה של Cytosine
פתרון
התשובה הנכונה היא (4).
הבדלים מרכזיים:
- ב־RNA יש ריבוז עם OH על פחמן 2’, ולכן הוא פחות יציב
- ב־DNA יש Thymine ולא Uracil
- אם Cytosine עובר דמינציה והופך ל־Uracil, מערכת התיקון יכולה לזהות שיש כאן תקלה דווקא משום שב־DNA לא אמור להיות U
הנקודה הזאת חשובה כי היא מחברת בין מבנה כימי לבין היגיון ביולוגי.
מקור: שיעור 1 - RNA לעומת DNA
שאלה 11: אנזימי רסטריקציה וליגאז
מהו התיאור הנכון ביותר של תהליך קלוניה גנטית בסיסי, כפי שהוצג בשיעור?
- אנזים רסטריקציה מזהה את אתרי המטרה ומחבר את הקצוות המשלימים באמצעות קשרי מימן, בעוד שליגאז יוצר את הקצוות הדביקים (Sticky ends).
- חותכים מקטע DNA ופלסמיד עם אותו אנזים רסטריקציה, הקצוות משלימים עוברים היברידיזציה, ואז ליגאז סוגר את הקשר.
- ליגאז מחבר את מקטע ה־DNA לפלסמיד על ידי יצירת קשרי מימן בין הבסיסים, ואנזים רסטריקציה מייצב את השלד הסוכר-פוספטי של המולקולה החדשה.
- שימוש באנזימי רסטריקציה שונים למקטע ה־DNA ולפלסמיד מאפשר לליגאז לבצע איחוי ישיר ומהיר יותר, ללא צורך בשלב יצירת קשרי המימן בין הבסיסים.
פתרון
התשובה הנכונה היא (2).
בקלוניה גנטית בסיסית:
- חותכים את מקטע ה־DNA הרצוי
- חותכים גם פלסמיד עם אותו אנזים רסטריקציה
- הקצוות המתאימים יכולים לעבור היברידיזציה
- Ligase סוגר את הקשר ויוצר מולקולה רציפה
זה אחד המקומות שבהם כדאי להבין מנגנון, לא רק לשנן שם של אנזים.
לגבי החלופות השגויות (ייתכן שמופיעות בסדר שונה):
-
חלופה 1 מבלבלת בין מי שחותך (יוצר קצוות) לבין מי שמחבר, אבל משתמשת במונחים מקצועיים שנשמעים נכונים.
-
חלופה 3 נוגעת בסוגי הקשרים הכימיים. היא טוענת שליגאז יוצר קשרי מימן (מה שקורה באופן ספונטני בהיברידיזציה), בעוד שתפקידו האמיתי הוא ליצור קשר קוולנטי (פוספודיאסטרי).
-
חלופה 4 מציעה “קיצור דרך” שנשמע יעיל מבחינה הנדסית, אך שגוי ביולוגית (כי בלי התאמה של הקצוות, ליגאז לא יוכל לחבר את המולקולות ביעילות).
שיעור 2: שיבוט DNA, טופולוגיית DNA וסופרקוילינג
נוקלאזות, אנזימי רסטריקציה, ליגאז, טופולוגיית DNA, סופרקוילינג, Lk/Tw/Wr, ג׳ל אלקטרופורזה, EtBr, SYBR Green ו־PFGE.
שאלה 1: אקסונוקלאז מול DNA מעגלי
איזו מהפעולות הבאות צפויה לא להשפיע כלל על פלסמיד DNA דו־גדילי, מעגלי וסגור קוולנטית?
- טיפול באקסונוקלאז
- טיפול באנדונוקלאז
- יצירת nick באחד הגדילים
- פעולה של טופואיזומראז
פתרון
התשובה הנכונה היא (1).
אקסונוקלאז פועל מהקצה, ולכן על מולקולה מעגלית שאין לה קצוות - הוא לא יכול להתחיל לחתוך.
למה האחרות שגויות:
- (2) אנדונוקלאז חותך בתוך המולקולה, ולכן בהחלט יכול להשפיע.
- (3) גם nick אחד משנה את המצב הטופולוגי ומשחרר את המתח הפיתולי.
- (4) טופואיזומראז משנה את ה־Lk ולכן בהחלט משפיע.
שאלה 2: אנזימי רסטריקציה
איזה מהמשפטים הבאים מתאר נכון אנזימי רסטריקציה?
- הם אקסונוקלאזות שחותכות מהקצה ברצפים עשירים ב־GC
- הם אנדונוקלאזות שמזהות רצף ספציפי, לרוב פלינדרומי
- הם חותכים רק RNA חד־גדילי ויוצרים תמיד blunt ends
- הם אוטמים ניקים לאחר היברידיזציה של קצוות דביקים
פתרון
התשובה הנכונה היא (2).
אנזימי רסטריקציה הם אנדונוקלאזות sequence-specific, ולרוב הרצף שהם מזהים הוא פלינדרומי.
למה האחרות שגויות:
- (1) הם לא אקסונוקלאזות ולא חותכים “מהקצה”.
- (3) הם פועלים על DNA ולא “רק RNA”, וגם לא תמיד יוצרים blunt ends.
- (4) זה תפקידו של ליגאז, לא של אנזים רסטריקציה.
שאלה 3: Sticky ends לעומת blunt ends
מה היתרון המרכזי של sticky ends בקלוניה גנטית?
- הם מכילים ATP ולכן לא צריך ליגאז
- הם יכולים לעבור היברידיזציה ספונטנית עם קצה משלים מתאים
- הם יציבים יותר כימית מקשר פוספודיאסטרי
- הם מאפשרים חיתוך גם במולקולה שאין לה קצוות
פתרון
התשובה הנכונה היא (2).
בקצוות דביקים נחשף רצף חד־גדילי קצר שיכול לעבור היברידיזציה ספונטנית עם קצה משלים.
למה האחרות שגויות:
- (1) גם אחרי ההיברידיזציה נשארים ניקים, ולכן עדיין צריך ליגאז.
- (3) sticky end הוא לא קשר כימי יציב יותר, אלא אזור זמני של זיווג בסיסים.
- (4) זו לא פונקציה של sticky ends בכלל.
שאלה 4: תפקיד הליגאז
לאחר שחיברו שני מקטעי DNA שנחתכו באותו אנזים רסטריקציה, מה בדיוק נשאר לליגאז לעשות?
- לפרק את קשרי המימן בין הגדילים
- להוסיף dNTPs חדשים לקצות הגדילים
- לסגור ניקים בשלד הפוספוסוכרי תוך שימוש באנרגיית ATP
- לזהות את הרצף הפלינדרומי ולחתוך מחדש את ה־DNA
פתרון
התשובה הנכונה היא (3).
אחרי שהקצוות הדביקים עברו היברידיזציה, השלד עדיין לא רציף - יש nick. הליגאז משתמש ב־ATP כדי לאפשר יצירת קשר פוספודיאסטרי.
למה האחרות שגויות:
- (1) ליגאז לא מפרק קשרי מימן.
- (2) ליגאז לא מאריך שרשרת כמו פולימראז.
- (4) זה תיאור של אנזים רסטריקציה, לא ליגאז.
שאלה 5: תנאי ליצירת מתח פיתולי
מהו התנאי הבסיסי לכך של־DNA יהיה מצב טופולוגי עם מתח פיתולי משמעותי?
- שהמולקולה תהיה תמיד עשירה ב־GC
- שהקצוות יהיו מקובעים, או שלא יהיו קצוות כלל
- שה־DNA יהיה בהכרח ליניארי
- שיתקיים לפחות nick אחד באחד הגדילים
פתרון
התשובה הנכונה היא (2).
כדי להחזיק מתח פיתולי, הקצוות חייבים להיות מוחזקים או שהמולקולה תהיה מעגלית. אחרת המתח פשוט ישתחרר.
למה האחרות שגויות:
- (1) הרכב בסיסים משפיע על יציבות, לא על עצם היכולת להחזיק טופולוגיה.
- (3) להפך - DNA מעגלי הוא דוגמה קלאסית למצב כזה.
- (4) nick דווקא משחרר מתח פיתולי, לא יוצר אותו.
שאלה 6: Negative supercoiling
איזה מהמשפטים הבאים מתאר נכון את התכונות או התפקוד של negative supercoiling ב־DNA?
- הוא גורם לעלייה בטמפרטורת ההתכה ($T_m$) של ה־DNA, כיוון שהפיתול השלילי מהדק את קשרי המימן בין הבסיסים המזווגים.
- הוא המצב הנפוץ ברוב התאים בטבע, שכן המתח הפיתולי שהוא משרה מקטין את כמות האנרגיה החופשית הנדרשת להפרדת הגדילים.
- הוא מוגדר כמצב שבו ה־Linking Number ($Lk$) גדול מה־Linking Number של המולקולה במצב הרפוי ($Lk_0$), מה שיוצר “לחץ” לפרימת הדו־גדיל.
- במצב זה, ערך ה־Writhe ($Wr$) הוא בהכרח חיובי, מה שמאפשר אריזה יעילה יותר של ה־DNA בתוך הגרעין או הנוקלאואיד.
פתרון
התשובה הנכונה היא (2).
Negative supercoiling מרפה את הדו־גדיל יחסית, ולכן מקל על פתיחה מקומית של ה־DNA - דבר חשוב לשעתוק ולרפליקציה.
למה האחרות שגויות: (לא בהכרח בסדר הופעתן)
-
חלופה 1 (יציבות תרמית): היא נשמעת הגיונית למי שחושב ש”פיתול” תמיד “מהדק”. למעשה, supercoiling שלילי דווקא מקל על פתיחת הגדילים (מוריד את ה־$T_m$), בעוד ש־positive supercoiling (הנפוץ למשל בחיידקים תרמופיליים) הוא זה שמהדק את המבנה ומגן עליו מחום.
-
חלופה 3 (הנוסחה הטופולוגית): כאן הטעות היא מתמטית-הגדרתית. ב־negative supercoiling, ה־$Lk$ הוא קטן יותר מה־$Lk_0$ (חסרים פיתולים ביחס למצב הרפוי). סטודנטים לעיתים מתבלבלים בכיוון של ה־Delta $Lk$.
-
חלופה 4 (Wr vs Tw): חלופה זו נוגעת בפרמטר ה־Writhe. ב־DNA “ימני” (B-DNA), פיתול־על שלילי מתבטא בדרך כלל ב־$Wr$ שלילי. הטענה שערך חיובי עוזר לאריזה נשמעת משכנעת כי supercoiling אכן עוזר לאריזה, אך הסימן (חיובי/שלילי) שגוי.
הערות נוספות:
- הוא מהדק את הדו־גדיל ולכן מקשה על פתיחה זה יותר מתאים ל־positive supercoiling.
- הוא נוצר רק לאחר חימום DNA מעגלי חימום DNA מעגלי מתואר בשיעור בצורה אחרת, ולא זו ההגדרה ל־negative supercoiling.
- הוא מוגדר כמצב שבו Wr תמיד חיובי ב־negative supercoiling ה־Wr שלילי, לא חיובי.
שאלה 7: שמירת Lk
במולקולת DNA מעגלית סגורה, אם ה־Tw ירד ב־12 בלי שנחתך השלד הפוספוסוכרי, מה חייב לקרות?
- ה־Lk ירד גם הוא ב־12
- ה־Wr יעלה ב־12
- ה־Wr ירד ב־12
- ה־Lk יהפוך לאפס
פתרון
התשובה הנכונה היא (2).
במולקולה סגורה מתקיים:
\[Lk = Tw + Wr\]אם ה־Lk קבוע וה־Tw ירד ב־12, ה־Wr חייב לעלות ב־12 כדי לשמור על אותו Lk.
למה האחרות שגויות:
- (1) Lk לא משתנה בלי חיתוך וחיבור מחדש של השלד.
- (3) אם גם Wr ירד, הסכום הכולל היה משתנה.
- (4) אין שום סיבה ש־Lk יהפוך לאפס.
שאלה 8: Nick בפלסמיד סופרקויל
פלסמיד מעגלי סגור במצב negative supercoiling עבר nick באחד הגדילים. מה צפוי לקרות?
- המולקולה תתפרק לשני מקטעים ליניאריים
- ה־Wr יתאפס, המולקולה תישאר מעגלית, והניידות שלה בג׳ל תרד
- ה־Tw יתאפס מיד והמולקולה תיהפך לקטננט
- האקסונוקלאז יתחיל אוטומטית לפרק את כל המולקולה
פתרון
התשובה הנכונה היא (2).
Nick אחד מספיק כדי לשחרר את המתח הפיתולי (Wr→0), אבל המולקולה עדיין נשארת מעגלית כי הגדיל השני שלם. בג׳ל היא תתנהג כ־relaxed circular ותנוע לאט יותר.
למה האחרות שגויות:
- (1) זה היה קורה בחיתוך דו־גדילי, לא ב־nick.
- (3) Tw לא “מתאפס מיד” כאן, והמבנה לא הופך לקטננט.
- (4) לא קיים מנגנון כזה “אוטומטית”.
שאלה 9: סדר הניידות בג׳ל
לאותה מולקולת DNA, עם אותו מספר זוגות בסיסים, מהו סדר הניידות הצפוי בג׳ל אגרוז - מהמהיר ביותר אל האיטי ביותר?
- relaxed circular → linear → supercoiled
- linear → supercoiled → relaxed circular
- supercoiled → linear → relaxed circular
- supercoiled → relaxed circular → linear
פתרון
התשובה הנכונה היא (3).
למולקולות באותו גודל, ההבדל הוא בצורה הטופולוגית:
- supercoiled - הכי קומפקטית, נעה הכי מהר
- linear - ביניים
- relaxed circular - הכי מסורבלת במרחב, נעה הכי לאט
שאלה 10: EtBr לעומת SYBR Green
איזה מהמשפטים הבאים נכון?
- EtBr נקשר ל־Major Groove ואילו SYBR Green עובר אינטרקלציה
- EtBr עובר אינטרקלציה ועלול להשפיע על Tw, ואילו SYBR Green נקשר ל־Major Groove
- שני הצבעים פועלים רק על RNA ולכן לא מתאימים לג׳ל DNA
- שני הצבעים משמשים רק להפרדה לפי גודל, ולא לויזואליזציה
פתרון
התשובה הנכונה היא (2).
- EtBr עובר אינטרקלציה בין בסיסי ה־DNA
- SYBR Green נקשר ל־Major Groove
- לכן EtBr גם יכול להשפיע על המצב המבני/טופולוגי של ה־DNA
למה האחרות שגויות:
- (1) זו החלפה בין המנגנונים.
- (3) שניהם משמשים ל־DNA.
- (4) שניהם משמשים קודם כל לויזואליזציה של DNA.
שאלה 11: PFGE
מדוע משתמשים ב־PFGE במקום בג׳ל אגרוז רגיל?
- כדי למנוע דנאטורציה תרמית של גדילי ה־DNA כתוצאה מהחום הנוצר בהרצה של מתח גבוה, באמצעות פעימות חשמליות קצרות המאפשרות קירור של המערכת.
- כדי להתגבר על תופעת ה־limiting mobility, שבה מקטעי DNA מעל גודל מסוים נעים כולם באותה מהירות בשדה חשמלי קבוע ולא ניתנים להפרדה.
- השינוי בכיוון השדה החשמלי ב־PFGE נועד להגדיל את המטען החשמלי האפקטיבי של המולקולה, ובכך לאפשר לה לעבור דרך פורות קטנות במיוחד של אגרוז בריכוז גבוה.
- PFGE משמש להפרדה של DNA ליניארי מ־DNA מעגלי (כמו פלסמידים) באותו משקל מולקולרי, מכיוון שרק DNA מעגלי מסוגל לשנות את כיוון האוריינטציה שלו במהירות הנדרשת.
פתרון
התשובה הנכונה היא (2).
Pulsed field gel electrophoresis (PFGE) (chromosome size)
מולקולות DNA גדולות מאוד, כמו כרומוזומים שלמים, לא נעות טוב בג׳ל רגיל. ב־PFGE משנים כל הזמן את כיוון השדה החשמלי, וכך המולקולה “מסתדרת מחדש” ומסוגלת להתקדם דרך הג׳ל.
בשיעור בעיקר התייחסנו להקשר של מולקולות DNA ארוכות מאוד, ופחות לפיזיקה של התהליך.
