פתיחה - חזרה על השיעורים הקודמים
השיעור נפתח בחזרה קצרה על NGS וספריות DNA דרך שאלות Kahoot. הנקודות שעלו בחזרה:
-
היתרונות של NGS לעומת Sanger כוללים ריצוף תוך כדי סינתזה, ריצוף מקבילי ומהירות. שימוש ב־reads ארוכים יותר אינו יתרון טיפוסי של NGS לעומת Sanger.
-
סלקציה שלילית של rRNA נעשית בעזרת RNase H, שמזהה RNA בתוך היבריד DNA-RNA.
-
תיקון קצוות של מקטעי DNA לפני חיבור אדפטורים קשור להכנת המקטע לחיבור האדפטור.
-
אם מתקבלים מקטעים קטנים מדי בספרייה, הבעיה קשורה ל־Size selection, ובפרט לשימוש לא מתאים ב־carboxyl beads.
-
Genome indexing נועד להפוך את החיפוש בגנום ליעיל יותר.
-
האתגר הייחודי במיפוי RNA-seq הוא gapped reads, בגלל שחבור: ה־read מגיע מ־cDNA, אבל רפרנס DNA גנומי כולל אינטרונים.
-
Protein A משמש לצימוד ביד מגנטי לנוגדן, דרך הקישור לחלק הקבוע של הנוגדן.
-
הרזולוציה של ChIP-seq תלויה בגודל המקטעים בספרייה: ככל שהמקטע קטן יותר, קל יותר למקם את אתר הקישור המדויק.
מכאן עברנו לנושא המרכזי של החלק הזה בקורס: נזקי DNA ואי־יציבות גנומית.
אי־יציבות גנומית - למה זה חשוב?
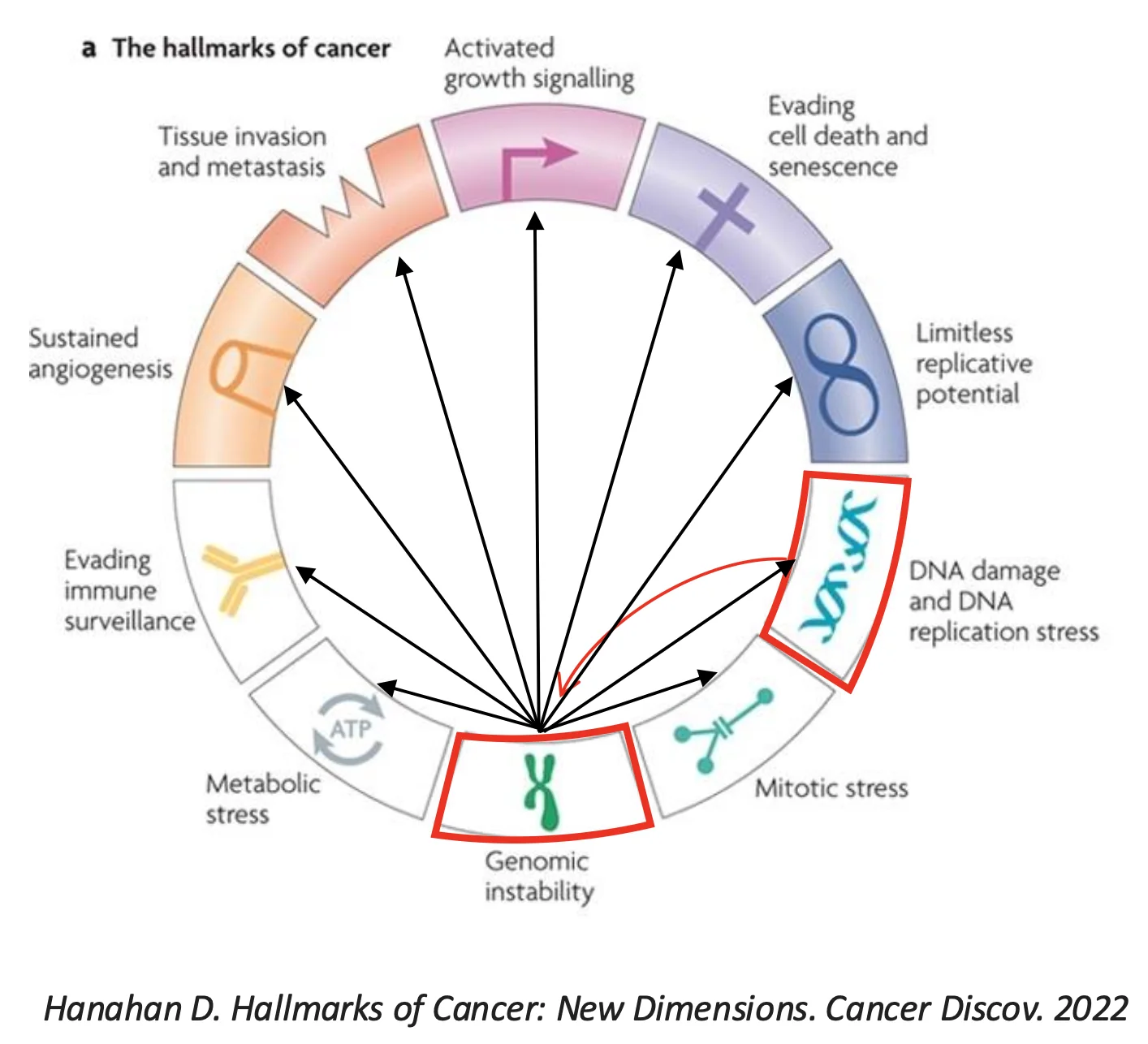
אי־יציבות גנומית קשורה ישירות לפתולוגיות שונות, ובעיקר לסרטן. מתמונה מתוך המאמר The Hallmarks of Cancer עולה, כי נזקי DNA ואי־יציבות גנומית מופיעים כחלק ממאפייני התא הסרטני.
יש להבחין בין שני מושגים קרובים:
- DNA damage - נזק שמתרחש בזמן אמת במולקולת DNA, כתוצאה מגורם מסוים.
- Genome instability - תוצר אפשרי של נזקי DNA: שינויים כרומוזומליים ארוכי טווח שיכולים לעבור לתאי הבת.
כלומר, נזק ל־DNA לא בהכרח הופך מיד לשינוי קבוע. ברוב המקרים הנזק מתוקן. הוא הופך לבעיה יציבה יותר כאשר הוא שורד את שכפול ה־DNA. אחרי השכפול, השינוי כבר יכול לעבור לתאי הבת.
אי־יציבות גנומית כמאפיין-על בסרטן
אי־יציבות גנומית יכולה להיחשב מאפיין-על, משום שהיא מאפשרת לתא לרכוש תכונות סרטניות נוספות.
כדי שתא ירכוש תכונה חדשה, כמו חלוקה מוגברת, התחמקות מאפופטוזיס או התחמקות ממערכת החיסון, נדרש שינוי גנטי כלשהו: מוטציה, מחיקה, טרנסלוקציה או שינוי כרומוזומלי אחר.
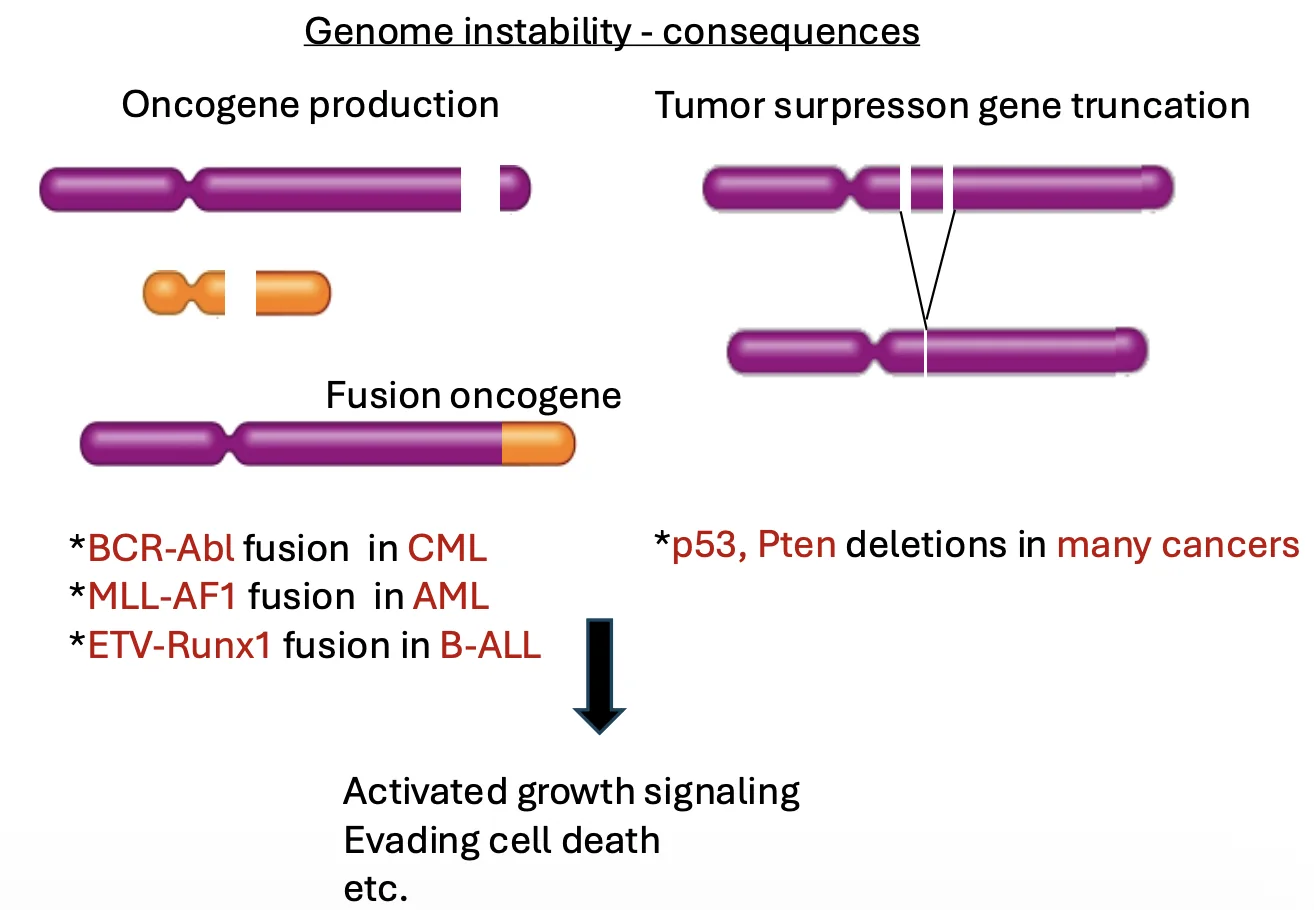
דוגמאות לתוצרים של אי־יציבות גנומית:
- יצירה או הפעלה של אונקוגן - גן שהתוצר שלו מקדם תכונות סרטניות, למשל חלוקה מוגברת או מניעת אפופטוזיס.
