כל התכנים בעמוד אחד
כל סיכומי השיעורים בווירולוגיה זמינים כאן להדפסה מרוכזת או ייצוא כ־PDF (התוכן בקובץ מעודכן ליום 18 בפברואר 2026 - ייתכן שתכני האתר יהיו עדכניים יותר).
התוכן מוצג בסדר כרונולוגי - מהסיכומים המוקדמים ביותר למאוחרים ביותר. למידה מהנה!
תוכן העניינים
- שיעור 1 וירולוגיה - מבוא לווירולוגיה 23/10/2025
- שיעור 2 וירולוגיה - מחזור החיים של וירוסים 30/10/2025
- שיעור 3 וירולוגיה - מבנה וסימטריה של וירוסים 06/11/2025
- שיעור 4 וירולוגיה - מחזור החיים של וירוסים 17/11/2025
- שיעור 5 וירולוגיה - רטרו־וירוסים, HIV, ו־HBV 20/11/2025
- שיעור 6 וירולוגיה - נגיפי RNA, רפליקציה, בקרת תרגום, הרכבה ויציאה 27/11/2025
- שיעור 7 וירולוגיה - מה וירוסים עושים לתא ומה התא עושה בחזרה 07/12/2025
- שיעור 8 וירולוגיה - עקרונות בתכנון תרופות 11/12/2025
שיעור 1 וירולוגיה - מבוא לווירולוגיה
תאריך: 23/10/2025האם וירוס הוא יצור חי?
השאלה הזו פותחת את הקורס ומלווה אותו לכל אורכו. התשובה הקצרה היא לא - יצור חי מוגדר ככזה שיש לו מטבוליזם ויכולת התרבות עצמאית, ווירוס אינו עומד בלפחות אחד מהקריטריונים הללו. בשיעור הזה נראה מדוע, ולמרות זאת - מדוע וירוסים הם בין היצירות הביולוגיות המורכבות והמרתקות ביותר בטבע.
משאבים נוספים ללימוד עצמי
This Week in Virology (TWiV) - פודקאסט ובלוג עם ראיונות שבועיים עם וירולוגים על מחקרים חדשים, כולל פודקאסט רפואי על מגיפות.
Virology Blog - חדשות עדכניות בתחום הווירולוגיה.
אתר פרופ’ ג’וני דירשוני - מאגר וירולוגים בישראל; ניתן לחפש לפי אוניברסיטה או לפי וירוס.
היסטוריה של חקר הנגיפים
עדויות עתיקות ומחלות היסטוריות
Smallpox - אבעבועות שחורות
עדויות ל־Smallpox קיימות כבר מלפני 3,500 שנה. מדובר במחלה שבה הווירוס מתרבה בגוף ללא סימנים חיצוניים בימים הראשונים, ואז מופיעות שלפוחיות על העור שבסופו של דבר מתייבשות ונופלות - ומשאירות צלקות, ולעיתים אף גורמות לעיוורון. הנתון המזעזע: 30% מהנדבקים מתו, ובמאה ה־20 לבדה כ־300 מיליון איש נפטרו מהמחלה.
🟢 Variolation - החיסון המוקדם (סין, המאה ה־11). השיטה הייתה פשוטה ומסוכנת בו־זמנית: לקחו חומר משלפוחיות של חולה, שרטו את העור של אדם בריא, והקווה הייתה שההדבקה המקומית דרך העור לא תגרום למחלה מערכתית מלאה. התוצאות: 90% קיבלו חסינות, אך 10% חלו - ומתוכם 30% מתו. כלומר התמותה הכוללת ירדה מ־30% ל־3%. לא אידיאלי, אבל הרבה יותר טוב ממה שהיה.
לידי מרי וורטלי מונטגו, אשת השגריר הבריטי במזרח, נחשפה לשיטה הזו והביאה אותה לאירופה.
🟢 Edward Jenner והמצאת החיסון (1796). ג’נר שם לב שכפריים שעובדים עם פרות לא חלו ב־Smallpox. לפרות הייתה מחלה דומה עם שלפוחיות על העור (Cowpox), והרעיון היה שהכפריים נדבקו מהפרות וקיבלו חיסון טבעי. ג’נר לקח את הווירוס מהפרות, שרט את העור של אנשים, והכניס להם את הווירוס - וזה עבד. זה היה החיסון הראשון בהיסטוריה. Smallpox נשאר הווירוס היחיד עד היום שהוצא מהעולם לחלוטין: ב־1980 ארגון הבריאות העולמי הכריז על חיסולו. לצערנו, עם וירוסים אחרים לא הצלחנו להשיג תוצאה דומה.
Rabies - כלבת
מחלה ידועה כבר 4,000 שנה מכתבים עתיקים, וחשובה לרפואה עד היום. הווירוס מגיע דרך נשיכה, נכנס לגוף, ומתחיל להתקדם לכיוון המוח - מבלי להתרבות בדרך. ברגע שהגיע למוח מופיעות תופעות קליניות, אבל מאותו רגע אין דרך חזרה - גם היום, ב־2025, הגעת הווירוס למוח היא פסק דין.
המטרה היא מניעה. לואי פסטור פיתח חיסון פסיבי: הזריק את הווירוס לארנבות, אסף מהדם שלהן נוגדנים, ומי שנישך קיבל הזרקת נוגדנים. היעילות גבוהה מאוד - בתנאי שהטיפול ניתן לפני שהווירוס הגיע למוח.
פריצות דרך מדעיות
הפילטר של Chamberland (1884) - פילטר עם נקבים כל־כך קטנים שחיידקים לא עוברים. זה אפשר לראשונה לסנן נוזלים ולקבל תמיסה נקייה מחיידקים.