-
חלופה 1 (ניהול חום): זה נשמע הגיוני כי הרצות PFGE הן ארוכות מאוד (לפעמים ימים) ומערכות PFGE אכן כוללות בקרת טמפרטורה. עם זאת, הפעימות החשמליות הן לא הדרך שבה המערכת מתקררת, אלא הדרך שבה המולקולות “מתמרנות”.
-
חלופה 2 (התשובה הנכונה): משתמשת במונח המקצועי Limiting Mobility. ב־DNA ארוך מאוד (מעל 50kb בערך), המולקולות נמתחות לאורך קווי השדה ונעות כמו “נחש” (Reptation). בשדה קבוע, כולן נעות באותה מהירות ללא קשר לאורכן. PFGE מכריח אותן להתכווץ ולהסתדר מחדש בכל פעימה, וזמן ה”הסתדרות” הזה תלוי באורך המולקולה.
-
חלופה 3 (ריכוז האגרוז): מערבבת מושגים של מטען חשמלי וצפיפות הג’ל. ב־PFGE דווקא משתמשים לעיתים קרובות בריכוזי אגרוז סטנדרטיים, והמטען של ה־DNA לא משתנה.
-
חלופה 4 (ליניארי מול מעגלי): זו טעות קלאסית. למעשה, PFGE יעיל בעיקר למולקולות ליניאריות גדולות מאוד (כמו כרומוזומים של שמרים). מולקולות מעגליות גדולות נוטות “להינעל” בתוך הרשת של הג’ל בזמן שינוי כיוון השדה ודווקא מתקשות לזוז ב־PFGE.
שיעור 3: ארגון הגנום, כרומטין ואפיגנטיקה
ארגון הגנום, כרומוזומים, כרומטין, נוקלאוזומים, וריאנטים של היסטונים, רגולציה אפיגנטית וסופרקוילינג כתוצר לוואי של טרנסקריפציה ורפליקציה.
שאלה 1: כרומוזומים הומולוגיים מול כרומטידות אחיות
איזה מהמשפטים הבאים מתאר נכון את הקשר בין כרומוזומים הומולוגיים לכרומטידות אחיות?
- כרומוזומים הומולוגיים הם העתקים זהים לחלוטין ברמת רצף הנוקלאוטידים, בעוד שכרומטידות אחיות נושאות את אותם גנים אך עשויות להכיל אללים שונים.
- כרומוזומים הומולוגיים מכילים את אותה סדרת גנים אך עשויים לשאת אללים שונים, בעוד שכרומטידות אחיות הן תוצר של שכפול ה־DNA וזהות גנטית זו לזו.
- כרומטידות אחיות נוצרות במהלך שלב ה־G2 של מחזור התא ומופרדות זו מזו במהלך המיוזה הראשונה, בעוד שכרומוזומים הומולוגיים נצמדים זה לזה רק במיטוזה.
- המונח “הומולוגי” מתאר את זוג הכרומטידות המחוברות פיזית באזור הצנטרומר לאחר סיום שלב ה־S, שמקורן באותו הורה.
פתרון
התשובה הנכונה היא (2).
- כרומוזומים הומולוגיים הם זוג כרומוזומים - אחד מהאב ואחד מהאם - שנושאים אותם גנים.
- כרומטידות אחיות הן שתי מולקולות DNA זהות שנוצרו משכפול של אותו כרומוזום בשלב S.
למה האחרות שגויות: (שוב, ייתכן שהן מופיעות בסדר שונה)
-
חלופה 1 (היפוך הגדרות): היא משתמשת במונחים נכונים (“אללים”, “רצף נוקלאוטידים”) אבל מחליפה בין המושגים. סטודנטים לעיתים מתבלבלים וחושבים שהומולוגיים הם ה”זהים” בגלל השם הדומה, בעוד שהם למעשה רק “דומים” (אחד מאמא, אחד מאבא).
-
חלופה 2 (התשובה הנכונה): בגנטיקה היה אפשר להוסיף כאן את המושג לוקוס (מיקום פיזי על הכרומוזום) ואת ההבחנה בין גנים (למשל: גן לצבע עיניים) לאללים (למשל: כחול או חום), מה שהופך את ההגדרה למדויקת ומדעית יותר.
-
חלופה 3 (מחזור התא): חלופה זו “מפגיזה” במושגים משלבי החלוקה (G2, אנאפאזה, מיוזה). היא שגויה כי כרומטידות אחיות נוצרות בשלב ה־S (שכפול), והן מופרדות במיוזה II (או במיטוזה), לא במיוזה I.
-
חלופה 4 (מבנה פיזי): מנסה לבלבל בין הקשר הפיזי (הצנטרומר שמחבר אחיות) לבין המושג “הומולוגי”. היא נשמעת משכנעת כי היא מתארת מבנה שקיים בתא, אך מצמידה לו את השם הלא נכון.
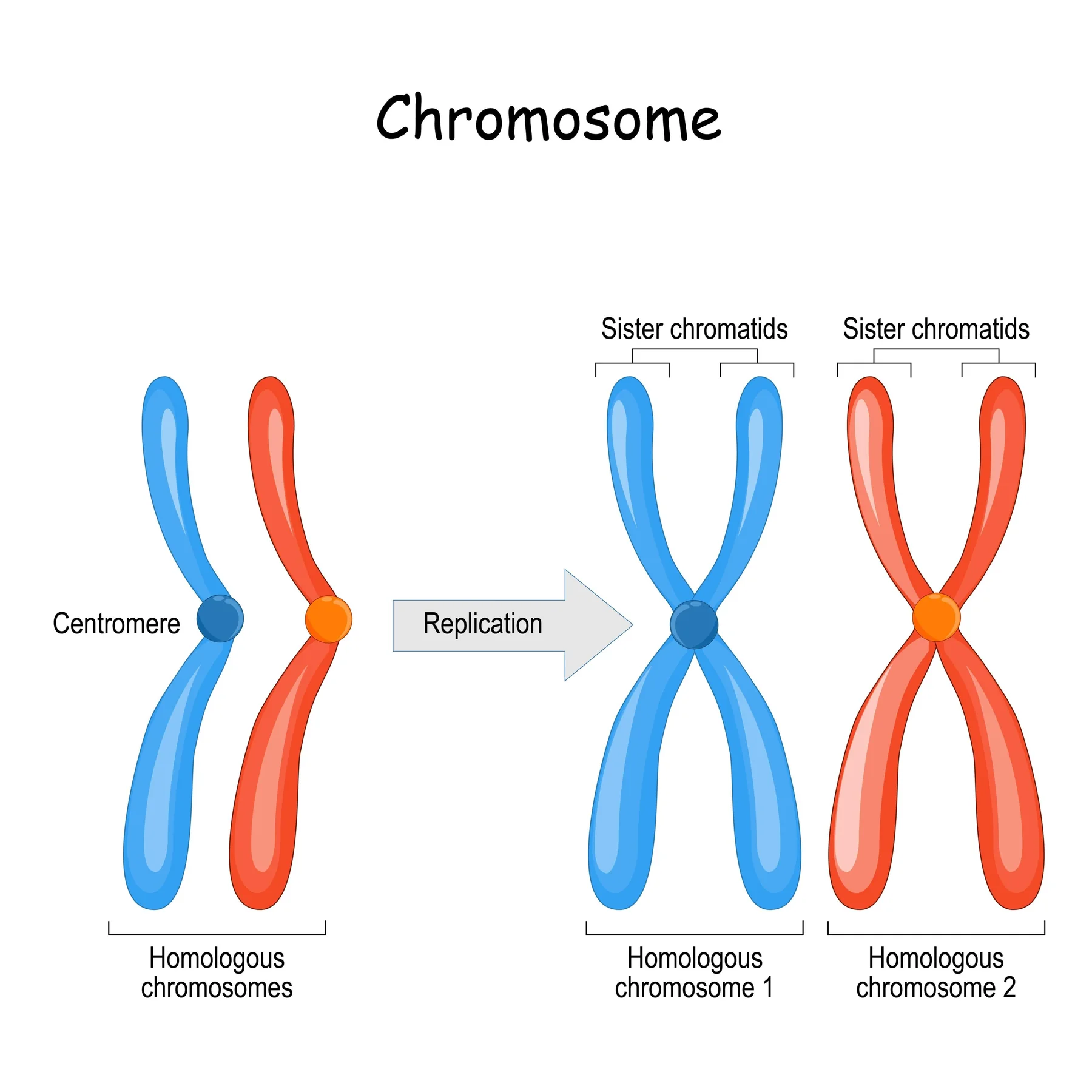
שאלה 2: מבנה הכרומוזום לאחר שלב S
איזה תיאור נכון לכרומוזום אנושי לאחר שלב S ולפני אנאפאזה?
- כל כרומוזום בנוי משתי כרומטידות אחיות המחוברות זו לזו
- כל כרומוזום בנוי משני כרומוזומים הומולוגיים מאוחים
- כל כרומוזום מכיל רק גדיל DNA יחיד
- מספר הכרומוזומים בתא מוכפל כי כל כרומטידה נחשבת כרומוזום נפרד בשלב זה
פתרון
התשובה הנכונה היא (1).
לאחר שלב S כל כרומוזום כבר שוכפל, ולכן הוא בנוי משתי כרומטידות אחיות שמחוברות עד להפרדה במיטוזה.
למה האחרות שגויות:
- (2) הומולוגיים אינם “מאוחים” זה לזה.
- (3) כרומוזום לאחר שכפול אינו גדיל יחיד.
- (4) בשלב הזה מדברים עדיין על אותו כרומוזום משוכפל, לא על הכפלת מספר הכרומוזומים בגלל כל כרומטידה.
שאלה 3: צנטרומר
מהו התפקיד המרכזי של הצנטרומר?
- לשמש אתר ראשית הכפלה עיקרי של הכרומוזום
- להגן על קצות הכרומוזום מפני אקסונוקלאזות
- לאפשר קישור של הכרומוזום לסיבי הכישור דרך הקינטוכור
- להחזיק את הנוקלאוזום הראשון בכל זרוע כרומוזומלית
פתרון
התשובה הנכונה היא (3).
הצנטרומר הוא האזור שעליו נבנה הקינטוכור, והקינטוכור הוא מבנה הקישור לסיבי הכישור בזמן חלוקת התא.
למה האחרות שגויות:
- (1) אתר ראשית הכפלה הוא אלמנט אחר.
- (2) זה תפקידם של הטלומרים.
- (4) זה אינו תפקיד הצנטרומר.
שאלה 4: טלומרים
איזה מהמשפטים הבאים אינו תפקיד של טלומרים לפי השיעור?
- הגנה על קצות הכרומוזום מפני אקסונוקלאזות
- מניעת איחוי בין כרומוזומים על ידי מערכת תיקון DNA
- עיגון הקצוות כך שניתן יהיה לשמר מתח פיתולי בכרומוזום ליניארי
- קביעה ישירה של מיקום הקינטוכור על הכרומוזום
פתרון
התשובה הנכונה היא (4).
מיקום הקינטוכור נקבע על ידי הצנטרומר, לא על ידי הטלומרים.
למה האחרות נכונות:
- (1) טלומרים מגנים על הקצה מפני פירוק.
- (2) הם מונעים מהתא לזהות קצה כרומוזומלי כשבר DNA.
- (3) הם גם מסייעים בעיגון קצות הכרומוזום הליניארי.
שאלה 5: אלמנטים חיוניים לכרומוזום
איזה שילוב נדרש כדי שמקטע DNA יוכל להתקיים ככרומוזום יציב ועצמאי בתא איקריוטי?
- צנטרומר, טלומרים ואתר ראשית הכפלה
- צנטרומר, פרומוטר ואינהנסר
- אתר ראשית הכפלה, אינטרון וטלומרים
- טלומרים, היסטון H1 ו־CENP-A
פתרון
התשובה הנכונה היא (1).
לפי השיעור, שלושת האלמנטים החיוניים הם:
- צנטרומר
- טלומרים
- Origin of replication
בלי שלושתם מקטע DNA לא יתפקד ככרומוזום עצמאי יציב.
שאלה 6: יצירת קריוטיפ
איזה מהמשפטים הבאים מתאר נכון את הסיבה לשימוש בתאים בשלב המיטוטי (ובעיקר במטאפאזה) לצורך הכנת קריוטיפ?
- בשלב המיטוטי מעטפת הגרעין מתפרקת, מה שמאפשר לצבעי ה־Giemsa לחדור בצורה הומוגנית לכל אורך ה־DNA ללא הפרעה של ממברנות תאיות.
- במטאפאזה הכרומטין מגיע לדרגת הדחיסה המקסימלית שלו (Compact Chromatin), מה שמאפשר לזהות את המורפולוגיה של הכרומוזומים ואת תבנית הפסים (Banding) הייחודית להם.
- תאי אינטרפאזה אינם מתאימים לקריוטיפ כיוון שהכרומוזומים בהם עדיין אינם קשורים לצנטרומר, ולכן לא ניתן להבחין בין זרוע p לזרוע q.
- השימוש במיטוזה הכרחי משום שרק בשלב זה ה־DNA עובר דה־מתילציה נרחבת, תהליך שמאפשר לצבעים פלואורסצנטיים להיקשר לאזורי ה־Heterochromatin.
פתרון
התשובה הנכונה היא (2).
בקריוטיפ צריך לראות כרומוזומים כיחידות נפרדות. זה קורה היטב רק כאשר ה־DNA דחוס מאוד במיטוזה.
למה האחרות שגויות:
-
חלופה 1 (מעטפת הגרעין): זו עובדה ביולוגית נכונה (המעטפת אכן מתפרקת), וזה נשמע הגיוני מבחינה טכנית (“הצבע חודר טוב יותר”), אבל זו לא הסיבה. הסיבה היא המבנה הפיזי של ה־DNA ולא הממברנה.
-
חלופה 2 (התשובה הנכונה): המושג “דחיסה מקסימלית” הוא המפתח כאן. באינטרפאזה ה־DNA נראה כמו “קערת ספגטי” (Chromatin), ורק הדחיסה המסיבית של המיטוזה מאפשרת לנו לראות “מקלות” נפרדים.
-
חלופה 3 (צנטרומר וזרועות): חלופה זו מבלבלת סטודנטים שחושבים שצנטרומר או זרועות נוצרים רק במיטוזה. למעשה, המבנה קיים, אבל הוא פשוט לא דחוס מספיק כדי שנראה אותו במיקרוסקופ אור.
-
חלופה 4 (אפיגנטיקה): משתמשת במונחים מתקדמים כמו “דה־מתילציה” ו”הטרוכרומטין” כדי להישמע מתוחכמת. סטודנטים עשויים לחשוב שיש קשר ביוכימי מורכב לצביעה, בעוד שהסיבה היא בעיקר מכנית-מורפולוגית.
שאלה 7: שימושי קריוטיפ
איזה סוג שינוי הוא המתאים ביותר לזיהוי באמצעות קריוטיפ קלאסי?
- החלפת בסיס בודד בגן יחיד
- חסר של שני נוקלאוטידים בתוך אקסון
- טריזומיה של כרומוזום 21
- מוטציית missense ב־BRCA1
פתרון
התשובה הנכונה היא (3).
קריוטיפ מזהה שינויים גדולים וגסים ברמת כרומוזומים - למשל טריזומיות, טרנסלוקציות גדולות וחסרים/כפולים גדולים.
למה האחרות שגויות:
- (1), (2), (4) הן שינויים קטנים מדי לרזולוציה של קריוטיפ.
שאלה 8: כרומוזום חיידקי מול כרומוזום איקריוטי
איזה מהמשפטים הבאים מתאר נכון כרומוזום חיידקי טיפוסי?
- ליניארי, עם טלומרים והרבה אתרי ראשית הכפלה
- מעגלי, ללא טלומרים וללא צנטרומר
- ליניארי, עם נוקלאוזומים והיסטון H1
- מעגלי, אך זקוק לקינטוכור לצורך חלוקה לתאי בת
פתרון
התשובה הנכונה היא (2).
כרומוזום חיידקי טיפוסי הוא:
- מעגלי
- ללא טלומרים
- ללא צנטרומר
- עם אתר ראשית הכפלה אחד
למה האחרות שגויות:
- (1) זה הפוך כמעט מכל מה שנאמר.
- (3) חיידקים אינם אורזים את ה־DNA בנוקלאוזומים כמו אאוקריוטים.
- (4) אין להם קינטוכור וספינדל כמו באאוקריוטים.
שאלה 9: נוקלאוזום
איזה מהמשפטים הבאים מתאר נכון ובמדויק את מבנה ותכונות הנוקלאוזום?