- פגיעה ב־tumor suppressor gene - גן שתפקידו למנוע התפתחות סרטנית. כאשר הוא נמחק או עובר מוטציה, התא מאבד מנגנון הגנה.
דוגמה מרכזית היא p53: זהו tumor suppressor חשוב, שמזהה נזקי DNA ויכול להוביל את התא לאפופטוזיס. לכן, בתאים סרטניים יש יתרון באיבוד או פגיעה ב־p53: התא מאבד את ״השומר״, ויכול לצבור עוד ועוד שינויים.
הזדקנות
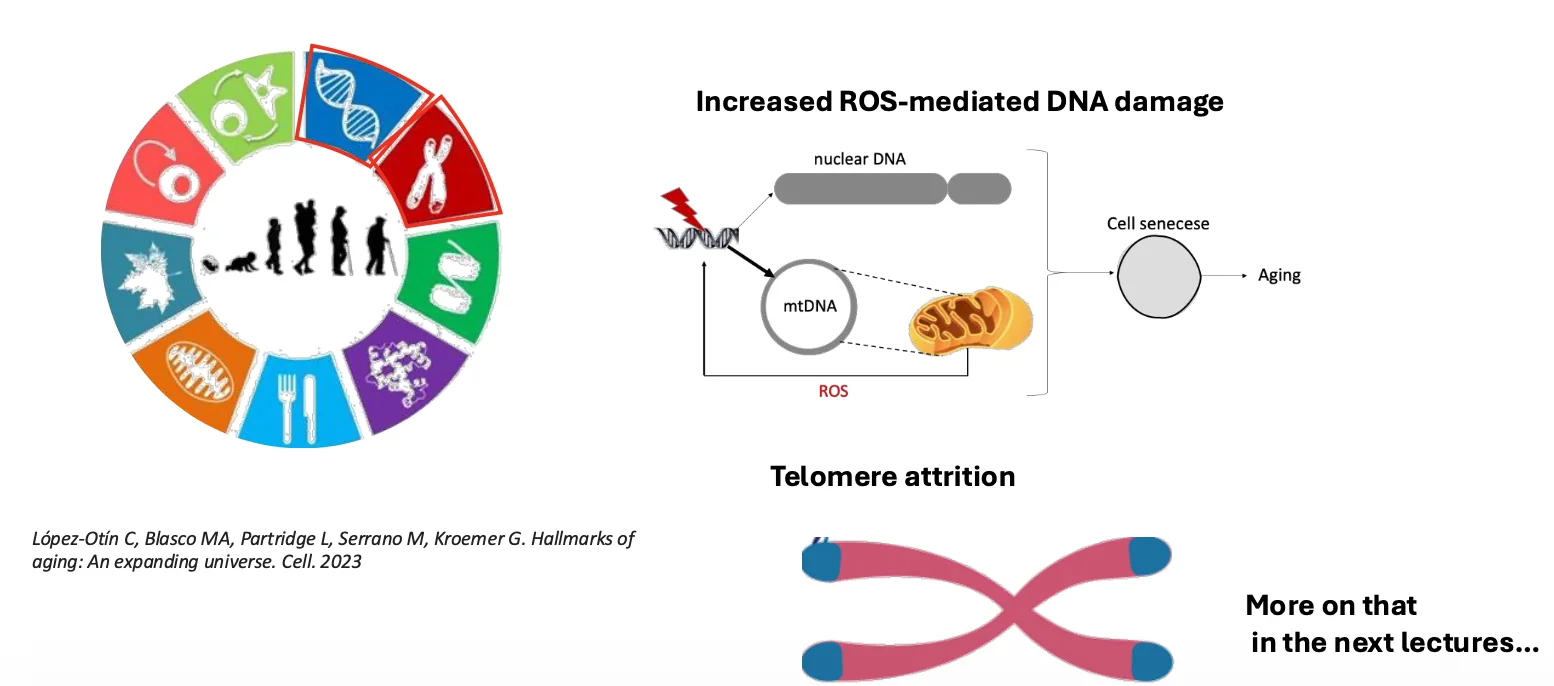
אי־יציבות גנומית קשורה גם להזדקנות. אחד המקורות לנזקי DNA במהלך ההזדקנות הוא המיטוכונדריה.
המיטוכונדריה מייצרת ATP, אבל כתוצר לוואי נוצרים גם Reactive Oxygen Species (ROS). ככל שהמיטוכונדריה מזדקנת ונעשית פחות יעילה, יש פחות יצירת ATP ויותר יצירת ROS.
ROS יכולים להגיב עם ה־DNA וליצור נזקים. כך נוצר מעגל שבו הזדקנות מיטוכונדריאלית מגבירה נזקי DNA, ונזקי DNA תורמים להזדקנות תאית.
נזקי DNA קשורים גם לשחיקת טלומרים - תהליך שמתרחש גם באופן טבעי עם הגיל, אבל נזקי DNA יכולים להגביר את הקצב שלו.
מחלות גנטיות וטיפולים אנטי-סרטניים
הוזכרו גם מחלות שבהן הבעיה היא פגם במנגנוני תיקון DNA, למשל Xeroderma Pigmentosum (XP), וכן דוגמאות כמו BRCA בהקשרים של תיקון נזקי DNA.
מצד שני, הרבה טיפולים אנטי-סרטניים דווקא משתמשים במכוון בהשריית נזקי DNA. הרעיון הוא לגרום מספיק נזק כך שהתא הסרטני ימות. לעיתים הטיפול משלב השריה של נזקי DNA יחד עם פגיעה ביכולת של התא לתקן את הנזק.
הגורמים לנזקי DNA
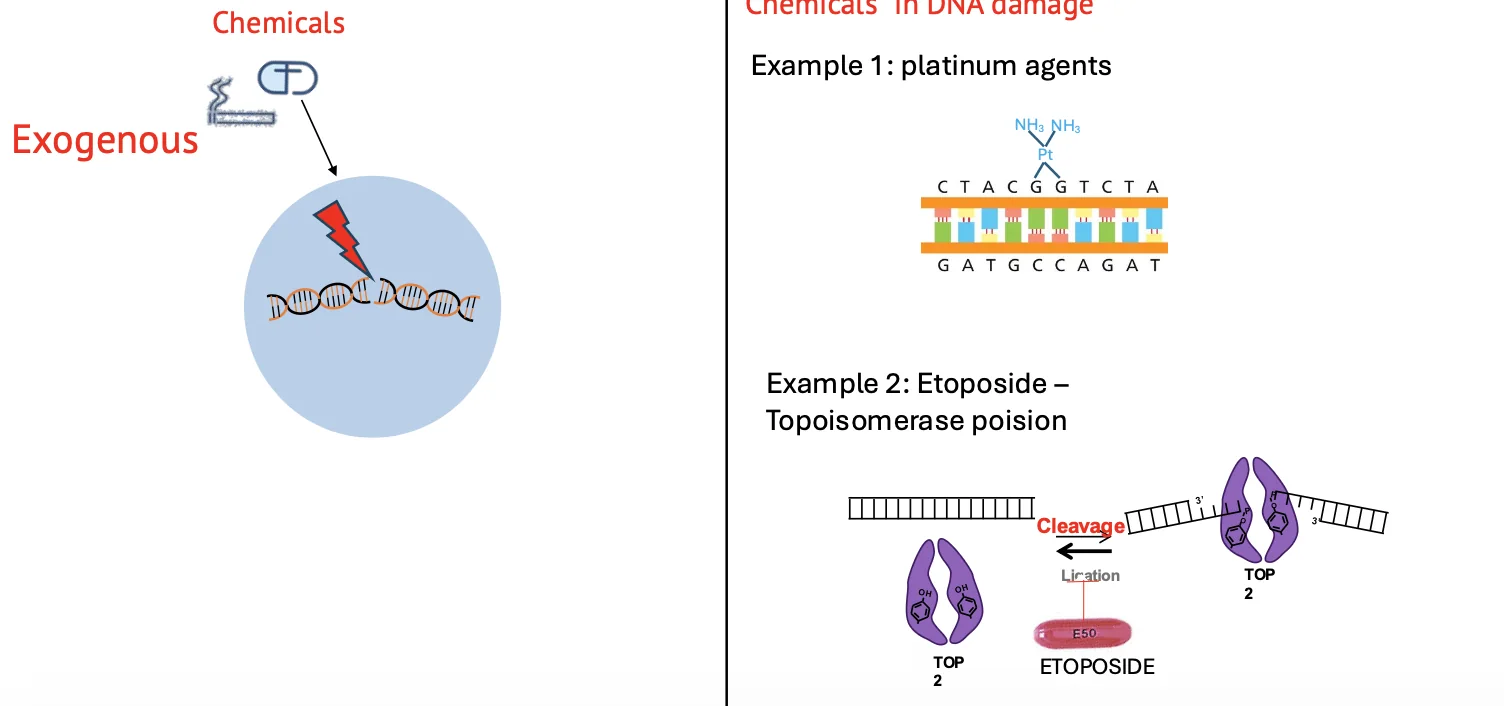
נזקי DNA יכולים להיגרם מגורמים חיצוניים או מגורמים פנימיים.