Tobacco Mosaic Virus - TMV (1892) - אדולף מאייר ודמיטרי איבנובסקי הבחינו בכתמים על עלי טבק. הם טחנו עלים נגועים, העבירו את התמיסה דרך פילטר שחיידקים לא עוברים, והדביקו צמח בריא - והכתמים עברו. המסקנה: הגורם למחלה קטן מחיידק. קראו לו “וירוס”, שפירושו “רעל” בלטינית.
Polio וריאת הברזל - בשנות ה־50 מגיפות פוליו פקדו שכונות בקיץ. ילדים שנדבקו איבדו פתאום את יכולת ההליכה. ריאת הברזל הייתה מכשיר שהפעיל את בית החזה מבחוץ, כי שרירי הנשימה לא תפקדו. הפריצה הגיעה ממיפוי: כל השכונות הנגועות קיבלו מים מאותו מקור - והמים היו מזוהמים בווירוס.
המיקרוסקופ האלקטרוני - וירוסים קטנים מאורך הגל של אור נראה, ולכן אי־אפשר לראותם במיקרוסקופ אור רגיל. רק עם המצאת המיקרוסקופ האלקטרוני ניתן היה לראות וירוסים לראשונה.
מבנה הנגיף
זיהוי החומר הגנטי
ניסוי מכונן בווירולוגיה: פירקו את TMV לשני מרכיביו - חלבונים וחומצת גרעין. כשהדביקו עלים רק בחלבונים, לא הייתה הדבקה; כשהדביקו רק בחומצת גרעין - כן הייתה הדבקה. השלב הבא: טיפול באנזים RNase (שמפרק RNA) ביטל את ההדבקה, בעוד DNase (שמפרק DNA) לא השפיע. המסקנה: החומר הגנטי של TMV הוא RNA - שלא כמו בכל היצורים החיים שבהם החומר הגנטי הוא DNA.
ניסוי ה־Reconstitution חיזק את הממצא: לקחו שני זני TMV עם פאטרן כתמים שונה (A ו־B), פירקו כל אחד, והרכיבו וירוס היברידי מחלבון A ו־RNA של B. כשהדביקו עלים - הפאטרן היה של B. כלומר ה־RNA הוא שקובע את התכונות.
רכיבי הנגיף
Capsid (קפסיד) - קופסה חלבונית סימטרית שעוטפת את הגנום. מורכבת מיחידות חוזרות של חלבון אחד או מספר חלבונים. האינטראקציות בין היחידות הן לא־קוולנטיות - כדי שהווירוס יוכל גם להיסגר ולהגן על הגנום, וגם להיפתח כשמגיע לתא חדש. כשהקפסיד מכיל בתוכו חומצת גרעין, הוא נקרא Nucleocapsid.
Envelope (מעטפת) - שכבת שומנים שמקיפה את הקפסיד. הנקודה המרכזית: הווירוס לא מייצר מעטפת - הוא “גונב” אותה מממברנת התא המודבק. אבל בתוך אותה מעטפת שזורים חלבונים ויראליים (כמו חלבון ה־Spike של הקורונה), שהם אלו שמערכת החיסון “רואה”. לא לכל הווירוסים יש מעטפת - יש כאלה שמסתדרים בלעדיה.
שכבת ביניים - בחלק מהווירוסים יש שכבה בין הקפסיד למעטפת: אם היא מסודרת היא נקראת Matrix, ואם היא אמורפית - Tegument.
דוגמאות למבנים
הווירוסים מגוונים מאוד במבנה: Bacteriophage הוא וירוס מורכב עם ראש, זנב ורגליים שמדביק רק חיידקים; TMV הוא בצורת מקל מוארך; HIV הוא כדורי עם קפסיד בצורת קונוס עטוף במעטפת; ו־Herpesvirus הוא וירוס גדול עם מעטפת, טגומנט וקפסיד.
גודל
רוב הווירוסים נעים בין 20 ל־300 ננומטר - קטנים בהרבה מחיידקים (שהם בסדר גודל של מיקרון). אבל קיימים Giant Viruses שמדביקים אמבות: ה־Mimivirus, למשל, גדול כמו חיידק, גנום שלו (2.5 מיליון זוגות בסיסים) גדול יותר מגנום של חיידקים רבים, והוא מקודד לכ־2,500 חלבונים. אפשר אפילו לראות אותו במיקרוסקופ אור רגיל.
יחס זיהומיות
לא כל הווירונים שנוצרים הם בעלי יכולת הדבקה. בשפעת, למשל, רק כ־10% מהווירונים אינפקטיביים; בהרפס - רק אחד מכל אלף. הווירוס מפצה על כך בייצור כמויות עצומות - מעין “הפצצה” בווירונים שמבטיחה שמספיק מהם יצליחו.
מחזור חיים של וירוס - דוגמה: SV40
הדוגמה של SV40 ממחישה עיקרון מרכזי: הווירוס לא מביא כלום חדש - הוא מגייס מערכות קיימות.
התהליך:
- DNA של וירוס נכנס לגרעין התא
- התא אורז אותו בנוקלאוזומים (בדיוק כמו DNA תאי)
- RNA polymerase תאי מבצע טרנסקריפציה
- ה־RNA יוצא לציטופלזמה ומתורגם לחלבון ויראלי
- אותו חלבון חוזר לגרעין ← משנה את מערכת הטרנסקריפציה כך שתייצר חלבונים ויראליים נוספים ותשכפל את הגנום
- הקפסידים נבנים, הגנום נארז, ויריונים חדשים משתחררים.
הנקודה: הווירוס מגייס את מערכת הטרנסקריפציה ואת מערכת הרפליקציה של התא - גם לביטוי הגנים שלו וגם להכפלת הגנום.
תכונות משותפות לכל הווירוסים
-
טפילות מוחלטת (Obligate parasites) - הווירוס תלוי לחלוטין בתא כדי להתרבות. ללא תא מאכסן, הוא חלקיק אינרטי.