- ליבת הנוקלאוזום מורכבת מאוקטמר של היסטונים, כאשר ה־DNA מלופף סביבם ב־2.5 סיבובים הדוקים המוחזקים על ידי קשרים קוולנטיים בין סוכר ה־DNA לשיירי ליזין.
- יחידת הליבה כוללת כ־147 זוגות בסיסים של DNA המלופפים כ־1.65 פעמים סביב קומפלקס חלבוני המורכב משני הטרו־דימרים של H2A-H2B ושני הטרו־דימרים של H3-H4.
- ההיסטון H1 נקשר ל־Linker DNA ובכך הופך לחלק בלתי נפרד מהאוקטמר הליבתי, מה שמעלה את מספר זוגות הבסיסים המוגנים ל־200.
- זנבות ההיסטונים (Histone Tails) פונים אל פנים האוקטמר כדי לייצב את המבנה המרכזי, ולכן אינם נגישים למודיפיקציות אפיגנטיות לאחר יצירת הנוקלאוזום.
פתרון
התשובה הנכונה היא (2).
הנוקלאוזום בנוי מ־אוקטמר היסטונים: שני עותקים מכל אחד מ־H2A, H2B, H3, H4, וה־DNA מלופף סביבו כ־1.65 סיבובים.
למה האחרות שגויות:
- (1) H1 אינו חלק מהאוקטמר הליבתי.
- (3) דווקא ה־Linker DNA חשוף יותר.
- (4) שוב, H1 הוא linker histone, לא חלק מהליבה.
-
חלופה 1 (כימיה ומספר סיבובים): היא משתמשת במספר סיבובים שנשמע הגיוני (2.5) וטוענת לקשרים קוולנטיים. סטודנטים יודעים שיש אינטראקציה עם ליזין (חומצה אמינית חיובית), אך האינטראקציה היא אלקטרוסטטית (מטענים) ולא קוולנטית.
-
חלופה 2 (התשובה הנכונה): כוללת פירוק האוקטמר לדימרים (זו הדרך שבה הם באמת מתארגנים בביוסינתזה: H3-H4 יוצרים טטרמר, ו־H2A-H2B מצטרפים כדימרים). זה דורש מהסטודנט להכיר את המבנה הפנימי ולא רק את רשימת השמות.
-
חלופה 3 (תפקיד H1): חלופה זו מנסה לבלבל בין המושג “נוקלאוזום” (הליבה) לבין המושג “כרומטוזום” (נוקלאוזום + H1). היא טוענת ש־H1 הופך לחלק מהאוקטמר, מה שגורם לבלבול סביב מספר הבסיסים המוגנים (147 מול 166 או 200).
-
חלופה 4 (זנבות ההיסטונים): זו טעות ביוכימית קלאסית. הזנבות הם אלו שיוצאים החוצה (N-terminus tails) כדי לעבור מודיפיקציות (אצטילציה, מתילציה). מי שחושב שהם מייצבים את הליבה מבפנים יטעה כאן.
שאלה 10: נוקלאוזומים ונוקלאזות
מדוע עיכול חלקי בנוקלאזות משאיר לעיתים קרובות מקטעי DNA באורך של כ־146bp?
- כי נוקלאזות חותכות תמיד בדיוק כל 146 בסיסים
- כי 146bp הוא האורך הממוצע של טלומר אנושי
- כי ה־DNA שמלופף סביב הנוקלאוזום מוגן יחסית, בעוד שה־Linker DNA חשוף
- כי זהו האורך האופייני של רצף CpG island
פתרון
התשובה הנכונה היא (3).
ה־DNA המלופף סביב ההיסטונים מוגן יחסית, ואילו ה־Linker DNA נגיש יותר לנוקלאזות. לכן מתקבלים לעיתים מקטעים בגודל של יחידת הנוקלאוזום.
למה האחרות שגויות:
- (1) הנוקלאז לא “מודד” 146bp.
- (2) אין קשר לטלומר.
- (4) אין קשר ל־CpG islands.
שאלה 11: וריאנטים של היסטונים
איזה צימוד בין וריאנט היסטון לתפקידו הוא הנכון?
- CENP-A - סימון אזורי שבר דו־גדילי; H2A.X - הגדרת הצנטרומר
- CENP-A - הגדרת הצנטרומר; H2A.X - סימון אזור של שבר דו־גדילי
- CENP-A - יצירת טלומרים; H2A.X - דחיסת 30nm fiber
- CENP-A - השתקת כרומוזום X; H2A.X - התחלת רפליקציה
פתרון
התשובה הנכונה היא (2).
- CENP-A מחליף H3 בצנטרומר ומסמן את מיקומו
- H2A.X קשור לאזורי שבר דו־גדילי ומסמן אותם למערכת התיקון
שאלה 12: מתילציה של DNA
מהו כלל האצבע שניתן בשיעור לגבי מתילציה של DNA על CpG islands?
- מתילציה בדרך כלל קשורה לפתיחת כרומטין ולהגברת שעתוק
- מתילציה בדרך כלל קשורה לסגירת כרומטין ולהשתקה
- מתילציה מתבצעת על אדנין בלבד ולכן אינה רלוונטית באדם
- מתילציה של DNA שקולה לאצטילציה של היסטונים
פתרון
התשובה הנכונה היא (2).
לפי השיעור:
- מתילציה של DNA על CpG קשורה בדרך כלל ל־השקטה
- דה־מתילציה קשורה לפתיחה
למה האחרות שגויות:
- (1) הפוך.
- (3) בשיעור דובר על ציטוזין ב־CpG.
- (4) אלו שתי רמות רגולציה שונות.
שאלה 13: מודיפיקציות על היסטונים
איזה מהמשפטים הבאים תואם את כלל האצבע שניתן בשיעור לגבי זנבות היסטונים?
- בדרך כלל אצטילציה קשורה לכרומטין פתוח ומתילציה קשורה לכרומטין סגור
- אצטילציה ומתילציה של היסטונים משפיעות רק על יציבות הקשר הפוספודיאסטרי
- אצטילציה ומתילציה מתבצעות על ה־Histone fold המוסתר בתוך הנוקלאוזום
- אצטילציה תמיד משתיקה גנים ומתילציה תמיד מפעילה גנים ללא חריגים
פתרון
התשובה הנכונה היא (1).
זהו כלל אצבע מהשיעור, עם חריגים:
- אצטילציה ← כרומטין פתוח יותר
- מתילציה ← כרומטין סגור יותר
למה האחרות שגויות:
- (2) לא בזה מדובר.
- (3) המודיפיקציות נגישות בעיקר על הזנבות N-terminal שבולטים החוצה.
- (4) השיעור הדגיש שיש חריגים, ולכן זו קביעה קשיחה מדי.
שאלה 14: Chromatin Remodeling
איזה מהבאים הוא דוגמה לפעולה של קומפלקס chromatin remodeling?
- הוספת קבוצת מתיל לציטוזין ב־CpG
- הוספת קבוצת אצטיל לזנב היסטון
- הזזה, פינוי או הרפיה של נוקלאוזום תוך שימוש ב־ATP
- סינתזה של DNA חדש במהלך רפליקציה
פתרון
התשובה הנכונה היא (3).
קומפלקסי remodeling משתמשים ב־ATP כדי לבצע:
- sliding
- ejection
- unwrapping
- replacement
למה האחרות שגויות:
- (1) זו מתילציה של DNA.
- (2) זו מודיפיקציה קוולנטית של היסטון.
- (4) זו לא פעולת remodeling.
שאלה 15: XIST
מהו התיאור הנכון של XIST לפי השיעור?
- חלבון היסטוני שמחליף את H2A בצנטרומר
- lncRNA שנקשר לאחד מכרומוזומי X וגורם להשתקתו
- אנזים רימודלינג שמפנה נוקלאוזומים מפרומוטורים
- רצף DNA טלומרי שמגן על כרומוזום X
פתרון
התשובה הנכונה היא (2).
XIST הוא RNA לא מקודד ארוך (lncRNA) שמעורב ב־X-inactivation וגורם להשתקת אחד מכרומוזומי ה־X.
שאלה 16: סופרקוילינג במהלך טרנסקריפציה
כאשר RNA polymerase מתקדם על DNA, מה צפוי להיווצר מבחינה טופולוגית?
- לפני הפולימראז נוצר negative supercoiling, ואחריו positive supercoiling
- לפני הפולימראז נוצר positive supercoiling, ואחריו negative supercoiling
- לפני ואחרי הפולימראז נוצר תמיד אותו סוג של supercoiling
- אין השפעה טופולוגית כל עוד הפולימראז נע על גדיל יחיד בלבד
פתרון
התשובה הנכונה היא (2).
כשהפולימראז מתקדם:
- לפניו מצטבר positive supercoiling
- מאחוריו נוצר negative supercoiling
זו בדיוק הסיבה שנדרשות טופואיזומראזות לשחרור המתח.
שיעור 4: מבנה העל של הכרומטין
הטרוכרומטין ואאוכרומטין, LADs, Hi-C, TADs, קומפרטמנטים A/B, טריטוריות כרומוזומליות והמעבר מאינטרפאזה למיטוזה.
שאלה 1: אאוכרומטין מול הטרוכרומטין
איזה צימוד הוא המדויק ביותר מבחינת ארגון וסימון כימי?
- אאוכרומטין - מאופיין ב־H3K9me3 ומיקומו בפריפריית הגרעין; הטרוכרומטין - מאופיין ב־H3K4me3 ומרכזיות גיאומטרית.
- אאוכרומטין - כרומטין פתוח המאופיין ב־H3K4me3 ונגישות ל־RNA Polymerase; הטרוכרומטין - כרומטין דחוס המאופיין ב־H3K9me3 והשתקה.
- אאוכרומטין - אזור חסר נוקלאוזומים המאפשר תעתוק רציף; הטרוכרומטין - אזור שבו ה־DNA ליניארי לחלוטין ואינו מלופף סביב היסטונים.
- אאוכרומטין - כרומטין המצוי רק בקרבת ה־LADs; הטרוכרומטין - כרומטין המצוי אך ורק בתוך ה־Nucleolus לצורך השתקת גנים ריבוזומליים.
פתרון
התשובה הנכונה היא (2).
- אאוכרומטין מזוהה עם אצטילציה ומתילציה פעילה (כמו H3K4me3).
- הטרוכרומטין מזוהה עם סימני השתקה (כמו H3K9me3).
- למה האחרות שגויות:
- (1) הפוך בסימונים ובמיקומים.
- (3) בשניהם יש נוקלאוזומים.
- (4) LADs הם דווקא אזורי הטרוכרומטין.
שאלה 2: הגרעינון כדוגמה לארגון מרחבי
מה מדגים הגרעינון (Nucleolus) בהקשר של הקשר בין דחיסות לפעילות גנומית?
- שצפיפות כרומטין גבוהה במיקרוסקופיה אלקטרונית היא עדות מוחלטת להשתקה טרנסקריפציונית ולמבנה של הטרוכרומטין קונסטיטוטיבי.
- שפעילות טרנסקריפציונית גבוהה מחייבת פיזור מוחלט של הכרומטין למצב של אאוכרומטין דליל שאינו נראה כצבר (Aggregate) בגרעין.
- שאזור גרעיני מוגדר יכול להפגין דחיסות פיזית גבוהה מאוד ובכל זאת להוות אתר של פעילות טרנסקריפציונית אינטנסיבית (ייצור rRNA).
- שהגרעינון משמש כ”מחסן” להשתקת גנים ריבוזומליים עודפים, ולכן הוא נראה כצבר דחוס של כרומטין מושתק בדומה ל־LADs.
פתרון
התשובה הנכונה היא (3). הגרעינון הוא ה”יוצא מן הכלל המעיד על הכלל” - הוא נראה דחוס (Dense), אך הוא אחד האזורים הפעילים ביותר בתא.
למה האחרות שגויות:
- (1) ו־(2) הן תפיסות שגויות שהגרעינון מפריך.
- (4) הגרעינון הוא אתר ייצור, לא אתר השתקה.
שאלה 3: הטרוכרומטין קונסטיטוטיבי לעומת פקולטטיבי
איזה משפט מגדיר נכון את ההבדל התפקודי בין שני סוגי ההטרוכרומטין?
- קונסטיטוטיבי משתנה בהתאם לסיגנלים התפתחותיים (Cell-type specific); פקולטטיבי נשאר דחוס וקבוע בכל סוגי התאים באורגניזם.
- קונסטיטוטיבי מאפיין אזורים מבניים קבועים כמו צנטרומרים; פקולטטיבי מאפיין גנים שמושתקים באופן הפיך בהתאם לתוכנית הביולוגית.
- קונסטיטוטיבי נוצר רק בתגובה לנזקי DNA לצורך תיקון; פקולטטיבי הוא המצב הדיפולטיבי של כל הכרומוזומים במהלך שלב ה־S.
- קונסטיטוטיבי מאופיין בקישור של חלבוני CTCF בלבד; פקולטטיבי מאופיין בקישור בלעדי של חלבוני Lamin לממברנה הגרעינית.
פתרון
התשובה הנכונה היא (2).
- קונסטיטוטיבי: “תמיד שם” (צנטרומרים, טלומרים).
- פקולטטיבי: “לפי הצורך” (למשל השתקת כרומוזום X בנקבות או גנים ייחודיים לרקמה).
שאלה 4: LADs (Lamina-Associated Domains)
מהו התיאור המדויק ביותר של הדינמיקה והתפקיד של LADs בגרעין?
- אזורי DNA המקודדים לחלבוני הלמינה הגרעינית, ולכן הם ממוקמים פיזית בצמוד לממברנה לצורך תרגום יעיל של mRNA.
- מקטעי כרומטין, לרוב הטרוכרומטיים, המעוגנים ללמינה הגרעינית ונוטים להפגין רמות שעתוק נמוכות ודחיסות גבוהה יחסית.
- לולאות DNA פעילות מאוד הפורצות מתוך הטריטוריה הכרומוזומלית כדי להגיע לממברנה ולקבל סיגנלים חוץ־תאיים ישירים.
- אזורים המוגדרים על ידי הצטברות של חלבוני Cohesin בלבד, שתפקידם למנוע מהכרומטין לגעת בממברנת הגרעין בזמן האינטרפאזה.
פתרון
התשובה הנכונה היא (2). LADs מייצגים את ה”פריפריה המושתקת” של הגרעין.
למה האחרות שגויות:
- (1) מיקום הגן לא קשור למקום שבו החלבון פועל.
- (3) LADs הם מושתקים, לא פעילים.
- (4) Cohesin קשור ל־TADs, לא ל־LADs.
שאלה 5: מחלת ה־Progeria וארגון הכרומטין
מדוע מחלת הפרוגריה משמשת מודל חשוב להבנת ארגון הכרומטין בשיעור?
- כי היא נגרמת מחוסר מוחלט בהיסטונים, מה שמוביל לקריסה של מבנה ה־TADs בכל שלבי מחזור התא.
- כי היא מדגימה כיצד מוטציה בחלבוני הקינטוכור מונעת את הפרדת הכרומטידות האחיות בזמן המיטוזה.
- כי היא מדגימה כיצד פגם בחלבון Lamin A משבש את המבנה המרחבי של הגרעין ואת האינטראקציה התקינה של LADs עם הממברנה.
- כי היא הוכחה לכך שדחיסת כרומוזומים מיטוטית אינה תלויה ב־Condensin אלא רק באנזימי Topoisomerase II.
פתרון
התשובה הנכונה היא (3). פרוגריה נגרמת מגרסה פגומה של Lamin A (פרוגרין), שגורמת לעיוות של הגרעין ולאובדן ארגון ההטרוכרומטין הפריפריאלי.
שאלה 6: עקרון שיטת ה־Hi-C
מהו הרצף הלוגי של שלבי העבודה בשיטת Hi-C המאפשר מיפוי מרחבי של הגנום?
- בידוד כרומוזומים מטאפאזיים, צביעתם ב־DAPI, ומדידת המרחק הממוצע ביניהם תחת מיקרוסקופיה ברזולוציה גבוהה.
- ריצוף עמוק של כל ה־DNA בתא ולאחר מכן שימוש באלגוריתם המנבא את הקיפול המרחבי על בסיס רצף הנוקלאוטידים בלבד.
- קיבוע (Cross-linking) של חלבונים ל־DNA, חיתוך באנזימי רסטריקציה, וליגציה של קצוות סמוכים מרחבית ליצירת מולקולות כימריות.