גורמים חיצוניים
גורמים חיצוניים כוללים כימיקלים וקרינה.
Platinum agents הם דוגמה לכימיקלים שמגיבים עם DNA ויוצרים נזקים גדולים יותר מבחינה מבנית.
קרינה היא גורם חיצוני נוסף:
- UV radiation יכולה ליצור דימרים של תימין.
- Ionizing radiation יכולה לגרום לנזקים שונים, כולל שברי DNA.
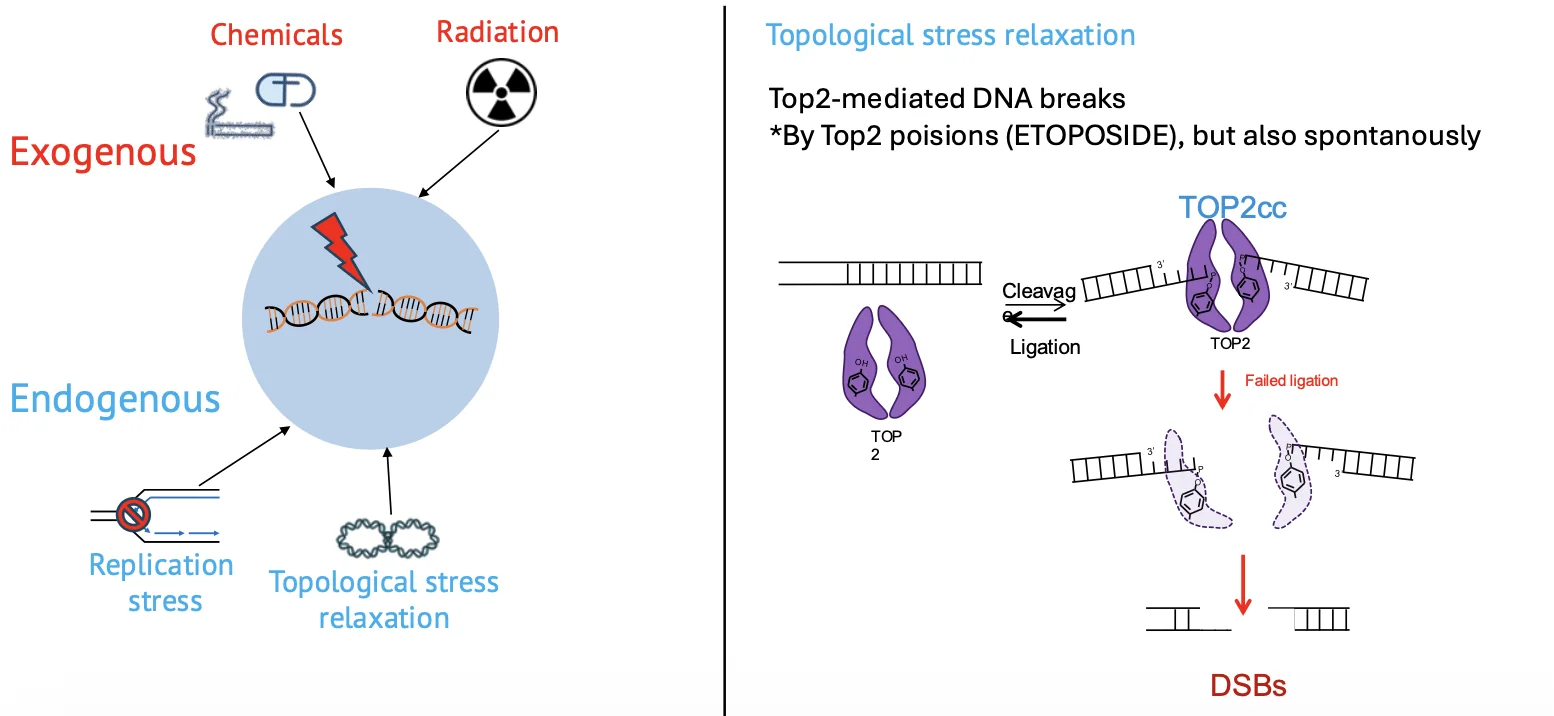
הוזכר גם Toposide, כתרופה אנטי-סרטנית שפועלת דרך פגיעה בפעילות של topoisomerase. העיקרון הוא שעיכוב או הרעלה של topoisomerase יכולים להוביל ליצירת נזקי DNA.
גורמים פנימיים
גם בלי גורמים חיצוניים, DNA יכול להינזק. התא עצמו מייצר מצבים שבהם ה־DNA מאוים.
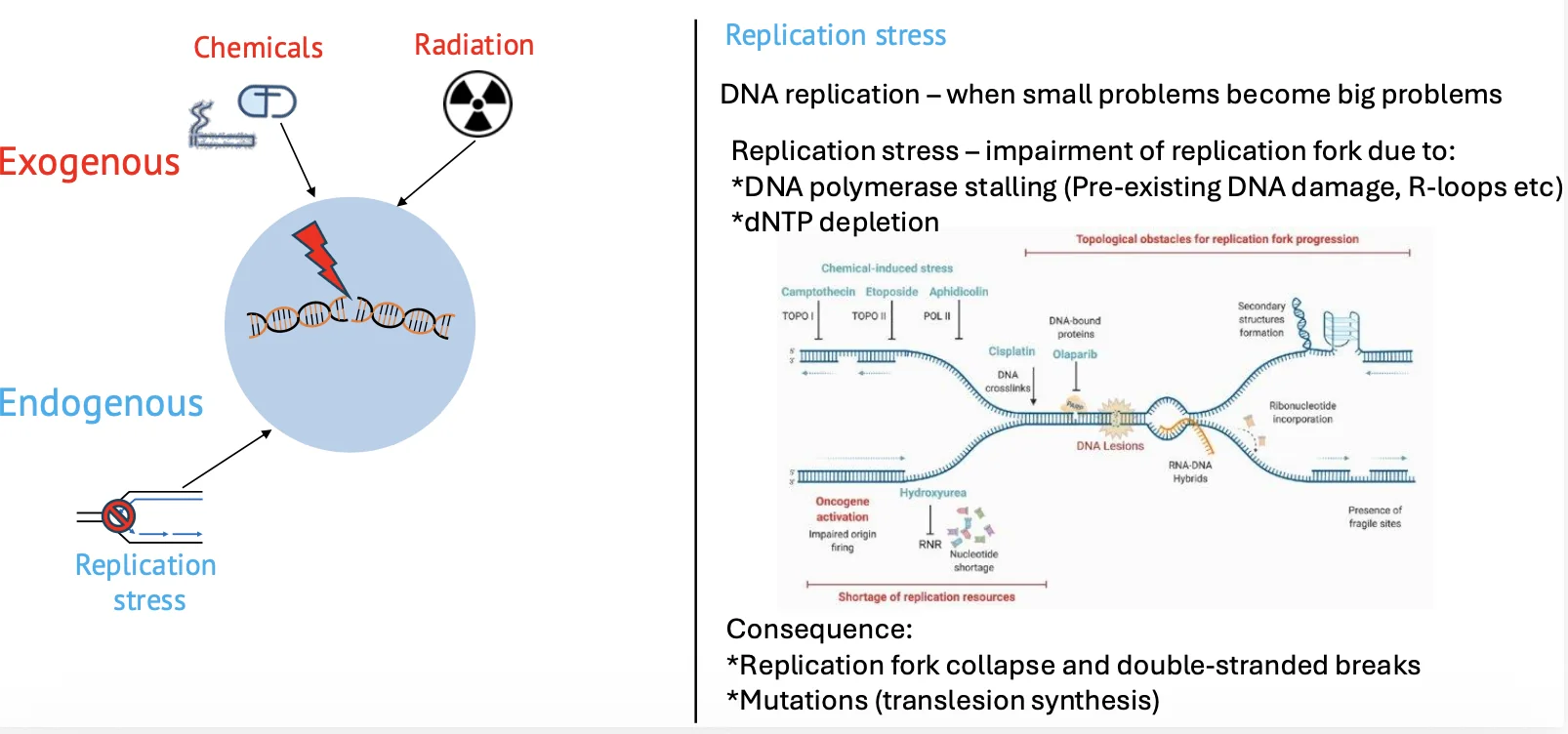
שכפול DNA
אחת מנקודות התורפה המרכזיות היא שכפול DNA. מזלג השכפול צריך ״כביש נקי״ כדי להתקדם. אם יש נזק, מבנה שניוני או מכשול אחר, מזלג השכפול יכול להיתקע.
שכפול DNA הוא המקום שבו בעיות קטנות יכולות להפוך לבעיות גדולות. כאשר מזלג השכפול קורס, נוקלאזות יכולות לחתוך את ה־DNA כדי לשחרר את המבנה התקוע, וכך נוצרים שברי DNA.
Topoisomerase
גם פעילות תקינה של topoisomerase מאתגרת את היציבות הגנומית. Topoisomerase II משחרר מתח טופולוגי על ידי יצירת שבר זמני והפיך ב־DNA, מעבר של מקטע DNA וחיבור מחדש.
בדרך כלל זה תהליך מבוקר ומהיר, אבל מכיוון שהוא מתרחש בתדירות גבוהה, יש סיכוי סטטיסטי שמשהו ישתבש והחיבור מחדש לא יקרה כראוי.