-
גודל קטן ומידע גנטי מצומצם - לבן אדם יש 21,000 גנים ו־3 מיליארד בסיסים; לווירוס רגיל יש 2–200 גנים ו־2,000–300,000 בסיסים. למרות הגודל הזעיר, מספיק לו 2,000 בסיסים כדי לייצר וירוס מתרבה.
-
גנום RNA או DNA - לא שניהם - שלא כמו כל היצורים החיים שהחומר הגנטי שלהם הוא תמיד DNA.
-
מבנה פשוט אך ביולוגיה מורכבת - למרות שהמבנה הפיזי פשוט יחסית (מעטפת, קפסיד, חומצת גרעין), הווירוסים מפגינים מורכבות עצומה ביכולתם לשחק עם מערכות תאיות ולגייס אותן לצרכיהם. כל הזמן מגלים שכבות חדשות של מורכבות.
שלושה דברים שווירוסים לא מייצרים
| לא מייצרים | הסבר |
|---|---|
| אנרגיה (ATP) | תלויים לחלוטין בהפקת אנרגיה של התא |
| ריבוזומים | משתמשים בריבוזומים התאיים לתרגום. אפילו Giant Viruses שמייצרים tRNA ספציפיים - לא מייצרים ריבוזומים שלמים |
| ממברנה | חלבונים ויראליים יכולים להשתלב בממברנה, אבל התא מייצר את הממברנה עצמה |
סיווג בולטימור (Baltimore Classification)
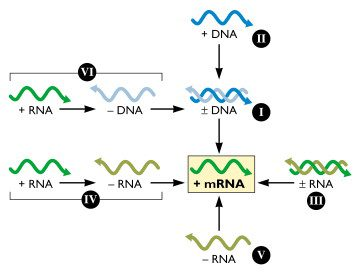
התמונה מתוך virology.ws
הסיווג מבוסס על עיקרון אחד: כל וירוס חייב לייצר mRNA כדי שהריבוזומים יתרגמו אותו לחלבונים. השאלה: איך כל סוג של גנום מגיע ל־mRNA?
תזכורת:
RNA+(פלוס) = mRNA = קריא ישירות על־ידי ריבוזומים.RNA-(מינוס) = הגדיל המשלים, לא קריא ישירות.
Class I - dsDNA → mRNA
מערכת הטרנסקריפציה התאית יודעת לעבוד עם dsDNA, אז אין בעיה.
דוגמאות: Herpesvirus, Adenovirus.
Class II - ssDNA → dsDNA → mRNA
קודם התא משלים את הגדיל החסר ליצירת dsDNA, ואז מתבצעת טרנסקריפציה רגילה.
דוגמאות: Parvovirus.
Class III - dsRNA → (+)RNA
הריבוזום לא יודע לתרגם dsRNA - צריך ליצור (+)RNA מתבנית ה־(-)RNA.
דוגמאות: Reovirus.
Class IV - (+)ssRNA
ה־RNA+ יכול לשמש ישירות כ־mRNA, אבל הכמות לא מספיקה. לכן הווירוס מייצר (-)RNA כתבנית, וממנו מייצר כמויות גדולות של (+)RNA.
דוגמאות: Poliovirus, Coronavirus.
Class V - (-)ssRNA → (+)RNA
ה־RNA- אינו קריא; צריך ליצור ממנו (+)RNA שישמש כ־mRNA.
דוגמאות: Influenza, Measles.
Class VI - Retroviruses: (+)RNA → DNA → mRNA
שלושה שלבים:
- RNA הופך ל־ssDNA (Reverse transcription),
- ssDNA הופך ל־dsDNA,
- ומ־dsDNA מתבצעת טרנסקריפציה רגילה ליצירת mRNA.
ה־mRNA שנוצר הוא (+)RNA - כי מערכת הטרנסקריפציה התאית היא שמייצרת אותו.
דוגמאות: HIV.
Class VII - Partially dsDNA → dsDNA → mRNA
הגנום הוא dsDNA חלקי. מערכת התיקון התאית משלימה אותו ל־dsDNA מלא, ואז אפשר לבצע טרנסקריפציה.
דוגמאות: Hepatitis B.
DNA לעומת RNA - יציבות והשלכות
DNA יציב יותר מ־RNA, ו־dsDNA יציב יותר מ־ssDNA. לכן, גנומים של RNA מוגבלים בגודלם (עד כ־40,000 בסיסים), בעוד וירוסי dsDNA יכולים להיות גדולים בהרבה (כמו Herpesvirus ו־Poxvirus).
בחיידקים רוב הווירוסים הם בעלי גנום DNA; בצמחים הרבה מאוד וירוסים הם בעלי גנום RNA.
אבולוציה ויראלית
וירוסים כמכונות דרוויניסטיות
אורגניזמים צריכים דורות רבים כדי לעבור שינויים משמעותיים, אבל מחזור החיים של וירוס נמשך ימים בודדים - ולכן קצב האבולוציה שלו מהיר להפליא.
יתרה מכך, בווירוסי RNA אין מנגנון תיקון (proofreading). הפולימראז מכניס מוטציות בכוונה - וזה יתרון, לא חיסרון. כל מחזור שכפול מייצר גיוון גנטי שמאפשר לווירוס לחמוק ממערכת החיסון.
דוגמה קלאסית - שפעת: אנחנו מייצרים נוגדנים נגד חלבוני מעטפת השפעת, אבל הם משתנים כל הזמן. לכן אנחנו נדבקים שוב ושוב, ולכן צריך חיסון שפעת חדש כל שנה.
מעבר למאכסן חדש - דוגמת COVID-19
כשווירוס עובר למאכסן חדש (Zoonosis), קורים שני דברים מקבילים:
בהתחלה - הווירוס לא מותאם למאכסן החדש. הוא לא עובר ביעילות בין בני אדם, אבל הוא גורם למחלה קשה מאוד. במקרה של SARS-CoV-2, הווירוס גרם לחיבור (fusion) של תאי ריאה אחד לשני, יצירת בצקת, וחנק.