- הפרדת DNA לפי גודל בג’ל אלקטרופורזה מסוג PFGE ולאחר מכן זיהוי רצפי CTCF באמצעות נוגדנים ספציפיים.
פתרון
התשובה הנכונה היא (3). המפתח ב־Hi-C הוא הליגציה: אם שני קטעי DNA שרחוקים ברצף התחברו, סימן שהם היו קרובים במרחב בזמן הקיבוע.
שאלה 7: פירוש מפת חום (Heatmap) של Hi-C
במפת Hi-C, מה ניתן להסיק מנוכחותם של אותות (נקודות חמות) הרחק מהאלכסון הראשי?
- שאזורים אלו ב־DNA עברו אמפליקציה (הכפלה) משמעותית בתאי הדגימה ולכן הם מופיעים פעמיים בריצוף.
- קיימת אינטראקציה מרחבית חזקה בין שני לוקוסים המרוחקים זה מזה מבחינה ליניארית על גבי הכרומוזום.
- מדובר בטעות טכנית בתהליך הליגציה שבה קצוות DNA חופשיים התחברו באופן אקראי לחלוטין ללא קשר למבנה הגרעין.
- אזורים אלו מייצגים DNA שאינו מלופף סביב נוקלאוזומים ולכן הוא נוטה “לנדוד” במפת החום למיקומים לא צפויים.
פתרון
התשובה הנכונה היא (2). האלכסון הראשי מייצג את ה”שכנים” ברצף. כל מה שרחוק ממנו הוא אינטראקציה ארוכת טווח (Long-range interaction).
שאלה 8: הגדרת TAD (Topologically Associating Domain)
איזה מהמשפטים הבאים מגדיר באופן המדויק ביותר את המושג TAD?
- תחום כרומוזומי שבו כל הגנים עוברים תעתוק סינכרוני על ידי אותו קומפלקס של RNA Polymerase.
- יחידה ארגונית שבה תדירות האינטראקציות המרחביות בתוך התחום גבוהה משמעותית מתדירות האינטראקציות עם אזורים מחוץ לו.
- מבנה המורכב מ־147 זוגות בסיסים של DNA המלופפים סביב שמונה חלבוני היסטונים ליבתיים.
- אזור בכרומוזום שבו ה־DNA נמצא במצב של Supercoiling חיובי המונע יצירת לולאות מרחביות.
פתרון
התשובה הנכונה היא (2). TAD הוא “שכונה” גנומית. המולקולות בפנים נפגשות הרבה יותר מאשר עם מולקולות משכונות אחרות.
שאלה 9: תפקיד ה־TADs בבקרת גנים
כיצד ארגון ה־DNA ל־TADs משפיע על בקרת הביטוי של גנים בתא?
- ה־TADs מבטיחים שכל פקטור שעתוק שנקשר ל־DNA יוכל להגיע לכל פרומוטר על פני הכרומוזום באותה יעילות.
- ה־TADs משמשים כמסגרת המגבילה את פעילות ה־Enhancers לפרומוטרים הנמצאים בתוך אותו תחום, ובכך מונעים הפעלה שגויה של גנים שכנים.
- ה־TADs מחליפים את הצורך במודיפיקציות היסטוניות ומבצעים השתקה של גנים באמצעות לחץ פיזי בלבד.
- ה־TADs גורמים לכל הגנים הנמצאים בתוכם לעבור דגרדציה מהירה של mRNA כדי לשמור על שיווי משקל תאי.
פתרון
התשובה הנכונה היא (2). ה־TAD יוצר “בידוד” (Insulation). אם ה־Enhancer והגן לא באותו TAD, הסיכוי שהם ייפגשו נמוך מאוד.
שאלה 10: מנגנון ה־Loop Extrusion (Cohesin ו־CTCF)
לפי מודל ה־Loop Extrusion, מהו התפקיד המכני של קוהסין ו־CTCF ביצירת לולאות?
- CTCF הוא מנוע מולקולרי הצורך ATP כדי לדחוף את ה־DNA, בעוד שקוהסין משמש כעוגן קבוע על הממברנה.
- קוהסין מתפקד כטבעת המגדילה את הלולאה באופן אקטיבי, בעוד ש־CTCF מתפקד כ”חסם” העוצר את התקדמות הקוהסין באתרים ספציפיים.
- CTCF מתפקד כטבעת המגדילה את הלולאה באופן אקטיבי, בעוד שקוהסין מתפקד כ”חסם” העוצר את התקדמות הקוהסין באתרים ספציפיים.
- קוהסין מחבר בין כרומוזומים לא־הומולוגיים, ו־CTCF מוודא שהחיבור יתבצע רק באזורי הטרוכרומטין.
פתרון
התשובה הנכונה היא (2). הקוהסין “פולט” לולאה (Extrusion) עד שהוא נתקע ב־CTCF. זהו המנגנון המקובל כיום ליצירת TADs.
שאלה 11: כיווניות (Orientation) של אתרי CTCF
מדוע הכיווניות של רצף הקישור של CTCF קריטית ליצירת לולאה יציבה?
- כי קוהסין יכול להיעצר בצורה אפקטיבית רק כאשר הוא פוגש חלבוני CTCF שפונים זה אל זה.
- כי אתרי CTCF שפונים לאותו כיוון גורמים לקוהסין להסתובב סביב עצמו וליצור קשרים קוולנטיים בלתי הפיכים.
- כי רק אתרי CTCF המכוונים לכיוון הצנטרומר מאפשרים שכפול תקין של ה־DNA במהלך שלב ה־S.
- כי כיווניות הפוכה (Divergent) גורמת ל־DNA להיחתך על ידי אנזימי טופואיזומראז ולהוביל לאפופטוזיס.
פתרון
התשובה הנכונה היא (1). זהו “חוק ה־Convergent”. אם האתרים לא פונים זה אל זה, הקוהסין לרוב פשוט יעבור דרכם או לא ייצור לולאה יציבה.
(Convergent orientation) = אתרי CTCF שפונים זה אל זה (← →) מאפשרים עצירה של קוהסין ויצירת לולאה יציבה.
שאלה 12: קומפרטמנטים A ו־B בגרעין
איזה מהמשפטים הבאים מתאר נכון את ההבדלים בין קומפרטמנט A לבין קומפרטמנט B?
- קומפרטמנט A מאופיין בהשתקה ומיקום מרכזי יותר בגרעין; קומפרטמנט B מאופיין בכרומטין פעיל ומיקום פריפריאלי.
- קומפרטמנט A מאופיין בכרומטין פעיל ומיקום מרכזי יותר בגרעין; קומפרטמנט B מאופיין בהשתקה ומיקום פריפריאלי.
- קומפרטמנט A מאופיין בהשתקה ומיקום פריפריאלי; קומפרטמנט B מאופיין בכרומטין פעיל ומיקום מרכזי יותר בגרעין.
- קומפרטמנט A מאופיין בכרומטין פעיל ומיקום פריפריאלי; קומפרטמנט B מאופיין בהשתקה ומיקום מרכזי יותר בגרעין.
פתרון
התשובה הנכונה היא (2). קומפרטמנטים הם רמת ארגון גבוהה יותר מ־TADs.
- A: Active (מרכז, אאוכרומטין).
- B: Bad/Blocked (פריפריה, הטרוכרומטין).
שאלה 13: דינמיות של קומפרטמנטים
מעבר של לוקוס גנטי מקומפרטמנט B לקומפרטמנט A במהלך התמיינות תאים מעיד בדרך כלל על:
- מעבר של הגן לאזור דחוס יותר בגרעין לצורך השתקה ארוכת טווח.
- איבוד של כל הנוקלאוזומים באותו אזור והפיכתו ל־DNA חשוף.
- שינוי במיקומו המרחבי של הגן ועלייה בפוטנציאל התעתוק והנגישות שלו לפקטורי שעתוק.
- שבירה פיזית של הכרומוזום וחיבורו מחדש לכרומוזום אחר (טרנסלוקציה).
פתרון
התשובה הנכונה היא (3). קומפרטמנטים הם דינמיים. כשגן צריך לעבוד, הוא “עובר דירה” מ־B ל־A.
שאלה 14: טריטוריות כרומוזומליות (Chromosome Territories)
מה נכון לגבי הארגון המרחבי של כרומוזומים שונים בגרעין האינטרפאזי?
- כל הכרומוזומים שזורים זה בזה בצורה אקראית לחלוטין כמו קערת ספגטי, ללא כל הפרדה פיזית.
- כל כרומוזום תופס נפח מוגדר (טריטוריה), כאשר כרומוזומים עשירים בגנים נוטים לתפוס מיקום מרכזי יותר בגרעין.
- שני הכרומוזומים ההומולוגיים (למשל שני עותקים של כרומוזום 1) תמיד צמודים פיזית זה לזה באותה טריטוריה.
- הטריטוריות הכרומוזומליות נקבעות במיטוזה ונשארות קבועות וזהות לחלוטין בכל התאים של אותו יצור חי.
פתרון
התשובה הנכונה היא (2). יש סדר בגרעין - כרומוזומים לא מתערבבים לגמרי. בנוסף, יש קשר בין תוכן הגנים למיקום (Gene-rich = Central).
למה האחרות שגויות:
- (3) הומולוגים בדרך כלל רחוקים זה מזה.
- (4) יש וריאביליות בין תאים.
שאלה 15: ארגון הכרומוזום במיטוזה (Mitotic Chromosomes)
מהו השינוי העיקרי המתרחש בארגון הכרומטין במעבר מאינטרפאזה למיטוזה?
- שמירה על מבנה ה־TADs והקומפרטמנטים תוך הכפלת מספר ההיסטונים בכל נוקלאוזום.
- פירוק של מבני ה־TADs והקומפרטמנטים לטובת ארגון של לולאות סביב ציר מרכזי (Axial Scaffold) המורכב מקונדנסין.
- מעבר של כל ה־DNA בגרעין לקומפרטמנט A כדי לאפשר גישה מקסימלית לאנזימי השכפול.
- הסרה מוחלטת של חלבוני ה־Lamin והחלפתם בחלבוני CTCF כדי למנוע מהכרומוזומים להידבק לממברנה.
פתרון
התשובה הנכונה היא (2). במיטוזה ה”סדר היומיומי” (TADs) מתפרק לטובת “אריזה למעבר” (כרומוזום דחוס). הציר המרכזי מבוסס על Condensin ו־Topoisomerase II.
שאלה 16: השוואה בין Cohesin ל־Condensin
מהו ההבדל המרכזי בתפקוד של קוהסין וקונדנסין לפי הנלמד בשיעור?
- קוהסין פועל בעיקר באינטרפאזה ליצירת TADs ובחיבור אחיות; קונדנסין פועל במיטוזה לצורך דחיסת הכרומוזומים.
- קוהסין פועל בעיקר במיטוזה לצורך דחיסת הכרומוזומים; קונדנסין פועל באינטרפאזה ליצירת TADs ובחיבור אחיות.
- קוהסין פועל בעיקר במיטוזה ליצירת TADs ובחיבור אחיות; קונדנסין פועל באינטרפאזה לצורך דחיסת הכרומוזומים.
- קוהסין פועל בעיקר באינטרפאזה לצורך דחיסת הכרומוזומים; קונדנסין פועל במיטוזה ליצירת TADs ובחיבור אחיות.
פתרון
התשובה הנכונה היא (1). שניהם ממשפחת ה־SMC (טבעות), אך התזמון והמטרה שונים.
שאלה 17: תפקיד החלבון Ki-67 במיטוזה
כיצד תורם החלבון Ki-67 להפרדה תקינה של כרומוזומים במהלך החלוקה?
- הוא משמש כאנזים המפרק את הקשרים בין כרומטידות אחיות באזור הצנטרומר.
- הוא יוצר מעטפת טעונה על פני הכרומוזומים המונעת מהם להיצמד זה לזה (Surfactant-like) באמצעות דחייה אלקטרוסטטית.
- הוא משמש כסיגנל המפעיל את פירוק מעטפת הגרעין בתחילת הפרופאזה.
- הוא נקשר למיקרוטובולי ומנחה אותם ישירות אל הקינטוכור של כל כרומוזום.
פתרון
התשובה הנכונה היא (2). Ki-67 פועל כמו “סבון” או שכבת מגן שמונעת מהכרומוזומים להפוך לגוש אחד דביק במיטוזה.
שאלה 18: חוסר ב־Ki-67 והשפעתו על התא
מהן התוצאות הצפויות בתא שבו הוסר החלבון Ki-67, כפי שהוסבר בשיעור?
- התא לא יצליח לבצע סגרגציה של כרומוזומים והמיטוזה תיעצר בשלב המטאפאזה.
- הכרומוזומים יעברו דגרדציה מוחלטת מיד עם הכניסה לשלב הפרופאזה בשל חשיפה לנוקלאזות.
- המיטוזה עשויה להסתיים בהצלחה, אך שחזור ארגון הכרומטין ותוכנית הטרנסקריפציה באינטרפאזה שלאחר מכן ייפגעו.
- התא יהפוך לתא סרטני באופן מיידי בשל הכפלה בלתי מבוקרת של הצנטרוזומים.
פתרון
התשובה הנכונה היא (3).
המיטוזה יכולה לעבור, אבל הבעיה היא ב”יום שאחרי” - בארגון מחדש של הגרעין בבת הצעירה.
שיעור 5: צנטרומרים, קוהזין, סגרגציה ומבוא להכפלת DNA
צנטרומרים, קוהזין, סגרגציה, גיל האם ותסמונת דאון, ההכפלה הסמי־קונסרבטיבית, ניסוי מזלסון־סטל ו־DNA polymerase.
שאלה 1: הגדרת צנטרומר פעיל
איזה ניסוח הוא המדויק ביותר לפי השיעור?
- צנטרומר פעיל נקבע רק לפי מיקום אנטומי קבוע על הכרומוזום
- צנטרומר פעיל נקבע רק לפי נוכחות רצף אלפא-סאטלייט בכל מקום שבו הוא מופיע
- צנטרומר פעיל נקבע משילוב של רצף מתאים עם הגדרה אפיגנטית הכוללת CENP-A
- צנטרומר פעיל נקבע רק לפי כמות הקוהזין המצטברת עליו במיטוזה
פתרון
התשובה הנכונה היא (3).
לפי השיעור, למרות שרצפי alpha satellite חשובים, הם לא מספיקים לבדם כדי להגדיר צנטרומר פעיל. ההגדרה היא אפיגנטית, ובמרכזה נמצא וריאנט ההיסטון CENP-A.
למה האחרות שגויות:
- (1) המיקום המורפולוגי אינו מה שמגדיר את הפעילות
- (2) alpha satellite יכול להופיע גם באזורים שאינם צנטרומר פעיל
- (4) קוהזין חשוב ליציבות ולסגרגציה, אבל אינו ההגדרה הראשונית של הצנטרומר הפעיל
שאלה 2: חשיבות צנטרומר יחיד
מה הסיבה המרכזית לכלל של “צנטרומר אחד ואחד בלבד” לכרומוזום?
- כדי לאפשר רק אתר ראשית הכפלה יחיד על אותו כרומוזום
- כדי למנוע קישור לשני כיוונים שונה ולגרום לעיכוב, שבירה או אנאופלואידיה
- כדי לשמור על סימטריה בין זרוע p לזרוע q בזמן רפליקציה
- כדי להגביל את מספר המיקרוטובולים שנקשרים לכל כרומוזום למספר קבוע
פתרון
התשובה הנכונה היא (2).
אם לכרומוזום יהיו שני צנטרומרים פעילים, הוא עלול להיקשר למיקרוטובולים בכיוונים מנוגדים, להיתקע כ־lagging chromosome, ואף להישבר או להתחלק בצורה לא שוויונית.
למה האחרות שגויות:
- (1) אין קשר ישיר למספר אתרי ראשית ההכפלה
- (3) סימטריה בין הזרועות אינה הדרישה
- (4) זו לא הבעיה המהותית, אלא הכיווניות המנוגדת של הכוחות
שאלה 3: נאוצנטרומר
איזה מצב מתאר נכון Neocentromere?
- אזור telomeric שעובר דחיסה והופך לאתר רפליקציה חדש
- רצף alpha satellite מושתק שנשאר שקט אך מושך מיקרוטובולים במיטוזה
- אזור שלא תפקד קודם כצנטרומר פעיל ועובר אקטיבציה לאחר פגיעה כרומוזומלית
- צנטרומר שני שמופעל זמנית כדי לחזק את הקינטוכור בכרומוזום גדול
פתרון
התשובה הנכונה היא (3).