תהליך יצירת נוגדנים
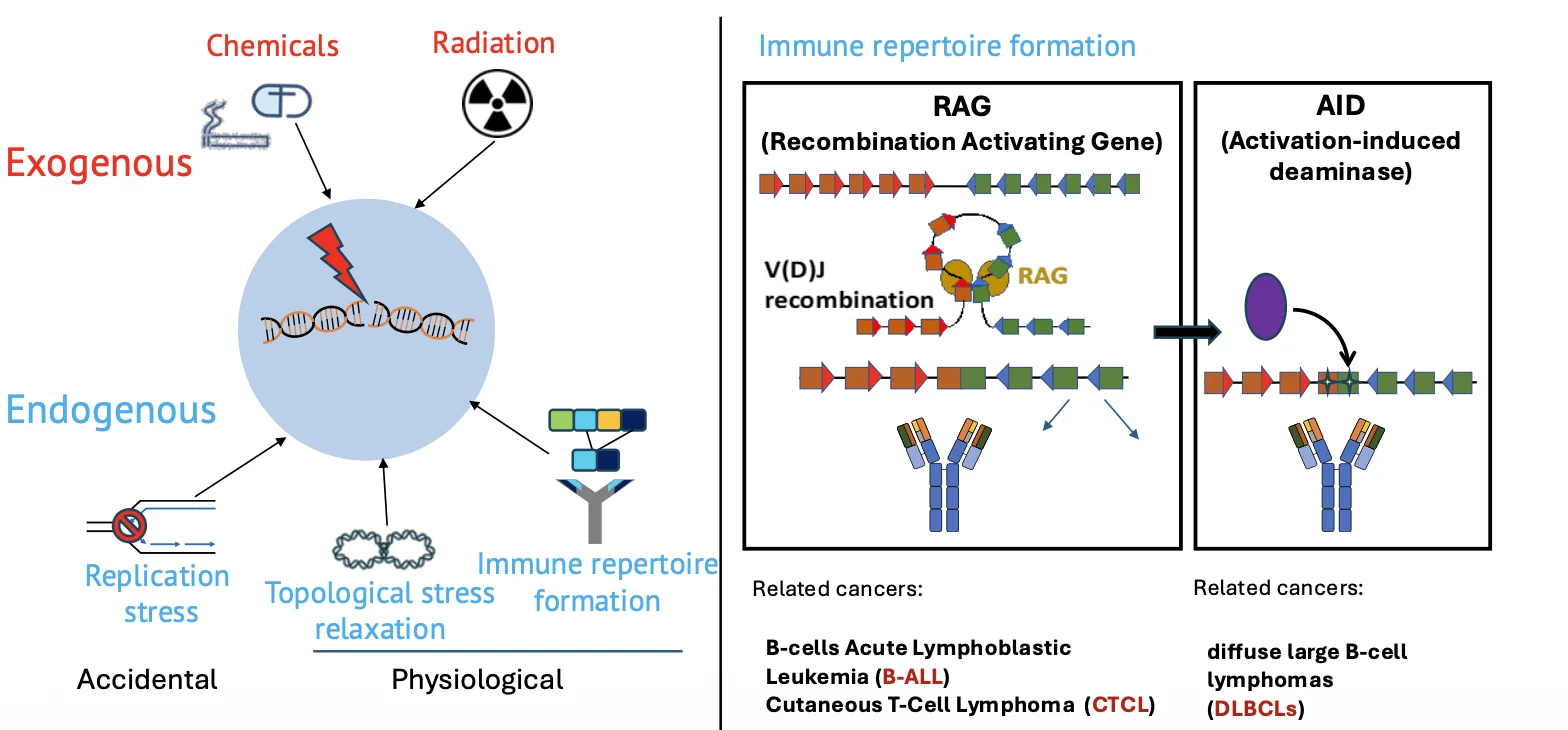
דוגמה נוספת היא יצירת נוגדנים ורצפטורים לאנטיגן. כדי ליצור מגוון עצום של נוגדנים ושל T-cell receptors, תאי מערכת החיסון מבצעים עריכה של DNA.
ברמת ה־DNA, זה דומה ל־cut and paste: חותכים את ה־DNA ומחברים אותו מחדש כדי ליצור רצפים שונים. זה הכרחי ליצירת מערכת חיסון אדפטיבית מגוונת, אבל זה גם מסוכן, כי מדובר ביצירת שברי DNA באופן יזום.
המנגנון אמור להיות מבוקר מאוד, אבל הוא לא מושלם. לכן יש לו גם קשר להתפתחות סרטני דם, למשל לימפומות.
Translesion synthesis - סבילות לנזק במקום תיקון
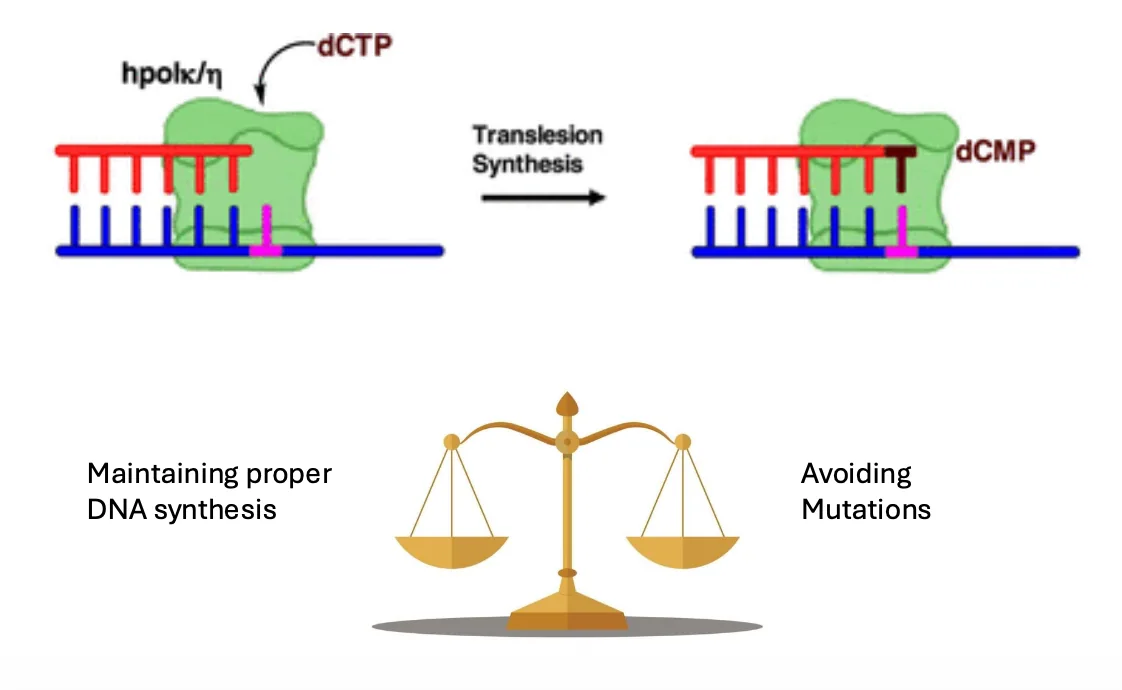
לא תמיד התא מתקן את הנזק לפני שהוא ממשיך; לפעמים הוא ״סובל״ את הנזק כדי לא לעצור את השכפול. זה נקרא Translesion synthesis.
במנגנון הזה התא מסנתז DNA מעל נזק קיים. היתרון הוא שהשכפול יכול להמשיך והתא יכול לשרוד. החיסרון הוא שזה מנגנון פחות מדויק, ולכן הוא יכול להכניס מוטציות.
זה אחד ההסברים לקשר בין נזקי DNA לבין מוטציות שעוברות לתאי הבת: אם התא משכפל מעל נזק במקום לתקן אותו, הוא עלול לקבע מוטציה.
Translesion synthesis הוא אינו מנגנון למניעת מוטציות.
שינויים כרומוזומליים
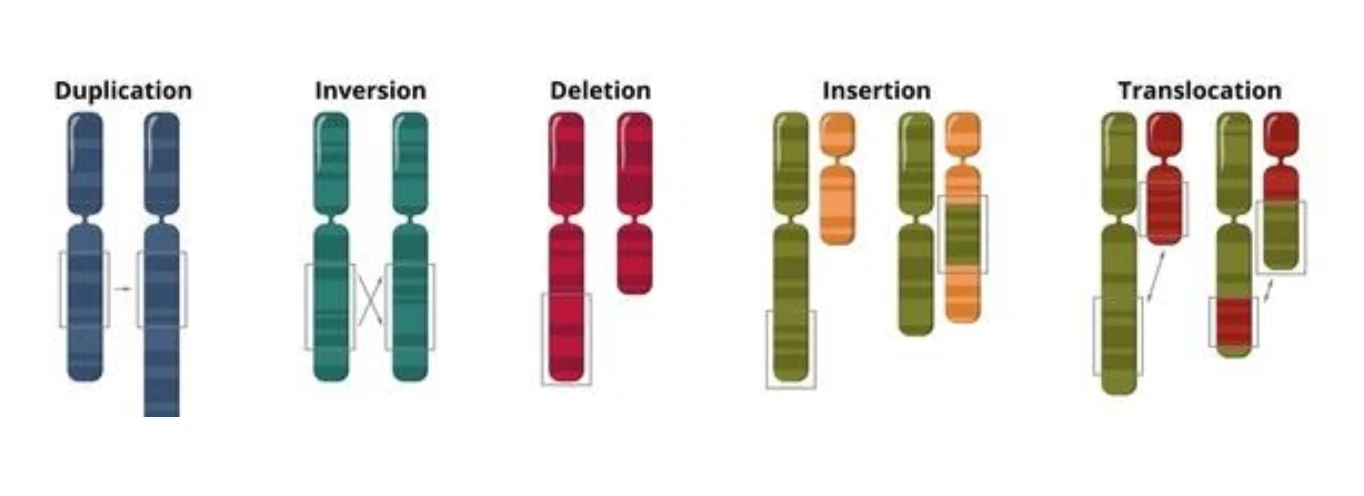
תוצר אפשרי של נזקי DNA ואי־יציבות גנומית הוא chromosomal aberrations - שינויים כרומוזומליים.