עם הזמן - הלחץ האבולוציוני פועל: וירוס שהורג את המאכסן מהר לא מספיק להתפשט; וירוס שגורם למחלה קלה מאפשר למאכסן להמשיך לתפקד ולהדביק אחרים. לכן האבולוציה מובילה לווירוס שעובר טוב יותר בין אנשים אבל גורם למחלה קלה יותר.
COVID-19 של היום כמעט לא דומה לזה של ההתחלה - הוא עבר מוטציה בחלבון ממברנה שביטלה את יכולת ה־Fusion.
💡 העיקרון: המטרה של הווירוס היא לעבור ממאכסן למאכסן - לא להרוג. מות המאכסן הוא תוצר לוואי, לא מטרה.
למה בכל זאת קיימים וירוסים קטלניים?
אם האבולוציה מובילה לווירוס “עדין” יותר, למה יש וירוסים שהורגים?
- מעבר טרי בין מאכסנים - בשלבים הראשונים, לפני שהתרחשה התאמה.
- דרך הדבקה שלא תלויה בחיי המאכסן - כלבת עוברת דרך נשיכה: אפילו אם החולה מת, הווירוס כבר הדביק. באותו אופן, וירוסים שעוברים דרך יתושים (Vector) לא תלויים בחיי החולה.
- אנשים מדוכאי חיסון - אצלם הווירוס שוהה חודשים, מתרבה כל הזמן, וצובר מוטציות. כך נוצרים ווריאנטים חדשים - וזה בדיוק מה שקרה עם הווריאנטים החדשים של COVID-19.
חיטוי והשמדת וירוסים
כדי להבין חיטוי, צריך לזכור את שלושת המרכיבים של הווירוס: מעטפת שומנית, קפסיד חלבוני, וחומצת גרעין.
-
סבון פועל על שומנים - ולכן יעיל נגד וירוסים עם מעטפת. הוא פשוט מפרק את המעטפת השומנית.
-
אלכוהול (70%) משפיע גם על המעטפת וגם על מבנה החלבונים (דנטורציה), ולכן יעיל גם נגד וירוסים ללא מעטפת - כי הוא פוגע בקפסיד.
-
אקונומיקה (0.1%) ו־מי חמצן - שניהם מחמצנים חזקים שפוגעים גם בחלבונים וגם בחומצת גרעין. אבל הם דורשים לפחות 10 דקות של מגע עם המשטח כדי להשפיע. חשוב: לשימוש על משטחים בלבד - לא לשתייה.
Virion לעומת Virus - ההבדל המהותי
ההבחנה הזו חשובה מאוד:
- Virion הוא חלקיק הווירוס מחוץ לתא - אינרטי לחלוטין, לא עושה דבר, “מולקולה ביולוגית” סבילה.
- Virus הוא מה שקורה בתוך תא מודבק - המערכת הפעילה שמשתלטת על התא ומייצרת עותקים חדשים.
סיווג וירוסים - קריטריונים
-
לפי מאכסן: וירוסי חיידקים (Bacteriophages), וירוסים של צמחים, בעלי חיים, פטריות ואורגניזמים חד־תאיים.
העיקרון: אין יצור חי על פני כדור הארץ שאין לו וירוס שיכול להדביק אותו. ובמקביל, וירוסים הם ספציפיים מאוד - פאג’ שמדביק חיידקים לא ידביק בן אדם.
קריטריונים נוספים: סוג הגנום (DNA/RNA, חד/דו־גדילי), סימטריה של הקפסיד (Icosahedral או Helical), נוכחות מעטפת, גודל, טווח מאכסנים, ופתוגנזה.
מגיפות עכשוויות - דוגמת Oropouche Virus
מאמר ב־Science (2024) תיאר התפרצות של וירוס Oropouche בברזיל. שני וירוסים שונים, שלא גרמו למחלה בבני אדם, הדביקו את אותו מאכסן. הגנום שלהם מורכב משלושה סגמנטים, והסגמנטים התערבבו - ונוצר וירוס חדש שגורם למחלה בבני אדם.
התסמינים (חום, כאבי ראש, כאבי פרקים) דומים למחלות רבות - כי מדובר בתגובה כללית של הגוף. במקרים חמורים: דלקת מוח, נזק נוירולוגי, מוות. נשים בהריון שנדבקו ילדו תינוקות עם מיקרוצפליה (מוח קטן מהרגיל). הריסוס שנעשה בשטח מכוון נגד היתוש (ה־Vector) ולא נגד הווירוס עצמו.
וירוסים לא תמיד רעים
פאג’ תרפיה - וירוסים נגד חיידקים
אנטיביוטיקה הורגת חיידקים באופן רחב, כולל את הפלורה המועילה. Bacteriophages, לעומת זאת, ספציפיים - אפשר לתכנן פאג’ שיהרוג רק סוג חיידק מסוים.
וירוסים באוקיאנוסים - מנוע אקולוגי
המספרים מרשימים: כפית מי ים מכילה כ־5 מיליון חיידקים, וליטר מי ים מכיל יותר וירוסים מכל בני האדם על פני כדור הארץ. 94% מכל החלקיקים באוקיאנוסים הם וירוסים - כ־$10^{30}$ בסך הכול. כל יום כ־20% מחיידקי האוקיאנוס מתים על־ידי וירוסים, מתפרקים לחומרי הזנה, ומאפשרים גדילה מחדש - מחזור חיים קריטי לאקו־מערכת הימית.