השיעור הדגיש שלאחר שבר כרומוזומלי, ייתכן שיופעל neocentromere - כלומר אזור שלא שימש קודם כצנטרומר פעיל, אך מקבל תפקוד כזה.
למה האחרות שגויות:
- (1) זה לא תיאור של נאוצנטרומר
- (2) אם הוא נשאר מושתק הוא לא יתפקד כצנטרומר
- (4) התא אינו “מוסיף” צנטרומר פעיל נוסף כדי לחזק, כי שני צנטרומרים פעילים הם בעיה
שאלה 4: צנטרומר בשמרים מול אדם
איזו השוואה היא המדויקת ביותר?
- בשמרים ובאדם הצנטרומר בנוי תמיד מנוקלאוזום יחיד עם CENP-A
- בשמרים הצנטרומר פשוט וקומפקטי יותר, ובאדם הוא רחב ומורכב מחזרות alpha satellite רבות
- בשמרים הצנטרומר אפיגנטי בלבד, ובאדם הוא נקבע רק לפי רצף
- בשמרים אין כלל CENP-A, ובאדם הוא מחליף את H2A
פתרון
התשובה הנכונה היא (2).
לפי השיעור:
- בשמרים מדובר במבנה פשוט יותר
- באדם מדובר באזור גדול ומורכב, עשיר ב־alpha satellite
- בשני המקרים CENP-A משחק תפקיד מרכזי
למה האחרות שגויות:
- (1) לא נכון לגבי האדם
- (3) בשני המקרים יש רכיב אפיגנטי חשוב
- (4) CENP-A מחליף את H3, לא את H2A
שאלה 5: שתי אוכלוסיות הקוהזין
איזה ניסוח מבטא נכון את שתי האוכלוסיות של הקוהזין?
- אוכלוסייה אחת מקפלת TADs, ואוכלוסייה אחרת מחזיקה כרומטידות אחיות יחד
- אוכלוסייה אחת בונה קינטוכור, ואוכלוסייה אחרת מזרזת פולימריזציה של DNA
- אוכלוסייה אחת מגינה על טלומרים, ואוכלוסייה אחרת מסמנת שברים דו־גדיליים
- אוכלוסייה אחת מצפה כרומוזומים במיטוזה, ואוכלוסייה אחרת דוחסת כרומטין לרוזטות
פתרון
התשובה הנכונה היא (1).
השיעור הבחין בין:
- קוהזין בארגון כרומטין ו־TADs
- קוהזין שמבצע Sister Chromatid Cohesion
למה האחרות שגויות:
- בניית קיטוכור וזירוז פולימריזציה - אלו אינם תפקידי קוהזין
- הגנה על טלמורים וסימון שברים - גם אלה אינם תפקידי קוהזין
- ציפוי כרומוזומים ומיטוזה ודחיסת כרומטין לרוזטות - זה מערבב בין Ki-67 ו־Condensin
שאלה 6: מחזור הקוהזין
איזה רצף אירועים הוא הנכון ביותר?
- הקוהזין נקשר ביציבות ב־G1, מוסר ב־S, וחוזר בצנטרומר רק באנאפאזה
- הקוהזין דינמי ב־G1, מיוצב בזמן S עם יצירת הכרומטידה האחות, ומוסר בשני שלבים במיטוזה
- הקוהזין עולה רק לאחר סיום מיטוזה, ונשאר קבוע עד מחזור התא הבא
- הקוהזין נקשר רק לצנטרומר ב־S, ורק לאחר מכן מתפשט לזרועות בפרופאזה
פתרון
התשובה הנכונה היא (2).
זה בדיוק הרצף שנלמד:
- G1 - קישור דינמי
- S - ייצוב הקישור עם יצירת האחות
- מיטוזה - הסרה דו־שלבית
למה האחרות שגויות:
- (1) הופך את סדר האירועים
- (3) הקוהזין פעיל הרבה לפני סוף המיטוזה
- (4) לא זה הרצף שנלמד
שאלה 7: הסרת קוהזין במיטוזה
איזו קביעה היא המדויקת ביותר?
- בפרופאזה מוסר כל הקוהזין, ולכן כרומוזום X נעלם לפני מטאפאזה
- בפרופאזה מוסר הקוהזין מהזרועות, ובאנאפאזה separase חותך את הקוהזין שנותר בצנטרומר
- בפרופאזה separase חותך את הקוהזין בצנטרומר, ובאנאפאזה APC/C מסיר אותו מהזרועות
- בפרופאזה condensin מחליף את הקוהזין בכל האזורים, ולכן אין עוד צורך ב־APC/C
פתרון
התשובה הנכונה היא (2).
יש תהליך דו־שלבי:
- הסרה מהזרועות קודם
- חיתוך הקוהזין הצנטרומרי באנאפאזה על ידי separase, לאחר אישור APC/C
למה האחרות שגויות:
- (1) אם הכל היה מוסר מוקדם מדי, לא הייתה אחיזה בין האחיות
- (3) הסדר בין APC/C ל־separase שגוי
- (4) condensin אינו מחליף פונקציונלית את תפקיד הקוהזין הזה
שאלה 8: מנגנון ה־checkpoint של מטאפאזה־אנאפאזה
מהו הסיגנל המכני שהתא “מחפש” כדי לאשר סגרגציה?
- היעלמות מלאה של כל מיקרוטובולי הכישור מצנטרומרים לא קשורים
- יצירת רווח קטן בין הכרומטידות באזור הצנטרומר עקב משיכה דו־קוטבית מול התנגדות הקוהזין
- קיצור סימטרי של שתי זרועות הכרומוזום עד שהצנטרומר נעשה בלתי נראה
- דחיסה מרבית של הכרומטין בצנטרומר עד שלא נותרת תנועה יחסית בין האחיות
פתרון
התשובה הנכונה היא (2).
המיקרוטובולים מושכים לשני כיוונים, הקוהזין מחזיק, ונוצר רווח קטן באזור הצנטרומר. זהו הסיגנל שהקישור הביפולרי התייצב.
למה האחרות שגויות:
- (1) לא זה הסיגנל המתואר
- (3) אין “קיצור זרועות” כזה
- (4) להפך - הרווח הוא הסימן החשוב
שאלה 9: גיל האם ותסמונת דאון
איזה הסבר משקף בצורה הטובה ביותר את הקשר בין גיל האם לטריזומיה 21?
- עם הגיל גדל מספר אתרי ראשית ההכפלה על כרומוזום 21 ולכן הוא מוכפל פעמיים
- עם הגיל גובר שיעור האקטיבציה של נאוצנטרומרים בביצית ולכן כרומוזום 21 נמשך פעמיים
- עם הגיל מתמעט קוהזין פונקציונלי באואוציטים שנעצרו זמן רב, ולכן הסגרגציה במיוזה נעשית פחות מדויקת
- עם הגיל עולה קצב הרפליקציה בביצית, ולכן DNA polymerase טועה יותר דווקא בכרומוזום 21
פתרון
התשובה הנכונה היא (3).
האואוציט נשאר זמן רב מאוד במצב עצירה, וה־cohesin שהוטל עליו מוקדם צריך להחזיק שנים. עם הזמן כמות הקוהזין הפונקציונלית יורדת, ולכן גדל הסיכון ל־non-disjunction.
למה האחרות שגויות:
- (1), (2), (4) אינן משקפות את ההסבר המולקולרי שניתן בשיעור
שאלה 10: גברים לעומת נשים
איזה ניסוח הוא המדויק ביותר?
- כי בספרמטוגנזה אין כלל קוהזין, ולכן גיל האב אינו רלוונטי לאנאופלואידיה
- כי תאי הזרע עצמם נשארים עצורים במשך עשרות שנים ולכן עוברים בקרה חזקה יותר
- כי תאי הזרע מתחדשים כל הזמן, ולכן גיל תא הזרע הממוצע אינו עולה כמו באואוציט
- כי במיוזה זכרית כרומוזום 21 אינו עובר סגרגציה דרך צנטרומר וקינטוכור
פתרון
התשובה הנכונה היא (3).
בגברים יש חידוש מתמשך של תאי זרע, ולכן אין אוכלוסיית תאים שעצורה עשרות שנים עם אותו קוהזין “ישן”.
למה האחרות שגויות:
- (1) ברור שיש קוהזין גם שם
- (2) זה מתאר דווקא יותר את המצב הנקבי
- (4) הסגרגציה עדיין תלויה בצנטרומר וקינטוכור
שאלה 11: שלושת מודלי ההכפלה
איזו קביעה מבחינה נכון בין המודלים שהוצעו לפני ניסוי מסלסון ושטאל?
- במודל הקונסרבטיבי כל גדיל ישן מזווג מיד עם גדיל חדש; במודל הסמי-קונסרבטיבי הדו־גדיל הישן נשמר
- במודל הדיספרסיבי כל מולקולה מכילה מקטעים ישנים וחדשים מעורבבים; במודל הסמי-קונסרבטיבי כל מולקולה מכילה גדיל ישן וגדיל חדש
- במודל הסמי-קונסרבטיבי נוצרת מולקולה ישנה ומולקולה חדשה; במודל הדיספרסיבי כל דור מכיל רק DNA חדש
- במודל הקונסרבטיבי ובמודל הדיספרסיבי מתקבלות אותן תחזיות לאחר שני דורות ולכן אי אפשר להבדיל ביניהם
פתרון
התשובה הנכונה היא (2).
זהו ההבדל המהותי:
- דיספרסיבי - ערבוב של ישן וחדש בתוך כל מולקולה
- סמי-קונסרבטיבי - כל מולקולה עם גדיל ישן אחד וגדיל חדש אחד
למה האחרות שגויות:
- (1) מחליף בין המודלים
- (3) שוב מחליף בין המודלים
- (4) אחרי שני דורות דווקא אפשר להבדיל
שאלה 12: ניסוי מסלסון ושטאל - דור ראשון
איזו תוצאה אחרי מחזור חלוקה אחד ב־N¹⁴ הייתה עקבית עם המודל הסמי-קונסרבטיבי?
- פס כבד בלבד, כי עדיין רוב החנקן ב־DNA הוא N¹⁵
- שני פסים, כבד וקל, כי מחצית המולקולות כבר נבנו מחדש
- פס ביניים יחיד, כי כל מולקולה מכילה גדיל ישן וגדיל חדש
- פס קל בלבד, כי כל הנוקלאוטידים החדשים באים ממדיום N¹⁴
פתרון
התשובה הנכונה היא (3).
אחרי דור אחד, כל המולקולות הן היברידיות - גדיל ישן כבד וגדיל חדש קל - ולכן מתקבל פס ביניים יחיד.
למה האחרות שגויות:
- (1) לא מתאים לרפליקציה שהתרחשה
- (2) זה היה מתאים יותר לתחזית אחרת
- (4) עדיין יש גדיל ישן כבד בכל מולקולה
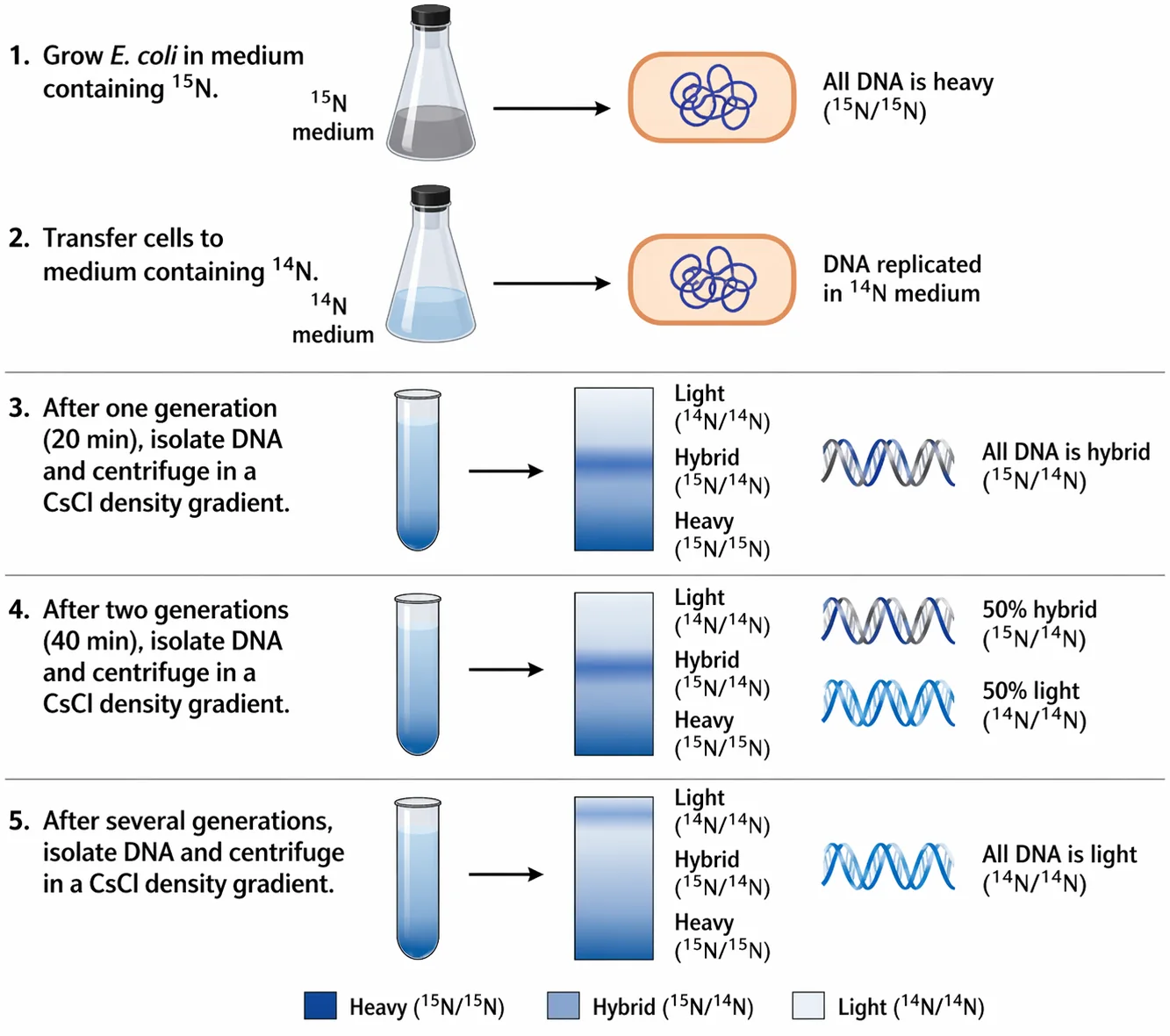
שאלה 13: ניסוי מסלסון ושטאל - דור שני
איזו תוצאה אחרי שני מחזורי חלוקה מפרידה בין סמי-קונסרבטיבי לדיספרסיבי?
- הופעת פס כבד נוסף לצד פס ביניים
- הופעת שתי אוכלוסיות: ביניים וקל
- הופעת פס יחיד שנעשה מעט קל יותר מהדור הראשון
- היעלמות מלאה של כל ה־DNA ההיברידי
פתרון
התשובה הנכונה היא (2).
אחרי שני דורות מתקבלות:
- מולקולות היברידיות
- מולקולות קלות
וזה מה שמאפשר לשלול את המודל הדיספרסיבי.
למה האחרות שגויות:
- (1) אין חזרה של אוכלוסייה כבדה טהורה
- (3) זה יותר ברוח תחזית דיספרסיבית
- (4) עדיין נותר DNA היברידי
שאלה 14: תפקיד DNA polymerase
איזה ניסוח הוא המדויק ביותר?
- הוא יוצר אנרגיה חדשה לקשר הפוספודיאסטרי שלא קיימת ב־dNTP
- הוא מקרב נכון בין 3’-OH לבין פוספט α ומבצע סלקציה פונקציונלית דרך זיווג בסיסים
- הוא מחליף את קשרי המימן בקשרים קוולנטיים בין שני הגדילים
- הוא פותח את הגדיל התבנית ומסמן היכן להתחיל את הרפליקציה
פתרון
התשובה הנכונה היא (2).
ל־DNA polymerase יש שני תפקידים מרכזיים בהקשר זה:
- קיצוב מרחבי של המגיבים
- סלקציה כך שרק זיווג נכון יוביל לשינוי קונפורמציה ולהכנסה
למה האחרות שגויות:
- (1) האנרגיה כבר נמצאת ב־dNTP
- (3) הוא לא מחבר בין שני הגדילים
- (4) זה לא תיאור מלא ונכון של תפקידו הישיר שנדון כאן
שאלה 15: כיווניות הסינתזה
איזה משפט הוא המדויק ביותר?