לדוגמה:
- מחיקות
- דופליקציות
- אינסרציות
- טרנסלוקציות
הוזכרו גם transposable elements ו־transposase, כמרכיבים שמקורם האבולוציוני קשור ל־DNA ויראלי ושיכולים לגרום לבלגן גנומי אם אינם מבוקרים.
זיהוי נזקי DNA במעבדה
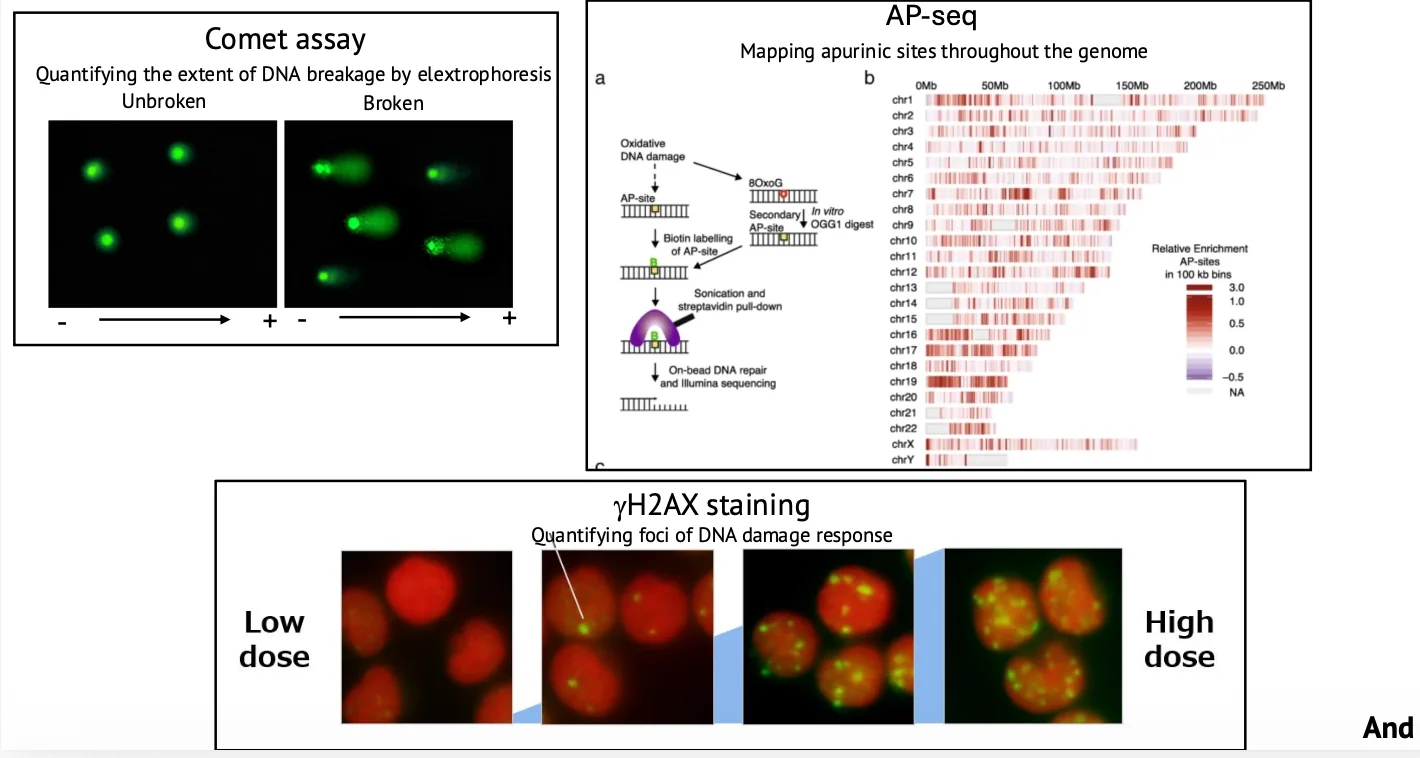
אחת הדרכים לזהות שברי DNA היא Comet assay. בבדיקה זו צובעים DNA בצביעה פלואורסצנטית ומריצים באלקטרופורזה. אם ה־DNA שלם, הוא נשאר יותר כגוש עגול. אם יש שברים, המקטעים הקטנים נודדים ויוצרים ״זנב״ שנראה כמו כוכב שביט.
ככל שהזנב גדול או אינטנסיבי יותר, יש יותר נזקי DNA.
הוזכרו גם דרכים נוספות לזיהוי נזקים, כמו ספריות DNA שמזהות שברים או נזקים אחרים, צביעה של γH2AX לזיהוי אזורים הקשורים לתיקון שברי DNA, ומיפוי של אתרים חסרי בסיס.
נזקי DNA ברמת הבסיס
בחלק העיקרי של השיעור נתמקד ב־Base-level DNA damage או Non-bulky DNA lesions. אלו נזקים שבהם שלד הסוכר-פוספט נשאר שלם, אבל הבסיס עצמו נפגע, משתנה או נעלם. זה שונה מ־bulky lesions, שבהם נזק גדול יותר משנה את המבנה המרחבי של ה־DNA.
שלושת הנזקים המרכזיים:
- דה־פורינציה (Depurination) - איבוד בסיס פוריני
- דה־אמינציה (Deamination) - הסרת קבוצת אמין
- חמצון/אוקסידציה (Oxidation) - חמצון בסיס, בעיקר גואנין
| סוג הנזק | בסיסים שנפגעים | מה קורה? | מנגנון תיקון |
|---|---|---|---|
| דה־פורינציה | פורינים: A ו־G | איבוד הבסיס ויצירת אתר חסר בסיס | BER |
| דה־אמינציה | בעיקר C | הסרת קבוצת אמין, למשל C→U | BER |
| אוקסידציה | בעיקר G | יצירת 8-oxoG | BER |
דה־פורינציה (Depurination)
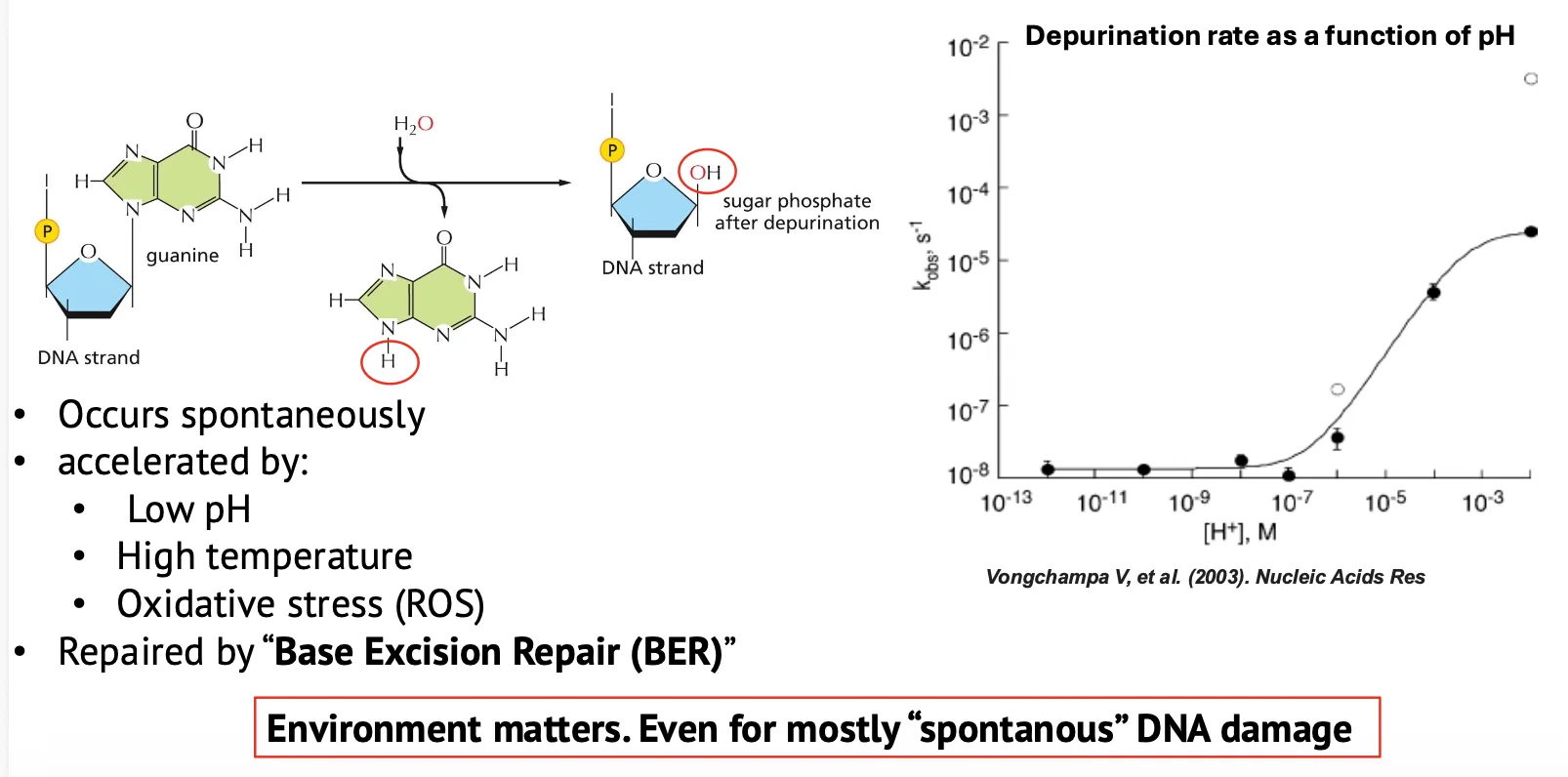
דה־פורינציה היא איבוד של בסיס פוריני (A או G). האיבוד מתרחש על ידי הידרוליזת קשר ה־N-glycosidic, שמחבר בין הבסיס לסוכר. לאחר איבוד הבסיס נשאר שלד סוכרי-פוספטי בלי בסיס. אתר כזה נקרא AP site (״אתר א-פוריני״, או באופן כללי: ״אתר חסר בסיס״).