הגנה צולבת - ניסוי בעכברים
עכברים שהודבקו ב־Herpesvirus שרדו בשיעור גבוה בהרבה כשנחשפו בהמשך לחיידקים קטלניים או לשפעת H1N1 - לעומת עכברים שלא נשאו Herpesvirus. ההסבר: הווירוס מעלה את מוכנות המערכת החיסונית באופן כללי (לא ספציפי). אולי ההדבקה הכרונית בהרפס שרוב האנשים נושאים נותנת לנו רמה מסוימת של הגנה.
סימביוזה - צרעה, זחל, ווירוס
דוגמה יפה לקשר בין וירוס ליצור חי: צרעות מטילות ביצים בתוך זחלים, אבל מערכת החיסון של הזחל תוקפת את הביצים. הפתרון: הצרעה מזריקה ביחד עם הביצים וירוס שמשתק את מערכת החיסון של הזחל. הווירוס הזה הוא חלק מהגנום של הצרעה - לא מתבטא בכלל בגוף הצרעה הבוגרת, אלא רק בזמן יצירת ביציות.
שאלות לבדיקה עצמית
- מדוע וירוס אינו נחשב ליצור חי?
- תאר את מחזור החיים של SV40 - אילו מערכות תאיות הוא מגייס?
- מה ההבדל בין RNA+ ל־RNA-? מדוע ההבחנה חשובה?
- מדוע אבולוציה ויראלית מובילה בדרך כלל למחלה קלה יותר? מתי זה לא נכון?
- תן שלוש דוגמאות לווירוסים שיכולים להיות מועילים.
- מדוע סבון יעיל נגד קורונה אבל פחות נגד Adenovirus?
- הסבר את סיווג בולטימור - מה השאלה המרכזית שהוא עונה עליה?
שיעור 2 וירולוגיה - מחזור החיים של וירוסים
תאריך: 30/10/2025מבוא
בשיעור זה נעסוק במחזור החיים של וירוסים, החל מהתהליך שבו הווירוס מדביק תא ועד לשחרור וירוסים חדשים. נדון במושגים חשובים כגון תאים רגישים ומתירניים, דרכי העברה שונות של וירוסים, מדדי הדבקה ($R$), ושיטות זיהוי מעבדתיות מתקדמות.
שלבי מחזור החיים של וירוסים
1. Attachment (היצמדות)
הווירוס צריך להגיע לתא המארח ולהיצמד לרצפטור ספציפי על פני התא. תהליך ההיצמדות הוא ספציפי ביותר – כל וירוס בחר בתהליך האבולוציה רצפטור מסוים.
2. Penetration (חדירה)
לאחר ההיצמדות, הווירוס חודר לתוך התא. תהליך זה יכול להתבצע בדרכים שונות:
- Fusion – התמזגות עם ממברנת התא
- Endocytosis – בליעה לתוך וזיקולה תאית
3. Uncoating (חשיפת החומר הגנטי)
הקפסיד (Capsid) נפתח ומשחרר את החומר הגנטי ($\ce{DNA}$ או $\ce{RNA}$) לתוך התא. תהליך זה מתרחש לרוב בעקבות שינוי ב־pH או פעולת אנזימים תאיים.
4. Expression (ביטוי גנים)
החומר הגנטי הוויראלי מתורגם לחלבונים על ידי מנגנוני התא המארח.
5. Replication (הכפלת הגנום)
הגנום הוויראלי מוכפל באמצעות אנזימים וויראליים או תאיים. תהליך זה תלוי בסוג החומר הגנטי של הווירוס.
6. Assembly (הרכבה)
חלבונים וויראליים וגנומים חדשים מתאספים ליצירת וירוסים שלמים (virions).
7. Maturation (הבשלה)
הווירוסים עוברים שינויים סופיים הנחוצים לתפקודם.
8. Release (שחרור)
הווירוסים החדשים משתחררים מהתא באחת משתי דרכים:
- Lysis – פיצוץ התא
- Budding – שחרור הדרגתי דרך הממברנה
תאים רגישים ומתירניים
הגדרות
- תא רגיש (Susceptible) – תא שיש לו רצפטור שהווירוס יכול להיצמד אליו ולחדור דרכו.
- תא מתירני (Permissive) – תא שמאפשר את התרבות הווירוס בתוכו, כלומר יש בו את כל המנגנונים הנדרשים לשכפול הוויראלי.
דוגמאות
- HIV – מדביק תאי T (CD4+), אך אם מוכנס באופן מלאכותי לתאי עכבר, לא יתרבה שם (לא מתירני).
- Hepatitis B – מדביק רק תאי כבד (הפטוציטים), שהם גם רגישים וגם מתירניים לווירוס זה.
חשיבות: כדי שוירוס יתרבה בתא, התא חייב להיות גם רגיש וגם מתירני.
דרכי העברה של וירוסים
העברה אופקית (Horizontal Transmission)
מעבר של וירוס ממארח למארח באותה הדור:
- אוויר – טיפות רוק (Droplets), אירוסולים
- מגע ישיר – דם, רוק, יחסי מין
- וקטורים – יתושים, קרציות
העברה אנכית (Vertical Transmission)
מעבר מאם לעובר או לתינוק:
- דרך השליה (Transplacental)
- במהלך הלידה
- הנקה
זואונוזות (Zoonotic Diseases)
מחלות שמקורן בבעלי חיים:
- West Nile Virus – מציפורים דרך יתושים
- Rabies – מבעלי חיים נגועים
- Avian Flu (H5N1) – מעופות
- Ebola – מקופים
- SARS-CoV-2 – כנראה מעטלפים
וירמיה (Viremia)
הגדרה: מצב שבו וירוסים נמצאים בזרם הדם.
משמעות: וירמיה מאפשרת לווירוס להתפשט מאיבר ההדבקה הראשוני לאיברים נוספים בגוף, מה שעלול להוביל לסיבוכים מערכתיים.
Entry site is not necessarily the final target!