- DNA polymerase מסוגל לעבוד לשני הכיוונים, לפי סוג הפולימראז
- DNA polymerase קורא תבנית 5’←3’ ובונה גדיל 3’←5’
- DNA polymerase מאריך תמיד בקצה 3’ של השרשרת ולכן הסינתזה היא 5’←3’
- DNA polymerase יכול להתחיל סינתזה מאפס אם יש זיווג בסיסים תקין באזור
פתרון
התשובה הנכונה היא (3).
הפולימראז מוסיף נוקלאוטיד ל־3’-OH של הגדיל הגדל, ולכן הסינתזה היא תמיד 5’←3’.
למה האחרות שגויות:
- (1) אין polymerase שמסנתז 3’←5’
- (2) הכיווניות הפוכה
- (4) הוא לא מסוגל להתחיל שרשרת חדשה לגמרי
שאלה 16: proofreading
מתי מנגנון ההגהה של הפולימראז נכנס לפעולה?
- כאשר הנוקלאוטיד השגוי לא הצליח כלל להיכנס לאתר הקטליטי
- כאשר נוצר זיווג נכון, אך ה־pyrophosphate לא משתחרר
- כאשר נוקלאוטיד שגוי כבר הוכנס, יוצר עיוות בשלד, ומועבר לאתר 3’←5’ אקסונוקלאז
- כאשר הפולימראז נופל מה־DNA, ואז אקסונוקלאז חופשי מסיר את כל המקטע האחרון
פתרון
התשובה הנכונה היא (3).
מנגנון ההגהה פועל אחרי שהטעות כבר נכנסה. אז נוצר kink בשלד, והקצה 3’ מועבר לאתר אקסונוקלאז 3’←5’.
למה האחרות שגויות:
- (1) אם הוא לא נכנס, אין מה לתקן
- (2) זה לא המנגנון שנלמד
- (4) ההגהה מתבצעת כחלק מאותו פולימראז, לא כך
שאלה 17: איזון בין פולימריזציה להגהה
איזו תוצאה צפויה מהפרת האיזון בין שני האתרים של הפולימראז?
- אקסונוקלאז חלש מדי יאט מאוד את S phase, ואילו פולימריזציה חלשה מדי תגרום רק ליותר שגיאות
- פולימריזציה חזקה מדי עלולה להעלות טעויות, ואקסונוקלאז דומיננטי מדי עלול להאט מאוד את הרפליקציה
- כל שינוי קטן באיזון יגרום מייד להפסקה מלאה של רפליקציה בכל סוגי התאים
- פולימריזציה חזקה מדי עלולה להאט מאוד את הרפליקציה, ואקסונוקלאז דומיננטי מדי עלול להעלות טעויות
פתרון
התשובה הנכונה היא (2).
לפי השיעור:
- עודף פולימריזציה ← יותר טעויות
- עודף אקסונוקלאז ← רפליקציה איטית מאוד
למה האחרות שגויות:
- (1) מחליף בין ההשפעות
- (3) קביעה חזקה מדי
- (4) ברור שהאיזון משפיע גם על דיוק וגם על מהירות
שאלה 18: פולימראזות אנושיות שהן רֶפּלִיקָטִיבִיות עם proofreading
איזו שלישייה נכונה?
- Pol α, Pol β, Pol γ
- Pol δ, Pol ε, Pol γ
- Pol I, Pol II, Pol III
- Pol β, Pol δ, Pol κ
פתרון
התשובה הנכונה היא (2).
לפי השיעור, שלוש הפולימראזות האנושיות עם פעילות 3’→5’ exonuclease proofreading הן:
- Pol δ
- Pol ε
- Pol γ
שיעור 6: מזלג רפליקציה, טלומרים וטופואיזומרז
ראשית הכפלה, ORC/MCM, הליקאז, טופואיזומרז, RPA, PCNA, Pol α/δ/ε, מקטעי אוקזקי, החלפת פריימרים, כרומטיניזציה מחדש ו־Trinucleotide Repeat Diseases.
שאלה 1: אתר ראשית הכפלה
איזה שילוב תכונות מאפיין בצורה הטובה ביותר Origin of Replication?
- אזור עשיר ב־GC, צפוף בנוקלאוזומים, ובעל positive supercoiling מקומי
- אזור עשיר ב־AT, דל בנוקלאוזומים, ובעל תנאים שמקלים על פתיחת הדו־גדיל
- אזור אקראי בגנום שנבחר מחדש בכל מחזור על פי קצב הטרנסקריפציה
- אזור החייב להיות בתוך הטרוכרומטין קונסטיטוטיבי כדי למנוע ירי כפול
פתרון
התשובה הנכונה היא (2).
לפי השיעור, אתרי ראשית הכפלה מאופיינים ב:
- AT-rich
- Nucleosome-free
- תנאים של negative supercoiling שמקלים על פתיחת הדו־גדיל
למה האחרות שגויות:
- (1) GC-rich ו־positive supercoiling דווקא מקשים על פתיחה
- (3) המקור אינו “אקראי” כך
- (4) לא נדרש הטרוכרומטין קונסטיטוטיבי
שאלה 2: חיידקים מול אאוקריוטים - ירי אתרי הכפלה
איזה משפט מבחין נכון בין חיידקים לאאוקריוטים?
- בחיידקים ובאאוקריוטים כל Origin יכול לירות מספר פעמים באותו מחזור, אך בחיידקים יש פחות Origins
- בחיידקים יש Origin יחיד שיכול להתחיל מחזור חדש לפני סיום הקודם, ואילו באאוקריוטים כל Origin אמור לירות פעם אחת בלבד בכל מחזור
- באאוקריוטים יש Origin יחיד לכל כרומוזום, אך הוא נטען מחדש בכל שלב S מספר פעמים באותו מחזור, בעוד שבחיידקים יש Origins רבים שכל אחד מהם יורה פעם אחת בלבד
- באאוקריוטים יש Origin יחיד שיכול להתחיל מחזור חדש לפני סיום הקודם, ואילו בחיידקים כל Origin אמור לירות פעם אחת בלבד בכל מחזור
פתרון
התשובה הנכונה היא (2).
ב־E. coli יש Origin אחד ויכול להתרחש re-firing בתנאי גידול מהירים. באאוקריוטים יש Origins רבים, אבל כל Origin אמור לירות פעם אחת בלבד במחזור תא.
שאלה 3: הרכבת מזלג הרפליקציה באאוקריוטים
איזה רצף הוא המדויק ביותר?
- MCM נקשר ראשון ל־DNA, אחריו ORC, ואז RPA מפעיל את ההליקאז
- ORC מסמן את האתר, מגייס MCM לא פעיל ב־G1, ובכניסה ל־S מופעל MCM ומשתחרר לפעולה
- PCNA נטען על ה־Origin, מפעיל את ORC, ורק אז מגויסים CDC6 ו־CDT1
- ORC מפעיל מיד את Pol α, ורק לאחר תחילת הסינתזה מגיע ההליקאז MCM
פתרון
התשובה הנכונה היא (2).
לפי השיעור:
- ORC נקשר לאתר
- מגייס MCM לא פעיל ב־G1
- בכניסה ל־S מתרחשת קסקדת פוספורילציה שמפעילה את MCM
למה האחרות שגויות:
- (1) הסדר בין ORC ל־MCM הפוך, ו־RPA לא מפעיל את ההליקאז
- (3) PCNA אינו השלב הראשוני
- (4) ההליקאז חייב לפעול לפני סינתזה מתקדמת של DNA
שאלה 4: תפקידי MCM וטופואיזומראז
איזה צימוד בין חלבון לתפקידו נכון ביותר?
- MCM מסיר positive supercoiling; טופואיזומראז פותח קשרי מימן בדו־גדיל
- MCM פותח את הדו־גדיל תוך שימוש ב־ATP; טופואיזומראז משחרר את המתח שנוצר לפני המזלג
- MCM מצפה ssDNA; טופואיזומראז מחליף פריימרים במקטעי אוקזקי
- MCM מסנתז פריימרים; טופואיזומראז טוען את PCNA על ה־DNA
פתרון
התשובה הנכונה היא (2).
- MCM הוא ההליקאז שפותח את הדו־גדיל
- טופואיזומראז משחרר את ה־positive supercoiling שנוצר לפניו
שאלה 5: RPA / SSB
מהו התפקיד המרכזי של RPA במזלג הרפליקציה?
- לזהות זיווג בסיסים שגוי ולבצע proofreading בזמן אמת
- לצפות DNA חד־גדילי פתוח ולמנוע היסגרות מחודשת של הגדילים
- להחזיק את ה־DNA פולימראז צמוד לתבנית כדי להעלות processivity
- לחתוך את הפריימר כאשר הפולימראז מגיע למקטע האוקזקי הבא
פתרון
התשובה הנכונה היא (2).
RPA נקשר ל־ssDNA ומייצב אותו במצב פתוח, כדי שהבועה לא תיסגר מיד אחרי פעולת ההליקאז.
למה האחרות שגויות:
- זיהוי זיווג שגוי - זה קשור ל־proofreading של הפולימראז
- החזקת DNA פולימראז - זה התפקיד של PCNA
- חיתוך הפריימר - זה קשור ל־FEN1 או למנגנונים של החלפת פריימר
שאלה 6: PCNA
איזה משפט מתאר נכון את תרומת PCNA לרפליקציה?
- הוא מעלה fidelity על ידי חיתוך נוקלאוטידים שגויים מקצה 3’
- הוא מחזיק את הפולימראז קרוב ל־DNA וכך מעלה processivity
- הוא פותח את הדו־גדיל ומפנה נוקלאוזומים ממסלול המזלג
- הוא מסנתז את קטע ה־DNA הראשון לאחר פריימר ה־RNA
פתרון
התשובה הנכונה היא (2).
PCNA הוא sliding clamp - טבעת שננעלת על ה־DNA ומאפשרת לפולימראז רֶפּלִיקָטִיבִי להישאר צמוד לתבנית.
שאלה 7: פולימראז α
איזה תיאור של DNA polymerase α הוא המדויק ביותר?
- פולימראז רֶפּלִיקָטִיבִי מהיר עם proofreading גבוה, האחראי לשכפול רציף של leading strand
- פולימראז בעל פרוססיביות נמוכה, שמתחיל de novo, מסנתז RNA קצר ולאחריו קטע DNA קצר
- פולימראז מיטוכונדריאלי שמחליף את הפריימרים של מקטעי אוקזקי
- פולימראז תיקון שמסוגל לפרק flap ולסגור nick ללא ליגאז
פתרון
התשובה הנכונה היא (2).
Pol α:
- מתחיל de novo
- מסנתז קודם RNA primer
- ואז עוד קטע DNA קצר
- בעל processivity נמוכה וללא proofreading
שאלה 8: Leading מול lagging strand באאוקריוטים
איזה צימוד הוא הנכון ביותר?
- Pol δ - leading strand; Pol ε - lagging strand
- Pol ε - leading strand; Pol δ - lagging strand
- Pol α - leading strand; Pol δ - lagging strand
- Pol ε - שני הגדילים; Pol α - רק תיקון שלאחר רפליקציה
פתרון
התשובה הנכונה היא (2).
- Pol ε - מסנתז את ה־leading strand
- Pol δ - מסנתז את ה־lagging strand ואת מקטעי האוקזקי
שאלה 9: מקטעי אוקזקי
איזה מהמשפטים הבאים מסביר בצורה המדויקת והמנגנונית ביותר מדוע מתבצעת סינתזה מקוטעת (מקטעי אוקזקי) על הגדיל המאחר (Lagging Strand)?
- פולימראז δ, האחראי על סינתזת הגדיל המאחר, פועל ללא טבעת ה־PCNA, ולכן הפרוססיביות שלו נמוכה והוא נאלץ להתנתק מהתבנית כל 100-200 בסיסים.
- אנזימי DNA פולימראז מסוגלים להוסיף נוקלאוטידים אך ורק בכיוון 5’←3’. מכיוון שהגדילים אנטי-מקבילים, הסינתזה על הגדיל המאחר חייבת להתבצע בכיוון הפוך להתקדמות המזלג, ולכן נעשית במקטעים.
- הליקאז (MCM) פותח את קשרי המימן בפעימות, ונאלץ לעצור זמנית בכל פעם שהוא נתקל בנוקלאוזום (כל כ־200 בסיסים), מה שמכתיב את אופי הסינתזה המקוטעת.
- חלבוני ה־RPA המצפים את ה־DNA החד־גדילי חוסמים פיזית את התקדמות הפולימראז הרֶפּלִיקָטִיבִי, שמחויב להתנתק ולהמתין להנחת פריימר חדש על ידי פולימראז אלפא.
פתרון
התשובה הנכונה היא (2).
הסיבה המהותית למקטעי אוקזקי היא הכיווניות האובליגטורית של ה־DNA פולימראז (5’←3’) בשילוב עם המבנה האנטי-מקבילי של ה־DNA. כאשר המזלג נפתח, גדיל אחד יכול להסתנתז ברצף לתוך המזלג (Leading), ואילו הגדיל השני חייב להסתנתז “אחורה”, הרחק מהמזלג, ולכן מחייב התחלות חדשות (פריימרים) בכל פעם שנפתח קטע חדש.
למה האחרות שגויות:
-
(1) פולימראז $\delta$ פועל עם PCNA ויש לו פרוססיביות גבוהה. הוא עוצר ונופל לא בגלל “עייפות”, אלא כי הוא נתקל באזור דו־גדילי (הפריימר של מקטע האוקזקי הקודם).
-
(3) נוקלאוזומים אכן קובעים את גודל מקטעי האוקזקי באאוקריוטים (100-200 בסיסים), אך הם אינם הסיבה לכך שהסינתזה מתבצעת במקטעים. גם בחיידקים (ללא נוקלאוזומים) יש מקטעי אוקזקי.
-
(4) חלבוני RPA שומרים על ה־DNA כחד־גדיל, אך הם מוסרים ונדחפים על ידי הפולימראז המתקדם; הם אינם מהווים מחסום שגורם להיווצרות המקטעים.
שאלה 10: גודל מקטעי אוקזקי
מה מסביר בצורה הטובה ביותר את ההבדל בגודל מקטעי אוקזקי בין חיידקים לאאוקריוטים?
- בחיידקים אין צורך בפריימר, ולכן המקטעים גדולים יותר
- באאוקריוטים גודל המקטעים מושפע מארגון הכרומטין והנוקלאוזומים
- בחיידקים ה־PCNA קטן יותר, ולכן הפולימראז רץ מרחק גדול יותר
- באאוקריוטים ההליקאז איטי יותר, ולכן כל מקטע חייב להסתיים מוקדם יותר
פתרון
התשובה הנכונה היא (2).
השיעור קישר בין הגודל האאוקריוטי של 100-200 nt לבין גודל נוקלאוזום + linker, בעוד שבחיידקים, שאין בהם אריזה כזו, המקטעים ארוכים יותר.
שאלה 11: הפולימראז על ה־lagging strand
מה קורה כשהפולימראז על ה־lagging strand מגיע לפריימר של המקטע הבא?
- הוא ממשיך דרך ה־RNA primer ומחליף אותו ישירות ללא עצירה
- הוא נעצר מול אזור דו־גדילי, ונשאר nick שדורש עיבוד נוסף
- הוא מדלג מעל הפריימר הבא ומתחיל את המקטע שאחריו
- הוא מבצע proofreading על הפריימר ורק אז ממשיך ברצף
פתרון
התשובה הנכונה היא (2).
הפולימראז הרֶפּלִיקָטִיבִי נעצר כשהוא מגיע לאזור דו־גדילי - כלומר לאזור שבו נמצא הפריימר/המקטע הבא - ואז נשאר nick שיש להשלים.
שאלה 12: החלפת פריימר בחיידקים מול אאוקריוטים
איזה צימוד נכון?
- חיידקים - FEN1; אאוקריוטים - Pol I עם 5’→3’ exonuclease
- חיידקים - Pol I עם 5’→3’ exonuclease; אאוקריוטים - FEN1 לאחר יצירת flap
- חיידקים - Pol δ; אאוקריוטים - Pol ε
- חיידקים - RPA; אאוקריוטים - PCNA
פתרון
התשובה הנכונה היא (2).