(P)─OCH₂ O OH
│ ╱ ╲ ┆
│╱ H H ╲┆
│▚ │ │ ▞│
│ ▚├▄▄▄▄▄▄▄▄▄▄┤▞ │
H │ │ H
OH H
AP site: P + Suger
דה־פורינציה שכיחה הרבה יותר מדה־פירימידינציה, כי הקשר בין פורין לסוכר חלש יותר מהקשר המקביל בפירימידינים.
- דה־פורינציה (Depurination): כ־18,000 מקרים ביום, בערך 5 בשנייה.
- דה־פירימידינציה (Depyrimidination): כ־600 מקרים ביום.
דה־פורינציה יכולה לקרות באופן ספונטני, כי יש מים ותנאים שמאפשרים הידרוליזה. היא מוגברת בתנאים של pH נמוך ושל טמפרטורה גבוהה. היא יכולה להיות מושפעת גם מ־oxidative stress.
דה־אמינציה (Deamination)
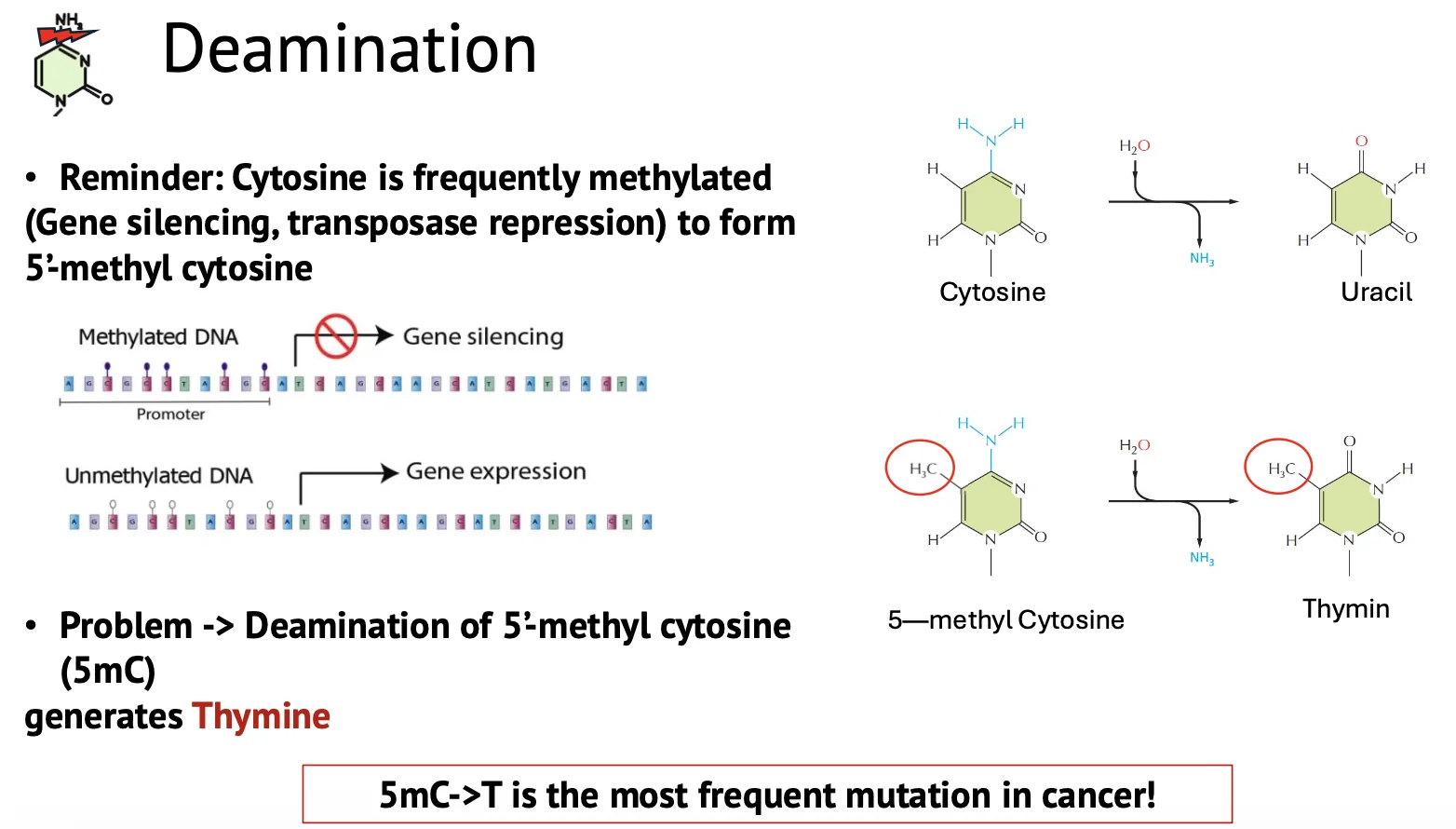
דה־אמינציה (Deamination, לעיתים גם נכתב ״דה־אמינציה״), היא הסרת קבוצת אמין מהבסיס. הדוגמה המרכזית היא: C->U (ציטוזין הופך לאורציל). זה פחות שכיח מדה־פורינציה (כ־100 מקרים ביום).
כאן מופיעה אחת הסיבות לכך שב־DNA יש תימין (T) ולא אורציל (U): אם ציטוזין עובר דה־אמינציה והופך לאורציל, התא יכול לזהות שאורציל לא אמור להיות ב־DNA ולתקן אותו.
אבל יש מקרה בעייתי יותר: ציטוזין יכול לעבור מתילציה ל־5-methylcytosine (5mC). אם 5mC עובר דה־אמינציה, תוצר הדה־אמינציה שמתקבל הוא תימין ולא אורציל: 5mC->T. זו בעיה, כי תימין הוא בסיס תקין ב־DNA, ולכן קשה יותר לתא לזהות שמדובר בטעות. המוטציה 5mC->T היא אחת המוטציות הנפוצות בסרטן.
דה־אמינציה יכולה לקרות באופן ספונטני, וגם היא מוגברת בתנאים כמו pH נמוך וטמפרטורה גבוהה. בנוסף לכך, יש אנזימים שעושים דה־אמינציה באופן פעיל.
מקומות שבהם מתיליציה נפוצה, כמו פרומוטורים שעברו מתיליציה, הם מקומות שבהם נצפה למצוא את המוטציות
C->Tבשכיחות גבוהה.
Base Excision Repair (BER)
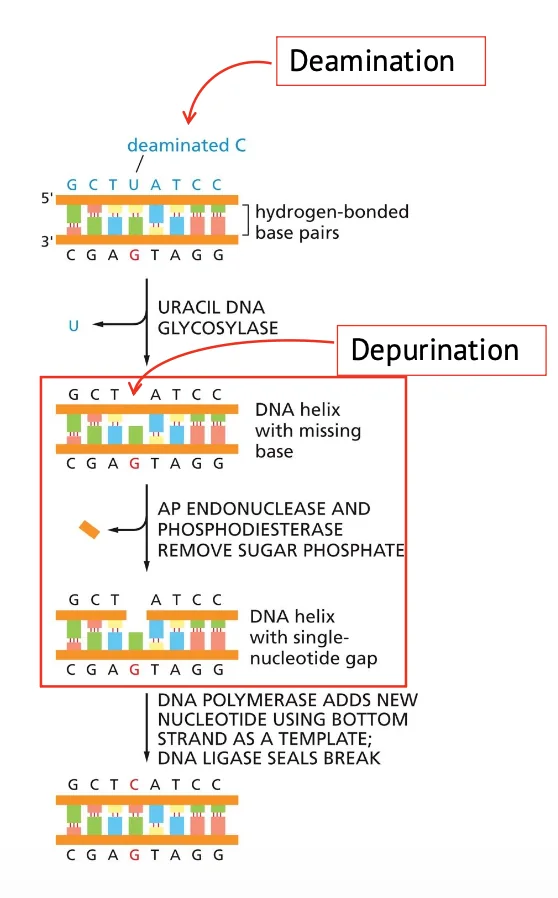
Base Excision Repair (BER) הוא מנגנון התיקון המרכזי לנזקים קטנים ברמת הבסיס. המנגנון עובד בשלושה שלבים כלליים שחוזרים גם במנגנוני תיקון אחרים:
- Recognition - זיהוי הנזקאו
- Processing - עיבוד הנזק והכנת האתר לתיקון
- Repair - השלמת הנוקלאוטיד החסר וסגירת השלד
השלב הראשון - יצירת אתר חסר בסיס
במקרה של דה־פורינציה, כבר יש אתר חסר־בסיס (כי הבסיס נעלם).
במקרה של דה־אמינציה, קודם צריך לזהות את האורציל ולהסיר אותו. האנזים שעושה זאת הוא: UNG (Uracil DNA Glycosylase). ה־UNG מזהה אורציל בתוך DNA, מסיר אותו, וכך יוצר אתר חסר בסיס (כמו ״שן חסרה״ ב־DNA).
הסרת הסוכר החסר בסיס
לאחר שנוצר AP site, צריך לחתוך את חלק הסוכר-פוספט חסר הבסיס (אי אפשר פשוט ״להכניס בסיס״ לתוך הסוכר הקיים - צריך להכניס לשם נוקלאוטיד שלם).
האנזים שעושה את החיתוך הוא: APE1 (AP endonuclease); ה־APE1 מזהה AP site וחותך את השלד, כך שנוצר nick חד־גדילי.