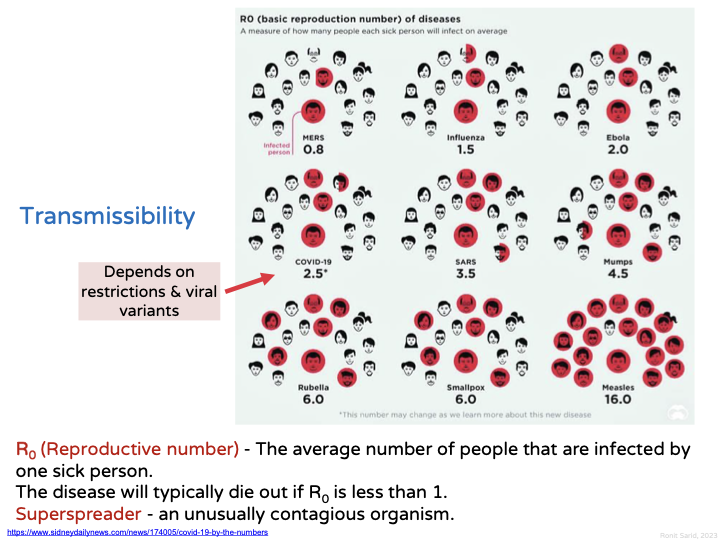
מדדי הדבקה – Reproductive Number ($R$)
הגדרה: מספר האנשים הממוצע שאדם חולה אחד מדביק.
דוגמאות
- $R < 1$ – המחלה אינה מתפשטת (דוגמה: MERS, $R \approx 0.7$)
- $R = 1$ – המחלה יציבה
- $R > 1$ – המחלה מתפשטת באוכלוסייה
- Ebola: $R \approx 2$
- SARS-CoV-2: $R \approx 2-3$ (משתנה לפי וריאנט)
- Measles (חצבת): $R \approx 16$ – אחת המחלות המדבקות ביותר!
חשיבות: ערך $R$ גבוה מחייב אחוזי חיסון גבוהים יותר להשגת חסינות עדר.
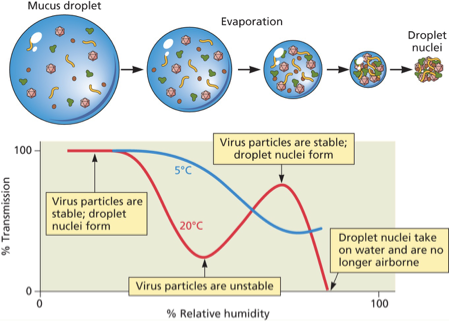
יציבות וירוסים מחוץ לגוף
גורמים המשפיעים על יציבות
- חום – טמפרטורות גבוהות מפרקות חלבונים וויראליים
- לחות – משפיעה על יציבות הקפסיד והממברנה
- קרינת UV – פוגעת בחומר הגנטי
- מעטפת – וירוסים ממוטפים (Enveloped) רגישים יותר לתנאי סביבה
דוגמאות
- HIV – בר-קיימא מספר שעות בלבד מחוץ לגוף
- Hepatitis B – יכול לשרוד שבועות על משטחים
- Norovirus – עמיד מאוד לתנאי סביבה
עקומת יציבות וטמפרטורה־לחות
- לחות נמוכה (<30%) – וירוסים יציבים בכל טמפרטורה
- לחות בינונית (50%) – יציבות גבוהה בטמפרטורות נמוכות (~5°C), יציבות נמוכה בחום (>20°C)
- לחות גבוהה (>80%) – וירוסים מתפרקים במהירות
משמעות קלינית: מרק חם בחורף יכול לעזור – החום והלחות מפחיתים את יציבות הווירוסים בדרכי הנשימה.
התפרצויות עונתיות (Seasonal Outbreaks)
גורמים
- תנאי אקלים – טמפרטורה ולחות מעדיפות
- התנהגות אנושית – שהייה במקומות סגורים בחורף
- מחזורי החיים של וקטורים – פעילות יתושים בקיץ
דוגמאות
- Influenza – פיק בחורף במדינות ממוזגות
- Poliovirus – דווקא בקיץ (תנאים חמים)
- West Nile Virus – קיץ (תלוי בפעילות יתושים)
שיטות זיהוי מעבדתיות
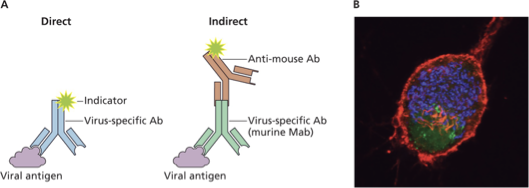
1. Immunofluorescence (אימונופלואורסצנציה)
Direct Method
- נוגדן ספציפי לאנטיגן וויראלי, מסומן בצבע פלואורסצנטי
- קושר ישירות לווירוס בדגימה
Indirect Method
- נוגדן ראשוני (לא מסומן) כנגד הווירוס
- נוגדן משני מסומן (למשל Anti-Mouse Antibody) כנגד הנוגדן הראשוני
- יתרון: חסכוני – נוגדן משני אחד לכל נוגדני העכבר
2. ELISA (Enzyme-Linked Immunosorbent Assay)
עיקרון
- נוגדן קשור למשטח
- הוספת דגימה (עם אנטיגן וויראלי)
- הוספת נוגדן שני מסומן באנזים
- הוספת סובסטרט שמייצר צבע
- מדידת עוצמת הצבע = כמות הווירוס
שימושים
- כימות וירוסים בדגימות
- בדיקות סרולוגיות (נוגדנים בדם)
3. Rapid Antigen Tests (בדיקות אנטיגן מהירות)
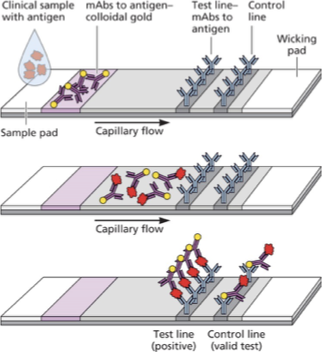
מבנה
- Control Line – נוגדן כנגד הנוגדן המסומן (תמיד חיובי אם הבדיקה תקינה)
- Test Line – נוגדן כנגד אנטיגן וויראלי ספציפי
תוצאות
- קו אחד (Control) – שלילי
- שני קווים (Control + Test) – חיובי
- אין קווים – הבדיקה פגומה
4. PCR (Polymerase Chain Reaction)
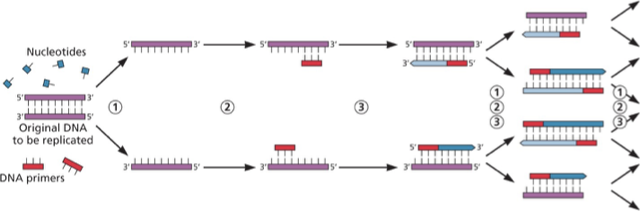
עיקרון בסיסי
PCR מאפשר הכפלה מעריכית של רצף $\ce{DNA}$ ספציפי.