- בחיידקים: Pol I מפרק את הפריימר מלפנים וממלא מאחור
- באאוקריוטים: נוצר flap, ו־FEN1 חותך אותו
שאלה 13: FACT לאחר מעבר מזלג הרפליקציה
איזה תיאור משקף נכון את תפקיד FACT בהקשר שנלמד?
- הוא מסיר את כל ההיסטונים לצמיתות כדי לאפשר ל־RPA לכסות את ה־DNA החדש
- הוא מסייע באריזה מחדש של ה־DNA לכרומטין מיד לאחר מעבר המזלג
- הוא מחליף CENP-A ב־H3 כדי לסמן שהרפליקציה הסתיימה
- הוא חותך את ה־linker DNA כדי להגדיר את גודל מקטע האוקזקי הבא
פתרון
התשובה הנכונה היא (2).
מיד לאחר מעבר מזלג הרפליקציה ה־DNA נארז מחדש לכרומטין, ו־FACT משתתף בתהליך הזה.
שאלה 14: Fragile X
איזה תיאור של Fragile X הוא המדויק ביותר?
- התארכות CAG באזור המקודד יוצרת חלבון polyQ רעיל
- התארכות CGG בפרומוטור גורמת להיפרמתילציה ולהשתקת FMR1
- התארכות CTG באינטרון פוגעת ב־origin firing וגורמת לפיגור שכלי
- התארכות CGG באזור הטלומרי מונעת טעינת קוהיזין על הכרומטידות
פתרון
התשובה הנכונה היא (2).
ב־Fragile X:
- הרצף החוזר הוא CGG
- הוא נמצא ב־promoter של FMR1
- חזרות רבות גורמות ל־hypermethylation
- והתוצאה היא silencing
שאלה 15: Huntington’s disease
איזה תיאור של Huntington הוא המדויק ביותר?
- חזרות CGG בפרומוטור גורמות להשתקת גן דרך מתילציה
- חזרות CAG באזור המקודד גורמות להארכת polyglutamine ולחלבון שנוטה לאגרגציה
- חזרות CTG באזור 3’UTR גורמות לאובדן קשירת PCNA
- חזרות CAG בצנטרומר מפריעות לקשירת קינטוכור ולסגרגציה
פתרון
התשובה הנכונה היא (2).
ב־Huntington’s disease:
- הרצף החוזר הוא CAG
- הוא נמצא באזור המקודד
- הוא מייצר polyglutamine expansion
- ומוביל לאגרגציה וחלבון לא תקין
שאלה 16: Myotonic dystrophy
איזו מהאפשרויות הבאות מתארת בצורה הטובה ביותר את המנגנון הכללי של Myotonic dystrophy?
- השתקת פרומוטור עקב hypermethylation של CpG islands
- יצירת חלבון polyQ רעיל עקב הרחבת CAG coding repeat
- פגיעה ב־alternative splicing עקב חזרות CTG
- כשל בטעינת MCM ולכן ירידה במספר Origins פעילים
פתרון
התשובה הנכונה היא (3).
לפי השיעור, ב־Myotonic dystrophy החזרות מובילות לפגיעה ב־alternative splicing.
שאלה 17: מנגנון ה־Slippage ברצפים חוזרניים
מהו התיאור המדויק ביותר של תופעת ה־Slippage (החלקה) במהלך רפליקציה של רצפים מסוג Trinucleotide Repeats?
- המנגנון נובע מכך ש־DNA פולימראז מאבד את האפיניות שלו ל־DNA בגלל היעדר חלבוני RPA, מה שמוביל לנפילה של האנזים וקיצור אקראי של הרצף החוזר.
- במהלך הרפליקציה מתרחשת דיסוציאציה זמנית בין הגדילים; קיפול של הגדיל החדש ללולאה יוביל להתארכות הרצף, בעוד קיפול של גדיל הטמפלייט יוביל להתקצרותו.
- מדובר בטעות של מנגנון ה־Proofreading, שבה האקסונוקלאז מזהה בטעות רצפי CAG תקינים כטעות ביוכימית ומסיר אותם שוב ושוב עד להתקצרות הגנום.
- החלקה מתרחשת אך ורק בגדיל המוביל (Leading Strand) כיוון שבו הפולימראז נע במהירות גבוהה מדי שאינה מאפשרת סגירה יציבה של קשרי המימן ברצפים חוזרים.
פתרון
התשובה הנכונה היא (2).
מנגנון ה־Slippage מתבסס על כך שברצפים חוזרים (כמו CGG או CAG), קל מאוד לגדילים להיפרד ולהתחבר מחדש ב”טעות” (Mispairing).
-
אם ה־Nascent strand (הגדיל החדש) יוצר לולאת Stem-loop, הפולימראז יסנתז שוב את אותם בסיסים והרצף יתארך (Expansion).
-
אם ה־Template strand (גדיל התבנית) הוא זה שמתקפל ללולאה, הפולימראז “ידלג” על האזור המקופל והרצף יתקצר.
למה האחרות שגויות:
-
(1) RPA דווקא עוזר למנוע מבנים שניוניים, אך Slippage קשור לדינמיקה שבין הפולימראז לגדילים ולא רק לנוכחות RPA.
-
(3) Proofreading הוא מנגנון לתיקון נוקלאוטיד בודד שגוי (Mismatch), הוא לא קשור לדילוג או הכפלה של רצפים חוזרים ארוכים.
-
(4) Slippage יכול להתרחש בשני הגדילים, ולמעשה הוא נפוץ מאוד בגדיל המאחר (Lagging) בגלל הדיסוציאציה התכופה של הפולימראז והצורך בהנחת פריימרים חדשים.
שאלה 18: סדר הפעולות הכרונולוגי במזלג הרפליקציה האאוקריוטי
איזה מהרצפים הבאים מתאר נכון את השתלשלות האירועים והשחקנים המרכזיים במזלג הרפליקציה?
- קישור ORC, טעינת והפעלת MCM, פעילות Topoisomerase ו־RPA, הנחת פריימר ע”י Pol α, קישור PCNA, סינתזה ע”י Pol ε/δ, הסרת Flaps ע”י FEN1 וחתימה ע”י Ligase.
- טעינת MCM, קישור ORC, פעילות Pol ε, הנחת פריימר ע”י Pol α, קישור PCNA, פעילות FACT, הסרת פריימר ע”י DNA Pol I וחתימה ע”י Separase.
- קישור ORC, טעינת MCM, קישור PCNA, פעילות Topoisomerase, הנחת פריימר ע”י Pol α, סינתזה ע”י Pol γ, פעילות CENP-A וחתימה ע”י Ligase.
- קישור ORC, טעינת MCM, פעילות Topoisomerase ו־RPA, הנחת פריימר ע”י Pol α, קישור Cohesin, סינתזה ע”י Pol ε/δ, פעילות FACT וחתימה ע”י APC/C.
פתרון
התשובה הנכונה היא (1).
פירוט השלבים לפי הסדר שנלמד:
- ORC: מסמן את ראשית ההכפלה ב־G1.
- MCM: ההליקאז שנטען (G1) ומופעל (שלב S) כדי לפתוח את הדו־גדיל.
- Topoisomerase ו־RPA: שחרור מתח פיתולי ומניעת סגירה מחדש של הגדילים.
- Pol α: פרימאז שמניח פריימר מעורב (RNA+DNA).
- PCNA: טבעת הצימוד (Sliding clamp) שמעלה את הפרוססיביות של הפולימראזות הרֶפּלִיקָטִיבִיות.
- Pol ε/δ: הפולימראזות שמבצעות את הסינתזה העיקרית (Leading ו־Lagging בהתאמה).
- FEN1: חיתוך ה־Flap של הפריימר הישן.
- Ligase: סגירת ה־Nick האחרון בין מקטעי אוקזקי.
למה האחרות שגויות:
-
(2) כולל את Separase (שייך להפרדת כרומטידות במיטוזה) ו־DNA Pol I (אנזים חיידקי).
-
(3) כולל את Pol γ (רפליקציה מיטוכונדריאלית) ו־CENP-A (וריאנט היסטון של הצנטרומר).
-
(4) כולל את APC/C (קומפלקס בקרת מחזור התא/מיטוזה).
שאלות נוספות - מגוון נושאים
The Structure and Function of DNA
שאלה 1: האם בתאי אדם אין כלל DNA מעגלי?
איזה מהמשפטים הבאים נכון?
- נכון, כי DNA מעגלי קיים רק בחיידקים.
- נכון, כי בכל תא אדם כל מולקולות ה־DNA הן כרומוזומים ליניאריים.
- לא נכון, כי בתאי אדם יש גם DNA מיטוכונדריאלי מעגלי.
- לא נכון, אבל DNA מעגלי מופיע רק בתאי מין.
פתרון
התשובה הנכונה היא (3).
המשפט שגוי. אמנם ה־DNA הגרעיני באדם מאורגן בכרומוזומים ליניאריים, אבל במיטוכונדריה יש DNA מעגלי.
זו מלכודת קלאסית: מי שחושב רק על הכרומוזומים בגרעין מפספס את המיטוכונדריה.
שאלה 2: הגדיל המשלים
תחילת האזור המקודד של הגן האנושי ל־β-globin היא:
5'-ATGGTGCAC-3'
איזה רצף מתאר נכון את הגדיל המשלים האנטי-פרללי?
5'-TACCACGTG-3'3'-TACCACGTG-5'3'-GTGCACCAT-5'5'-CACGTGGTA-3'
פתרון
התשובה הנכונה היא (2).
הזיווג המשלים הוא:
- A עם T
- T עם A
- G עם C
- C עם G
לכן מול 5'-ATGGTGCAC-3' נקבל:
3'-TACCACGTG-5'
שים לב: אם היו מבקשים לכתוב את הגדיל המשלים גם בכיוון 5’←3’, היית צריך לכתוב את ה־reverse complement: 5'-GTGCACCAT-3'.
אם אתה לא שולט בכיווניות, אתה תיפול גם בשכפול וגם בשעתוק.
שאלה 3: מהו ה”חומצה” ב־DNA?
כשאומרים DNA = DeoxyriboNucleic Acid, למה בעצם מתייחסת המילה Acid?
- לבסיסים החנקניים, כי הם חומציים.
- לקבוצות הפוספט בשלד הסוכר-פוספט.
- לדאוקסיריבוז, כי הוא סוכר מחזר.
- לקשרי המימן בין הבסיסים.
פתרון
התשובה הנכונה היא (2).
החומציות קשורה ל־קבוצות הפוספט שבשלד ה־DNA. בפועל, בתנאים פיזיולוגיים ה־DNA נמצא בדרך כלל במצב מנוטרל חלקית/מפורק פרוטון, ולכן הוא נושא מטען שלילי ואינו “חומצה מסוכנת” במובן היומיומי.
כלומר: כן, מקור השם הוא כימי. לא, פקיד המכס לא אמור להילחץ (בדיחה של ChatGPT).
שאלה 4: הרכב הבסיסים בווירוס M13
ב־DNA שבודד מהבקטריופאג’ M13 נמצאו:
- A = 25%
- T = 33%
- C = 22%
- G = 20%
מהי הפרשנות הסבירה ביותר?
- הנתונים בלתי אפשריים, כי בכל DNA חייב להתקיים A=T וגם G=C.
- הנתונים מרמזים שמדובר בזיהום של RNA.
- הנתונים מתאימים לכך שהגנום הוא DNA חד־גדילי.
- הנתונים מוכיחים שהדגימה עברה מוטציה נרחבת בזמן הבידוד.
פתרון
התשובה הנכונה היא (3).
ב־DNA דו־גדילי קלאסי מתקיימים בקירוב כללי צ’רגף:
- A = T
- G = C
אבל כאשר מדובר ב־DNA חד־גדילי, אין הכרח לשוויונים האלה. לכן ההרכב שנראה כאן אינו מוזר אם מדובר בגנום חד־גדילי, וזה אכן ההסבר הסביר.
זה לא פרט שולי. זו בדיוק הדרך להבחין בין עיקרון מבני לבין כלל שלא תמיד חל.
שאלה 5: כיווניות ההליקס
איזה משפט נכון לגבי הצורה ההליקלית הסטנדרטית של DNA בתאים?
- ה־DNA התאי הסטנדרטי הוא בדרך כלל הליקס ימני.
- ה־DNA התאי הסטנדרטי הוא תמיד הליקס שמאלי.
- כיוון ההליקס נקבע רק לפי אחוז ה־GC.
- אין משמעות למושג “הליקס ימני” או “שמאלי” ב־DNA.
פתרון
התשובה הנכונה היא (1).
הצורה הקלאסית והנפוצה של DNA היא B-DNA, והיא הליקס ימני. יש גם צורות אחרות, כמו Z-DNA שהוא שמאלי, אבל זו לא ברירת המחדל שעליה מדברים כשאומרים סתם “DNA”.
תשומת הלב שהליקס ימני או שמאלי לא קשור ישירות לסופר־קוילינג.
DNA תאי מוחזק בדרך כלל במצב של negative supercoiling או במצב שמעדיף פתיחה של הגדילים.
הדיוק פה חשוב:
- right-handed helix = צורת הדו־גדיל
- negative supercoiling = המצב הטופולוגי של הליפוף
שאלה 6: אחוזי הבסיסים ב־DNA אנושי
ב־DNA אנושי יש 20% ציטוזין (C) על בסיס מולרי. מה יהיו האחוזים של A, G ו־T?
- A = 20%, G = 30%, T = 30%
- A = 30%, G = 20%, T = 30%
- A = 40%, G = 20%, T = 20%
- A = 30%, G = 30%, T = 20%
פתרון
התשובה הנכונה היא (2).
אם:
- C = 20%, אז לפי כללי צ’רגף גם G = 20%
- נשארו 60% עבור A ו־T
- לכן A = 30% ו־T = 30%
בסוף זה פשוט, אבל רק אם אתה זוכר שמתחילים מ־G=C ו־A=T, ולא ממציאים מספרים מהבטן.
שאלה 7: אורך ה־DNA בתא אדם
הגנום הדיפלואידי האנושי כולל בערך 6.4 × 10^9 זוגות בסיסים. אם המרחק בין זוגות בסיסים עוקבים הוא 0.34 nm, מהו בערך האורך הכולל של ה־DNA בתא אדם אחד?
- כ־
2.2 mm - כ־
2.2 cm - כ־
2.2 m - כ־
22 m
פתרון
התשובה הנכונה היא (3).
החישוב:
6.4 × 10^9 bp × 0.34 nm/bp = 2.176 × 10^9 nm
נעביר למטרים:
2.176 × 10^9 nm = 2.176 m
כלומר, בתא אדם אחד יש בערך 2.2 מטר של DNA.
זו אחת העובדות שצריך ממש לעצור עליהן: מטרים של DNA נדחסים לגרעין מיקרוסקופי. בלי להבין את זה, אי אפשר באמת להבין למה כרומטין ואריזה הם סיפור מרכזי.
שאלה 8: איזה חלק מנפח הגרעין תופס ה־DNA עצמו?
נניח:
- אורך ה־DNA בתא אדם: כ־
2.176 m - קוטר ההליקס:
2.4 nm - קוטר הגרעין:
6 μm
אם מקרבים את ה־DNA לגליל ואת הגרעין לכדור, איזה סדר גודל מתאים לשבר הנפח שתופס ה־DNA?
- בערך
0.09% - בערך
0.9% - בערך
9% - בערך
90%
פתרון
התשובה הנכונה היא (3).
בקירוב:
- רדיוס ה־DNA:
1.2 nm - אורך ה־DNA:
2.176 m - רדיוס הגרעין:
3 μm
נפח ה־DNA כגליל: V = πr^2L
נפח הגרעין ככדור: V = (4/3)πr^3
מהחישוב מתקבל שה־DNA עצמו תופס בערך 8.7% מנפח הגרעין, כלומר סדר גודל של 9%.
זה מספר חשוב אינטואיטיבית: ה־DNA לא “ממלא לגמרי” את הגרעין כחומר מוצק, אבל הוא גם ממש לא זניח. לכן אריזה חכמה, קיפול ובקרה מבנית הם קריטיים.
שאלות נוספות - מגוון נושאים (ב׳)
DNA/כרומטין/צנטרומרים/טלומרים/סופרקוילינג. מנגנוני שכפול, טופואיזומרז, מקטעי אוקזקי, proofreading, primase ו־SSB.
שאלה 1: סופרקוילינג וטופואיזומרז
איזה תיאור משקף בצורה הטובה ביותר את הבעיה שטופואיזומרז פותר בזמן רפליקציה?