השלמת הנוקלאוטיד וסגירת השרשרת
לאחר מכן:
- DNA polymerase β מוסיף את הנוקלאוטיד החסר.
- ליגאז סוגר את ה־nick ומחבר מחדש את שלד הסוכר-פוספט.
לדימוי:
- פולימראז β הוא כמו פינצטה שמניחה את הנוקלאוטיד במקום.
- ליגאז הוא כמו דבק שסוגר את השרשרת.
מה קורה אם לא מתקנים בזמן?
״הטוב, הרע והמכוער״:
- הטוב - BER מתקן בזמן, והבסיס התקין משוחזר.
- הרע - אם אתר חסר בסיס עובר שכפול לפני תיקון, יכולה להיווצר באזור הזה מחיקה.
- המכוער - Translesion synthesis יכול לסנתז מעל האתר הפגוע בצורה פחות מדויקת, וכך להכניס מוטציה.
גם בדה־אמינציה, אם לא מתקנים בזמן, שינוי כמו C→U עלול להפוך לאחר שכפול לשינוי קבוע בזוג הבסיסים.
דה־אמינציה יזומה במערכת החיסון ובתגובה אנטי-ויראלית
אחרי שהוצגה דה־אמינציה כנזק, עולה השאלה: למה שהתא יעשה דה־אמינציה באופן יזום?
AID בתאי B
במערכת החיסון, תאי B צריכים לשפר את אפיניות הנוגדנים שלהם לאנטיגן. התהליך נקרא Affinity maturation. כדי לעשות זאת, נוצרים שינויים בגנים המקודדים לנוגדנים. המוטציות נוצרות באופן אקראי, ואז יש סלקציה לתאים שבהם הנוגדן נקשר טוב יותר לפתוגן.
האנזים המרכזי הוא: AID (Activation-Induced Deaminase). AID עושה דה־אמינציה לציטוזין (דה־אמינציה על פירימידין כשלב ביניים), ולאחר מכן מגייס פולימראז error-prone, במקום פולימראז β, ליצירת רמות גבוהות של מוטציות באזורי הנוגדנים (תהליך somatic hypermutation).
AID יוצר שינוי שמוביל למנגנוני תיקון פחות שמרניים, וליצירת מוטציות באזורי הנוגדנים.
זה חשוב ליצירת תגובה חיסונית טובה יותר, אבל יש לזה גם צד מסוכן: אנזים שמייצר מוטציות ב־DNA עלול לתרום לסרטני דם, כמו לימפומות.
APOBEC
הוזכרו גם דה־אמינזות ממשפחת APOBEC, שהן ״קרובות משפחה״ של AID; תפקידן קשור לתגובה אנטי-ויראלית: הן יכולות לבצע דה־אמינציה ב־DNA או ב־RNA ויראלי, וכך לפגוע בו כחלק מהתגובה נגד וירוסים.
גם כאן יש צד בעייתי: אם פעילות כזו מתרחשת במקום הלא נכון או בעוצמה לא מבוקרת, היא יכולה לתרום לנזק גנומי.
אוקסידציה של גואנין ו־8-oxoG
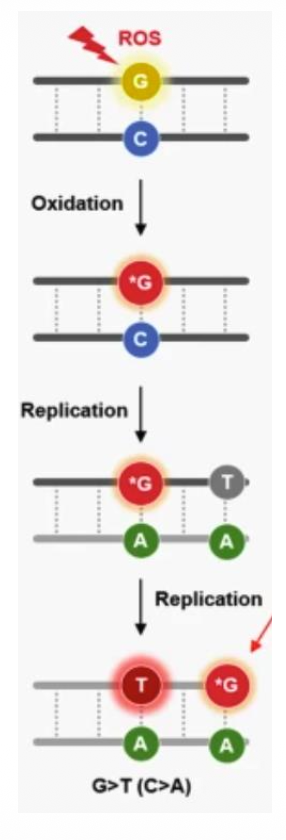
הנזק השלישי הוא אוקסידציה (Oxidation), בעיקר של גואנין.
בדיקה עצמית: מה היו שני סוגי הנזק הקודמים?
תשובה
- דה־פורינציה - איבוד בסיס פוריני (A או G)
- דה־אמינציה - הסרת קבוצת אמין (C → U)
ROS (Reactive Oxygen Species) נוצרים באופן טבעי במיטוכונדריה, בעיקר כתוצר לוואי של שרשרת מעבר האלקטרונים (נשימה תאית). הם יכולים לעלות גם בעקבות גורמים חיצוניים כמו קרינה מייננת, תרופות או הידבקויות חיידקיות וויראליות.
אחד הנזקים הנפוצים שהם יוצרים הוא: 8-oxoGuanine (8-oxoG). זהו גואנין שעבר חמצון בעמדה 8 (כ־1,500 מקרים ביום).
אם 8-oxoG לא מתוקן, הוא עלול לגרום למוטציה, משום שהוא יכול להזדווג בצורה לא תקינה עם אדנין. לכן חשוב להסיר אותו לפני שכפול DNA.
למה טלומרים פגיעים במיוחד?
כל ה־DNA חשוף לנזקי חמצון, אבל הטלומרים פגיעים במיוחד ל־8-oxoG. הסיבה היא שהטלומרים עשירים מאוד בגואנין (G). הרצף הטלומרי מורכב מחזרות של:
\[\text{TTAGGG}\]כלומר, חלק גדול מהרצף הוא G. מכיוון ש־8-oxoG נוצר מגואנין, אזורים עשירים בגואנין חשופים יותר לנזקי אוקסידציה.
תיקון 8-oxoG
כמו דה־אמינציה ודה־פורינציה, גם 8-oxoG מתוקן על ידי BER. אבל בשלב הראשון, נדרש אנזים שיודע לזהות ולהסיר 8-oxoG. האנזים הזה הוא: OGG1 (8-oxoguanine glycosylase 1).
OGG1 מקביל בתפקידו ל־UNG, אבל הוא מזהה 8-oxoG במקום אורציל. לאחר יצירת AP site, המשך התיקון דומה: APE1, פולימראז β וליגאז.
בדיקה עצית: מי היה UNG?
תשובה
UNG (Uracil DNA Glycosylase) הוא האנזים שמזהה אורציל ב־DNA ומסיר אותו, וכך יוצר אתר חסר בסיס (AP site).
בדיקה עצמית: מה זה AP site?
תשובה
AP site הוא אתר חסר בסיס, כלומר מקום ב־DNA שבו הבסיס נעלם או הוסר, אבל השלד הסוכרי-פוספטי נשאר שלם.
| נזק | אנזים זיהוי/הסרה ראשון | המשך תיקון |
|---|---|---|
| דה־אמינציה C→U | UNG | APE1 → Polymerase β → Ligase |
| אוקסידציה ל־8-oxoG | OGG1 | APE1 → Polymerase β → Ligase |
| דה־פורינציה | כבר נוצר AP site | APE1 → Polymerase β → Ligase |
מיפוי AP sites ו־8-oxoG בגנום
בחלק האחרון של השיעור דיברנו על הדרך לזהות ולמפות אתרים חסרי בסיס בגנום.
המטרה היא לא רק לדעת שיש נזק, אלא לשאול:
- האם טיפול מסוים מעלה את מספר ה־AP sites?
- איפה בגנום הם נוצרים?
- האם הם מפוזרים באופן אחיד, או מועשרים באזורים מסוימים?
הרעיון הכללי של AP-seq
השיטה היא AP-seq - מיפוי אתרים א-פוריניים / אתרים חסרי בסיס.
השלבים הכלליים:
- חותכים את ה־DNA באופן אקראי למקטעים.
- מבצעים סלקציה למקטעים שמכילים AP site.
- מרצפים את המקטעים.
- ממפים את הקריאות לגנום.
השלב החשוב הוא הסלקציה: איך מזהים דווקא מקטעים שיש בהם אתר חסר בסיס?
Isolate the Apurinic Sites
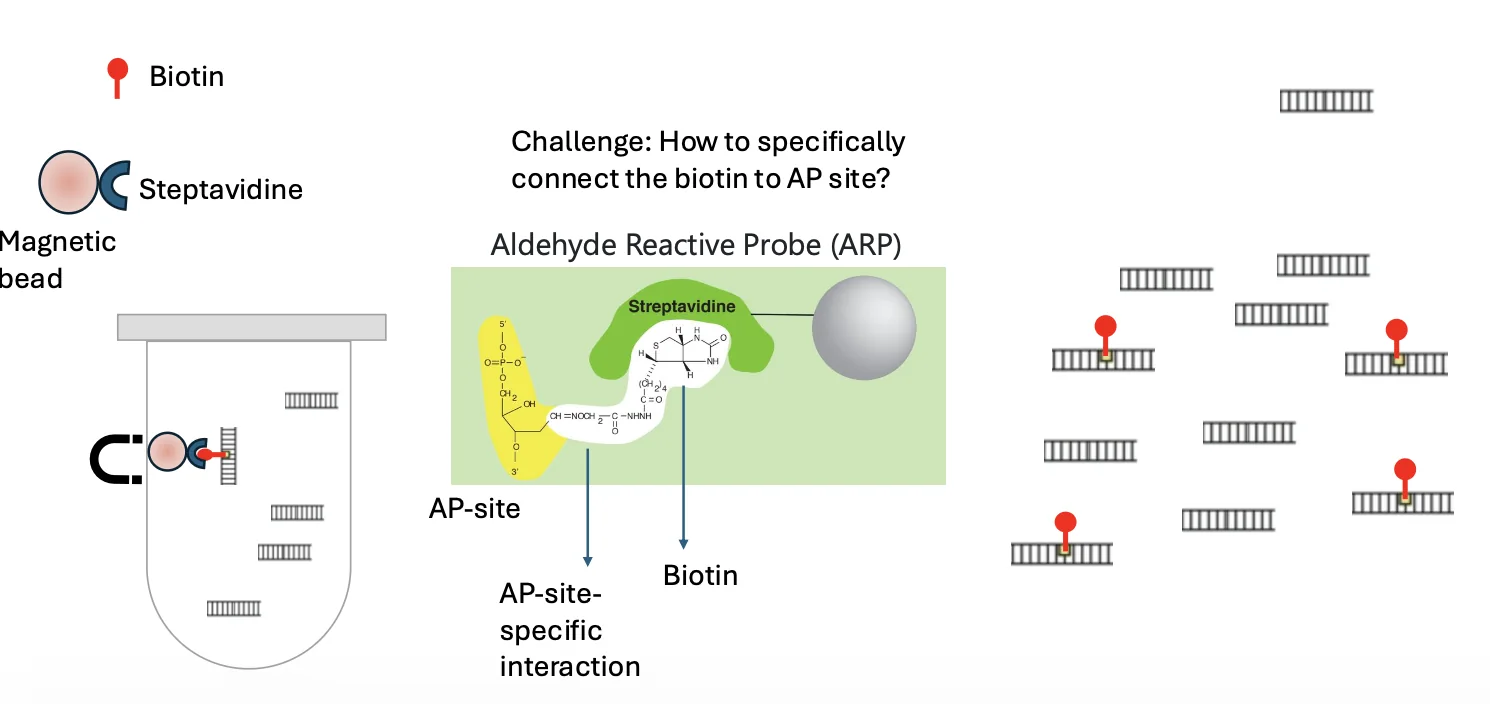
כדי לזהות AP sites משתמשים במערכת Biotin-Streptavidin.