שלבים
- Denaturation (95°C) – הפרדת גדילי $\ce{DNA}$
- Annealing (50-60°C) – היצמדות Primers
- Extension (72°C) – הארכת הגדיל על ידי DNA Polymerase
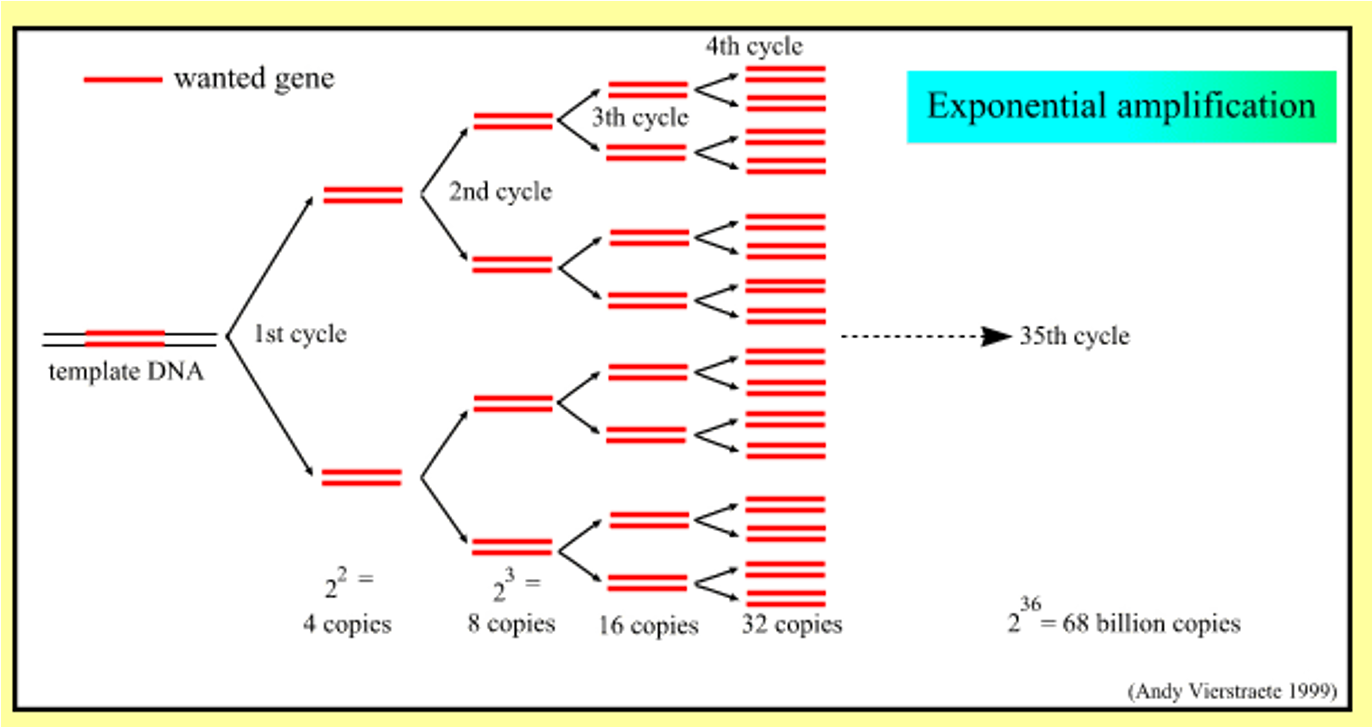
מחזור ההכפלה
- מחזור 1: 2 מולקולות
- מחזור 2: 4 מולקולות
- מחזור n: $2^n$ מולקולות
- 36 מחזורים: כ־68 מיליארד מולקולות
Taq Polymerase
- אנזים מחיידק Thermus aquaticus מעיינות רותחים
- עמיד לטמפרטורות גבוהות (עד 95°C)
- מאפשר ביצוע מחזורים חוזרים ללא הוספת אנזים חדש
5. RT-PCR (Reverse Transcription PCR)
שימוש
זיהוי וירוסי $\ce{RNA}$ (כמו SARS-CoV-2, Influenza)
שלבים
- Reverse Transcription – המרת $\ce{RNA}$ ל־$\ce{cDNA}$ באמצעות Reverse Transcriptase
- PCR רגיל על ה־$\ce{cDNA}$
דוגמה: אבחון COVID-19
- שני זוגות Primers לשני אזורים שונים בגנום
- שני פסים – חיובי
- פס אחד – אפשרי מוטציה באחד האזורים
- אפס פסים – שלילי
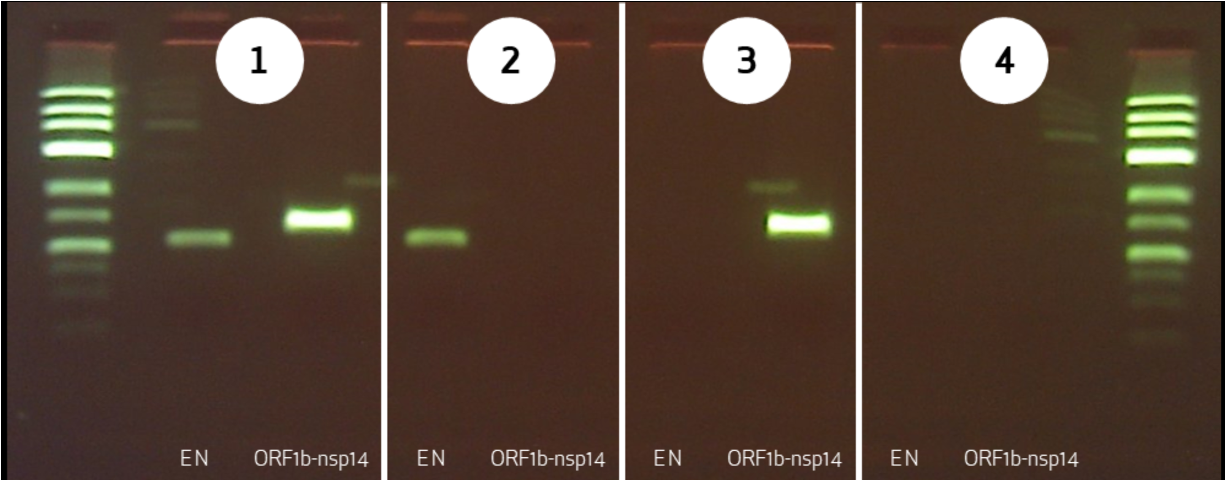
חשיבות: Primers הם רצפים ספציפיים לווירוס מסוים. PCR לגילוי קורונה לא יזהה וירוס שפעת!
שיטות תרבית וירוסים
1. תרבית תאים (Cell Culture)
- תאים בתרבית מדוגמה או שרה אחת של תאים
- מאפשרת מחקר ושכפול וירוסים במעבדה
2. ביצים מעוברות (Embryonated Eggs)
- הזרקת וירוס לאזורים שונים בביצה
- שימוש נפוץ בייצור חיסוני שפעת
- בעיה נוכחית (2023): מחסור בביצים בארה”ב בשל Avian Flu – הובילה לעלייה דרמטית במחיר (מדולר אחד, לשבע ולתריסר)
3. מודלים של בעלי חיים
- עכברים, אוגרים, פרימטים
- שימוש במחקר פתוגנזה וחיסונים
- דוגמה: עכברים מהונדסים עם רצפטור ACE2 אנושי למחקר COVID-19
מדידת כמות וירוסים – Plaque Assay
עיקרון
- שכבה של תאים (Monolayer)
- כיסוי בשכבת אגר חצי־מוצקה
- הדבקה בווירוס
- וירוס הורג תאים מקומיים ויוצר “חור” (Plaque)
חישוב