- הוא מונע יצירת קשרי מימן בין הבסיסים בשני הגדילים.
- הוא משחרר מתח טופולוגי שנוצר לפני מזלג הרפליקציה עקב פיתול־יתר של ה־DNA.
- הוא מסנתז את פריימרי ה־RNA על הגדיל המתעכב.
- הוא מייצב את הגדיל החד־גדילי במקום RPA/SSB.
פתרון
התשובה הנכונה היא (2).
ככל שההליקאז פותח את הדו־גדיל, ה־DNA שלפני המזלג נוטה להיכנס ל־positive supercoiling. בלי מנגנון שמשחרר את המתח הזה, הרפליקציה תיתקע מכנית.
טופואיזומרז לא “פותח קשרי מימן” ולא מייצר פריימרים. התפקיד שלו הוא לשבור ולחבר מחדש DNA כך שהפיתול העודף ישתחרר.
שאלה 2: האם בתא אדם יש תמיד 46 מולקולות DNA בגרעין?
איזה מהמשפטים הבאים הוא המדויק ביותר?
- נכון תמיד, כי לאדם יש 46 כרומוזומים.
- נכון רק בתאי זרע וביצית.
- לא בהכרח; זה תלוי בשלב מחזור התא, כי לאחר שכפול לכל כרומוזום יש שתי כרומטידות אחיות.
- לא נכון, כי בכל תא גוף יש 92 כרומוזומים.
פתרון
התשובה הנכונה היא (3).
המשפט “לתא אדם יש 46 מולקולות DNA בגרעין” הוא לא תמיד נכון. ב־G1 הוא מתאים: כל כרומוזום מיוצג כמולקולת DNA אחת. אבל אחרי S phase, כל כרומוזום כבר שוכפל ומכיל שתי כרומטידות אחיות, כלומר יותר מולקולות DNA בלי שינוי במספר הכרומוזומים.
שאלה 3: שלושת הרצפים המיוחדים בכרומוזום אאוקריוטי
איזה צירוף כולל את שלושת סוגי הרצפים הייעודיים הנחוצים לשמירה על מספר הכרומוזומים ומורפולוגיית הכרומוזום לאורך דורות של תאים?
- פרומוטר, אקסון, אינטרון
- Origin, enhancer, silencer
- Origin of replication, centromere, telomere
- Centromere, nucleosome, histone H1
פתרון
התשובה הנכונה היא (3).
כרומוזום אאוקריוטי תקין צריך לפחות שלושה דברים:
- origin of replication - כדי להתחיל שכפול
- centromere - כדי לאפשר סגרגציה תקינה
- telomere - כדי לשמור על הקצוות
שאלה 4: נוקלאוזומים
איזה משפט נכון יותר לגבי נוקלאוזומים?
- ברגע שנוקלאוזום הורכב על DNA, הוא לא יכול לזוז יותר.
- נוקלאוזומים נקבעים רק לפי רצף ה־DNA ולכן אין דינמיקה מבנית.
- נוקלאוזומים יכולים לשנות מיקום; האריזה אינה קפואה לחלוטין.
- נוקלאוזומים מופיעים רק בהטרוכרומטין ולא באאוכרומטין.
פתרון
התשובה הנכונה היא (3).
נוקלאוזומים אינם “מסמרים בקיר”. הם יכולים לעבור repositioning, לעבור פירוק והרכבה מחדש, ולהשתתף בדינמיקה של נגישות כרומטין.
שאלה 5: מקטעי אוקזקי
איזה משפט נכון?
- רק חלק זניח מה־DNA החדש מורכב ממקטעי אוקזקי.
- בכל סבב שכפול, בערך חצי מה־DNA החדש מסונתז תחילה כמקטעי אוקזקי.
- רק בגדיל המוביל נוצרים מקטעי אוקזקי.
- מקטעי אוקזקי נוצרים רק באאוקריוטים, לא בחיידקים.
פתרון
התשובה הנכונה היא (2).
בכל replication fork יש:
- leading strand אחד
- lagging strand אחד
כלומר, מבחינה עקרונית, בערך מחצית מה־DNA החדש נוצרת במסלול הלא־רציף של הגדיל המתעכב, ולכן מערבת מקטעי אוקזקי.
שאלה 6: DNA polymerase
מה צפוי לקרות אם DNA polymerase יאבד את פעילות ה־3’←5’ exonuclease שלו?
- קצב הסינתזה יעלה מעט או לא ישתנה מאוד, אבל הדיוק ירד משמעותית.
- קצב הסינתזה ייעצר לחלוטין, אך הדיוק לא ייפגע.
- הדיוק דווקא יעלה, כי האנזים לא “יתעכב” על תיקון.
- לא תהיה השפעה, כי mismatch repair מתקן הכול בכל מקרה.
פתרון
התשובה הנכונה היא (1).
פעילות proofreading אחראית להסרת נוקלאוטידים שהוכנסו בטעות. בלי הפעילות הזו:
- fidelity יורד
- הסינתזה עצמה לא בהכרח נעצרת
שאלה 7: רמת הדיוק (Fidelity) של הפרימאז
מהו ההסבר הביולוגי המדויק ביותר לכך שהתא “מתפשר” על רמת דיוק נמוכה והיעדר מנגנון הגהה (Proofreading) בפעילות ה־DNA פולימראז אלפא (פרימאז)?
- טעויות בפריימר אינן קריטיות מכיוון שאתרי ראשית ההכפלה (Origins) נמצאים תמיד באזורים שאינם מקודדים לחלבון (Intergenic regions).
- קצב הסינתזה של הפרימאז כה גבוה (מעל 1,000 נוקלאוטידים לשנייה), עד שהאנרגיה הדרושה להגהה הייתה מעכבת משמעותית את תחילת השכפול.
- הפרימאז נדרש ליכולת התחלת שרשרת De Novo; מכיוון שהפריימר הוא תוצר זמני שמוסר ומוחלף ב־DNA מדויק על ידי פולימראזות רפליקטיביות, אין צורך אבולוציוני להשקיע בתיקון טעויות בו.
- חלבוני ה־RPA המצפים את הפריימר מונעים פיזית מהאקסונוקלאז לגשת אליו, ולכן התא אינו יכול לבצע הגהה על מקטעי RNA-DNA היברידיים.
פתרון
התשובה הנכונה היא (3).
הסבר מנגנוני: האנזים DNA פולימראז אלפא (באאוקריוטים) הוא היחיד שיכול להתחיל לסנתז ללא קצה $\ce{3’-OH}$ חופשי. עם זאת, המחיר של בניית שרשרת מאפס הוא רמת דיוק נמוכה. התא “מרשה לעצמו” להשתמש באנזים כזה כי הוא ממילא תופס את הפריימר כמולקולה זמנית. בסוף התהליך, אנזימים כמו FEN1 מסירים את ה־Flap של הפריימר, והפולימראז הרפליקטיבי ($\delta$) – שהוא בעל יכולת הגהה ודיוק גבוה בהרבה – ממלא את הפער.
למה האחרות שגויות:
-
(1) פריימרים מונחים לאורך כל הגנום (במיוחד בגדיל המאחר), כולל בתוך אקסונים של גנים, ולכן מיקום ה־Origin אינו רלוונטי.
-
(2) להפך, לפרימאז יש פרוססיביות נמוכה מאוד והוא נופל מהר מה־DNA (לאחר כ־20 עד 30 נוקלאוטידים).
-
(4) RPA אינו מונע הגהה; הסיבה להיעדר ההגהה היא היעדר אתר פעיל של אקסונוקלאז $\ce{3’ \leftarrow 5’}$ באנזים פולימראז אלפא עצמו.
שאלה 8: תפקיד חלבוני RPA / SSB
מדוע קישור חלבוני RPA (באאוקריוטים) או SSB (בחיידקים) חיוני לתקינות תהליך השכפול?
- הם פועלים כמנוע מולקולרי המפרק באופן אקטיבי קשרי מימן מלפני הפולימראז תוך ניצול אנרגיה מהידרוליזת ATP.
- הם מצפים את ה־DNA החד־גדילי כדי למנוע היווצרות מבנים שניוניים (כמו Hairpins) וסגירה מחדש של הדו־גדיל, מה שמאפשר רציפות של הפולימראז.
- הם משמשים כעוגן פיזי המגייס את ה־PCNA (Sliding Clamp) אל הפריימר כדי להעלות את הפרוססיביות של DNA פולימראז אלפא.
- הם מזהים פגמים בקיפול ה־DNA ומגייסים באופן אקטיבי את הטופואיזומראז לצורך שחרור מתח פיתולי חיובי (Positive Supercoiling).
פתרון
התשובה הנכונה היא (2).
התפקיד המרכזי של RPA/SSB הוא לייצב את ה־DNA במצב חד־גדילי (ssDNA). ללא הגנה זו, ה־DNA נוטה להיסגר חזרה לדו־גדיל או להתקפל למבנים פנימיים (Hairpins) שחוסמים פיזית את התקדמות הפולימראז ועלולים להוביל לעצירת המזלג ולשברים.
למה האחרות שגויות:
-
פירוק קשרי מימן בעזרת ATP - זהו התפקיד של ההליקאז (MCM). RPA אינו מפרק את הדו־גדיל אלא רק שומר עליו פתוח לאחר שההליקאז עבר.
-
(3) ה־PCNA מגויס על ידי טוען הקלאמפ (RFC) לנקודת ה־Primer-Template Junction, ולא על ידי RPA.
-
(4) הטופואיזומראז פועל לרוב לפני המזלג (על הדו־גדיל) או מאחוריו, והוא מגיב למתח פיתולי ולא לקישור של RPA על החד־גדיל.
להלן שאלות תרגול חדשות בפורמט אקדמי, המבוססות על הסגנון המופיע בספר הלימוד שצירפת (שאלות מנגנוניות, פיזיקליות ותיאורטיות) ועל החומר המולקולרי המתקדם שלמדת.
שאלות נוספות - רפליקציה
שאלה 1: מגבלות פיזיקליות של מזלג הרפליקציה
קצב ההתקדמות של מזלג הרפליקציה ב־E. coli הוא כ־\(1,000\) נוקלאוטידים לשנייה. ידוע כי כל סיבוב של ה־DNA (Double Helix) מורכב מ־\(10.5\) זוגות בסיסים.
הנח כי פעילות הטופואיזומראז מעוכבת לחלוטין. מה יקרה למהירות התקדמות המזלג ככל שההליקאז ממשיך לפתוח את הגדילים? הסבר את המנגנון הפיזיקלי.
- המזלג יאיץ כיוון שהמתח הפיתולי החיובי (Positive Supercoiling) מסייע לשבירת קשרי המימן בין הגדילים.
- המזלג ייעצר במהירות כיוון שהצטברות פיתולי-על מלפני המזלג יוצרת התנגדות טורקית (Torque) הגוברת על הכוח המכני של ההליקאז.
- המזלג ימשיך כרגיל אך ה־DNA יעבור דנטורציה ספונטנית באזורים רחוקים מהמזלג כדי לשחרר את הלחץ.
- המזלג יעבור לסינתזה חד־גדילית בלבד כדי למנוע את הצורך בסיבוב של הדו־גדיל סביב עצמו.
פתרון והסבר
התשובה הנכונה היא (2).
כאשר ההליקאז מפריד את הגדילים, הוא חייב לסובב את הדו־גדיל מלפניו. ללא טופואיזומראז שישחרר את המתח (על ידי יצירת שברים זמניים), הצטברות של Positive Supercoiling תיצור לחץ פיזיקלי כה עצום שהאנרגיה המופקת מהידרוליזה של ATP על ידי ההליקאז לא תספיק כדי להתגבר עליו, והמזלג “יינעל” וייעצר.
שאלה 2: פרדוקס הדיוק של ה־DNA פולימראז
בספר הלימוד (שאלה 5-17) מועלית הסברה כי אנזים המסוגל להתחיל סינתזה De Novo (כמו פרימאז) הוא בהכרח פחות מדויק מאנזים שדורש פריימר.
מדוע מנגנון ה־Self-correction (הגהה) אינו מאפשר סינתזה מהבסיס הראשון?
- כי אתר האקסונוקלאז דורש נוכחות של חלבוני RPA כדי לזהות טעויות, ו־RPA לא נקשר לנוקלאוטיד בודד.
- כי זיווג הבסיס הראשון והשני לטמפלייט הוא חלש ופחות יציב מבחינה אנרגטית; מנגנון הגהה יעיל היה מזהה יציבות נמוכה זו כ”טעות” ומסיר את הבסיסים ללא הרף.
- כי פרימאז משתמש ב־Ribonucleotides, ומנגנון ה־Proofreading של ה־DNA פולימראז יודע לעבוד רק עם Deoxyribonucleotides.
- כי ה־Sliding Clamp (PCNA) אינו יכול להיסגר על ה־DNA עד שיש לפחות 50 נוקלאוטידים מסונתזים.
פתרון והסבר
התשובה הנכונה היא (2).
מנגנון ה־Proofreading מבוסס על היכולת של האנזים לחוש אם הנוקלאוטיד האחרון שנוסף יצר זיווג בסיסים יציב ומדויק. בתחילת שרשרת, הקשרים בין הנוקלאוטידים הראשונים לטמפלייט הם חלשים מטבעם (מעט קשרי מימן). אם לפולימראז היה מנגנון הגהה פעיל בשלב זה, הוא היה מסיר את הבסיסים מיד כיוון שהם “מרגישים” לא יציבים, והסינתזה לעולם לא הייתה מתחילה.
שאלה 3: גאומטריה של מפגש מזלגות רפליקציה
כאשר שני מזלגות רפליקציה הנעים בכיוונים מנוגדים (משני Origins שונים) נפגשים על אותו כרומוזום:
איזה מהתיאורים הבאים מתאר נכון את נקודת המפגש?
- גדיל מוביל (Leading) של מזלג אחד יפגוש את הגדיל המאחר (Lagging) של המזלג השני.
- שני הגדילים המובילים יתנגשו זה בזה ויגרמו לעצירה קבועה של הרפליקציה עד להתערבות של הליקאזות מיוחדות.
- שני הגדילים המאחרים יתאחדו למקטו אוקזקי ארוך במיוחד שאינו דורש ליגאז.
- המזלגות עוברים זה לצד זה וממשיכים לסנתז עד לקצה הכרומוזום.
פתרון והסבר
התשובה הנכונה היא (1).
מכיוון ששני המזלגות נעים זה לכיוון זה על אותה מולקולת DNA אנטי-מקבילית, הגדיל המשמש כטמפלייט לסינתזה רציפה (Leading) במזלג אחד, יהיה הגדיל המשמש כטמפלייט לסינתזה מקוטעת (Lagging) במזלג הבא מולו. בנקודת המפגש, הגדילים יתחברו זה לזה (בעזרת ליגאז) ליצירת מולקולה רציפה אחת.
שאלה 4: “צלקות” גנומיות ושגיאות רפליקציה (קליני)
בהמשך לדיון בספר על נזקי DNA מצטברים (שאלה 5-13), ידוע כי תהליך ה־Slippage נפוץ במיוחד ברצפים חוזרניים.
מדוע מחלות מסוג Trinucleotide Repeat (כמו Fragile X) נוטות להחמיר מדור לדור (תופעת ה־Anticipation)?
- כי ה־DNA פולימראז הופך ליותר “עצלן” ככל שהתא מתבגר ומפסיק להשתמש ב־PCNA.
- כי לולאות ה־Stem-loop שנוצרות בגדיל החדש במהלך ה־Slippage מובילות להרחבת הרצף; ככל שהרצף ארוך יותר, הסיכוי ליצירת לולאות נוספות ברפליקציה הבאה עולה.
- כי מערכת ה־Mismatch Repair (MMR) מכוילת לזהות רק טעויות בחיידקים ולא בתאי אדם.
- כי רצפים חוזרים מושכים אליהם יותר טופואיזומראזות שגורמים לשברים כפולים ב־DNA.
פתרון והסבר
התשובה הנכונה היא (2).
זוהי שאלה המשלבת את המנגנון המולקולרי עם ההשלכה הקלינית. ככל שרצף החזרות (כמו CGG) ארוך יותר, גדלה ההסתברות שבמהלך פתיחת הגדילים ברפליקציה, הגדיל המסונתז “יחליק” וייצור מבנה שניוני של לולאה. הפולימראז יסנתז מחדש את האזור המקופל, ובכך יאריך את הרצף עוד יותר בכל מחזור תא (או במעבר בין דורות בתאי מין).