העיקרון:
- משתמשים ב־aldehyde-reactive probe שמזהה את הסוכר באתר חסר־בסיס.
- הפרוב מחובר ל־biotin.
- כך AP sites מקבלים סימון ביוטין.
- משתמשים בבידים מגנטיים מצופים streptavidin.
- Streptavidin נקשר לביוטין בצורה מאוד ספציפית.
- בעזרת מגנט, מבודדים את מקטעי ה־DNA שמכילים AP sites.
אם יש נוגדן או חלבון שמזהה את התכונה הרצויה, אפשר להשתמש בו. אם אין, צריך לבנות מערכת זיהוי. כאן המערכת היא ביוטין-סטרפטאבידין.
שלבים 3 ו־4: Sequencing and Alignment
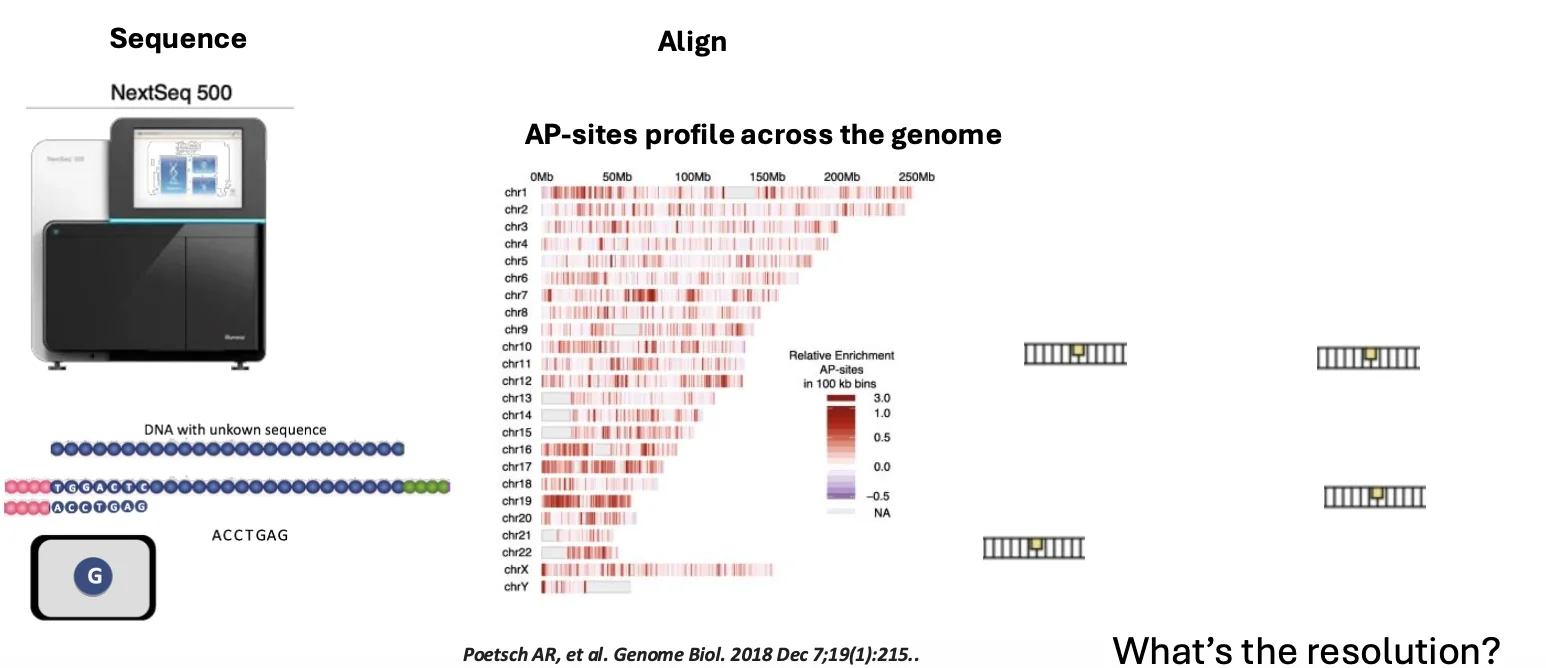
אחרי בידוד המקטעים, ממשיכים כמו בספריות DNA אחרות:
- חיבור אדפטורים
- ריצוף
- alignment לגנום
- ניתוח פיזור האתרים לאורך הגנום
התוצאה היא מיפוי AP sites. בתמונה למעלה (צד ימין) יש מפה, שבה רואים שהנזק אינו מפוזר באופן אחיד: יש אזורים שבהם יש יותר AP sites, ואזורים שבהם יש פחות.
איך ממפים 8-oxoG?
אם רוצים למפות אתרי 8-oxoG, ולא רק AP sites קיימים, אפשר להשתמש בידע ממנגנון התיקון.
הרעיון:
- לוקחים את ה־DNA.
- מטפלים בו ב־OGG1.
- OGG1 מזהה 8-oxoG ומסיר אותו.
- ההסרה יוצרת AP site.
- עכשיו אפשר להשתמש באותה שיטת AP-seq כדי למפות את האתרים.
OGG1 הופך את אתרי האוקסידציה לאתרים חסרי בסיס שניתן למפות בשיטה שתוארה.
ניסוי עם X-ray
ניסוי שבו השוו בין תאי ביקורת לתאים שטופלו ב־X-ray; מכיוון ש־X-ray משרה oxidative stress, מצפים לראות יותר נזקי חמצון.
ההשוואה הראתה:
- לאחר X-ray יש עלייה ב־AP sites לאורך הגנום.
- באתרים שהועשרו בעזרת OGG1 (אתרים שמייצגים 8-oxoG), השפעת X-ray בולטת יותר.
זה מתאים לרעיון ש־X-ray מעלה oxidative stress, ולכן מעלה בעיקר נזקי חמצון.
העשרה בטלומרים
בסוף השיעור הוצגה דוגמה לניתוח העשרה באזורים גנומיים מסוימים.
הנקודה שהודגשה: בטלומרים יש העשרה של AP sites שמקורם בנזק חמצוני, במיוחד באתרים שהועשרו בעזרת OGG1. זה מתאים למה שהסברנו לעיל: טלומרים עשירים בגואנין, ולכן הם רגישים יותר ל־8-oxoG.
סיכום
אי־יציבות גנומית היא תוצר אפשרי של נזקי DNA שלא תוקנו בזמן. היא חשובה במיוחד בסרטן, כי היא מאפשרת לתאים לרכוש תכונות סרטניות כמו הפעלת אונקוגנים או איבוד tumor suppressors. היא קשורה גם להזדקנות, בין השאר דרך ROS ונזקי DNA, וכן למחלות שבהן יש פגם במנגנוני תיקון DNA.
נזקי DNA יכולים להגיע מבחוץ, למשל מכימיקלים או מקרינה, אבל הם יכולים להיווצר גם מבפנים: שכפול DNA, פעילות topoisomerase, יצירת הרפרטואר החיסוני ו־ROS מיטוכונדריאלי.
בחלק המרכזי של השיעור התמקדנו ב־non-bulky DNA lesions (נזקים קטנים ברמת הבסיס): depurination, deamination ו־oxidation. לכל אחד מהם יש ביוכימיה אחרת, אבל שלושתם מתוקנים בעיקר על ידי Base Excision Repair.
BER עובד לפי עיקרון חוזר.
בדיקה עצמית: מה הם שלושת השלבים של BER?
תשובה
- זיהוי הנזק, ואם צריך יצירת AP site (למשל על ידי UNG או OGG1). לא צריך במקרה של דה־פורינציה, כי כבר יש AP site.
- חיתוך השלד על ידי APE1
- מילוי על ידי Polymerase β וסגירה על ידי Ligase.
אם התיקון לא מתרחש בזמן, הנזק יכול להפוך למחיקה או למוטציה לאחר שכפול DNA.
בסוף השיעור ראינו איך אפשר למפות AP sites בגנום בעזרת AP-seq, ביוטין-סטרפטאבידין ובידים מגנטיים. אותה גישה יכולה לשמש גם למיפוי 8-oxoG אם קודם משתמשים ב־OGG1 כדי להפוך את הנזק ל־AP site. כך אפשר לראות לא רק כמה נזק יש, אלא גם איפה בגנום הוא מצטבר.
בשיעור נמשיך ל־bulky lesions.
דור פסקל