\[\text{Viral titer (PFU/ml)} = \frac{\text{Number of plaques} \times \text{Dilution factor}}{\text{Volume plated (ml)}}\]דוגמה
- דילול $10^{-7}$
- 10 Plaques
- נפח $0.1 \, \mathrm{ml}$
יתרון
מודד רק וירוסים אינפקטיביים – שונה מ־PCR (מודד גם $\ce{DNA}$ של וירוסים מתים) ו־ELISA (מודד גם קפסידים ריקים).
ייצור חיסונים מבוסס ביצים
תהליך
- בידוד זן שפעת מקומי
- שילוב (Reassortment) עם זן מעבדה שגדל היטב בביצים
- גנום שפעת מורכב מ־8 סגמנטים של $\ce{RNA}$
- הדבקת תא בשני זנים גורמת לערבוב סגמנטים
- הדבקת ביצים רבות בזן ההיברידי
- איסוף וירוסים
- Inactivation (השבתה)
- ניקוי וביקורת איכות
- אריזה להזרקה

Reassortment בשפעת
- 8 סגמנטים של $\ce{RNA}$ בגנום
- מאפשר שילוב מהיר של חומר גנטי בין זנים
- סיבה מרכזית לשונות הגבוהה והצורך בחיסון שנתי
תופעות מיוחדות בדינמיקה של וירוסים
Eclipse Phase
- תקופה שבה אין וירוסים זמינים מחוץ לתא
- הווירוס נמצא בתהליך שכפול בתוך התא
- משך: שעות עד ימים (תלוי בווירוס)
Burst vs. Budding Release
- Burst (Multiple Cycle): נפיחת תא משחררת וירוסים חדשים פתאום ← פיק שני אם לא כל התאים נדבקו בפעם הראשונה
- One-Step Growth: כל התאים נדבקו מלכתחילה ← פיק אחד
נקודות מרכזיות לזכור
- מחזור חיים וויראלי מורכב מ־8 שלבים: Attachment → Penetration → Uncoating → Expression → Replication → Assembly → Maturation → Release
- תא רגיש ≠ תא מתירני – נדרשים שניהם להדבקה מוצלחת
- העברה אופקית (בין אנשים) שונה מהעברה אנכית (אם-עובר)
- $R$ (Reproductive Number) – מדד קריטי להבנת התפשטות מגיפות
- PCR – הכפלה ספציפית של $\ce{DNA}$; עבור וירוסי $\ce{RNA}$ נדרשת RT-PCR
- Plaque Assay – השיטה היחידה למדידת וירוסים אינפקטיביים
- לחות וטמפרטורה משפיעות באופן דרמטי על יציבות וירוסים
- Reassortment בשפעת – מנגנון מרכזי לשונות גנטית מהירה