פתיחה: על מה השיעור
בשיעור הקודם הוצגו מספר פקטורי שעתוק חשובים בהתפתחות המוקדמת:
- Oct4
- Nanog
- Cdx2
- Gata4
- Gata6
בשיעור הנוכחי המוקד עובר ל־Cdx2 ולשאלה איך הוא משתתף ב־First cell fate decision, כלומר ביצירת ההבחנה בין תאים חיצוניים שיהפכו ל־Trophectoderm (להלן: TE) לבין תאים פנימיים שיהפכו ל־ICM.
בהמשך השיעור נכנס גם ה־Second cell fate decision, שבו תאי ה־ICM נפרדים לכיוון Epiblast או Primitive endoderm.
Cdx2: מה לומדים מניסוי Loss of Function
כדי להבין את התפקיד של Cdx2, מסתכלים על עובר תקין לעומת עובר מוטנט שבו Cdx2 חסר.
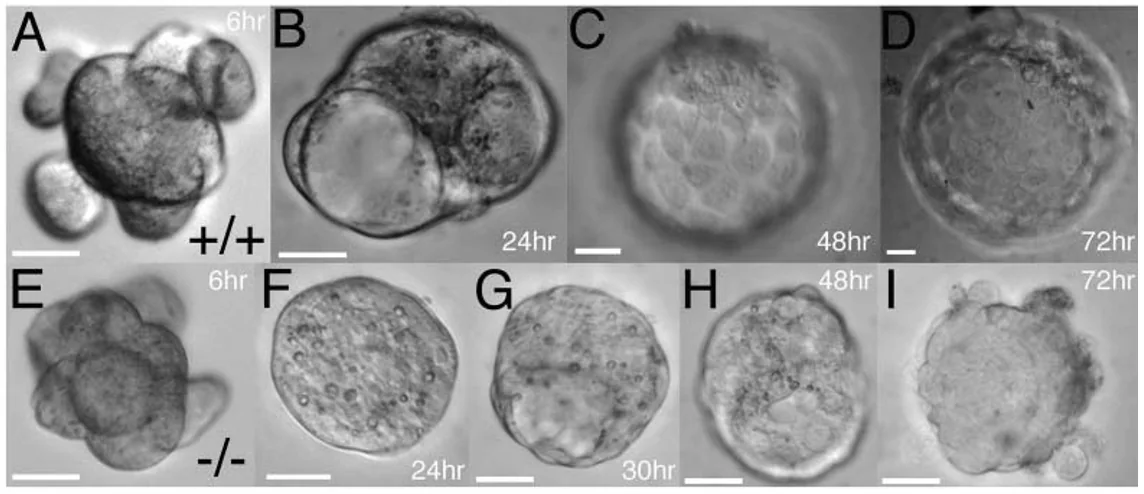
בעובר תקין (השורה למעלה), לאורך הזמן מתקבל בלסטוציסט עם TE, Blastocoel ו־ICM.
במוטנט ל־Cdx2 מתחיל להיווצר Blastocoel, ולכן נראה שהעובר מתחיל להיכנס למסלול של יצירת בלסטוציסט. אבל תוך זמן קצר ה־Blastocoel נעלם, מתרחשת קריסה ומתקבל גוש תאים.
המסקנה הראשונה: ללא Cdx2 העובר מסוגל להתחיל תהליך של יצירת Blastocoel, אבל לא לשמר אותו.
כדי להבין למה זה קורה, צריך להבין קודם איך נוצר ה־Blastocoel ומהו מחסום אפיתליאלי.
תאי אפיתל ו־Tight Junctions
תאי אפיתל הם שכבת תאים שמפרידה בין שני חללים. לדוגמה, תאי אפיתל יכולים להפריד בין חלל מערכת העיכול לבין הגוף, או בין לומן של כלי דם לבין הרקמה שסביבו.
לתאי אפיתל יש בדרך כלל שני צדדים:
| צד בתא | משמעות |
|---|---|
| Basal / Basolateral domain | הצד שפונה לממברנה בזלית או לתאים שכנים |
| Apical domain | הצד שפונה לחלל או לסביבה החיצונית |
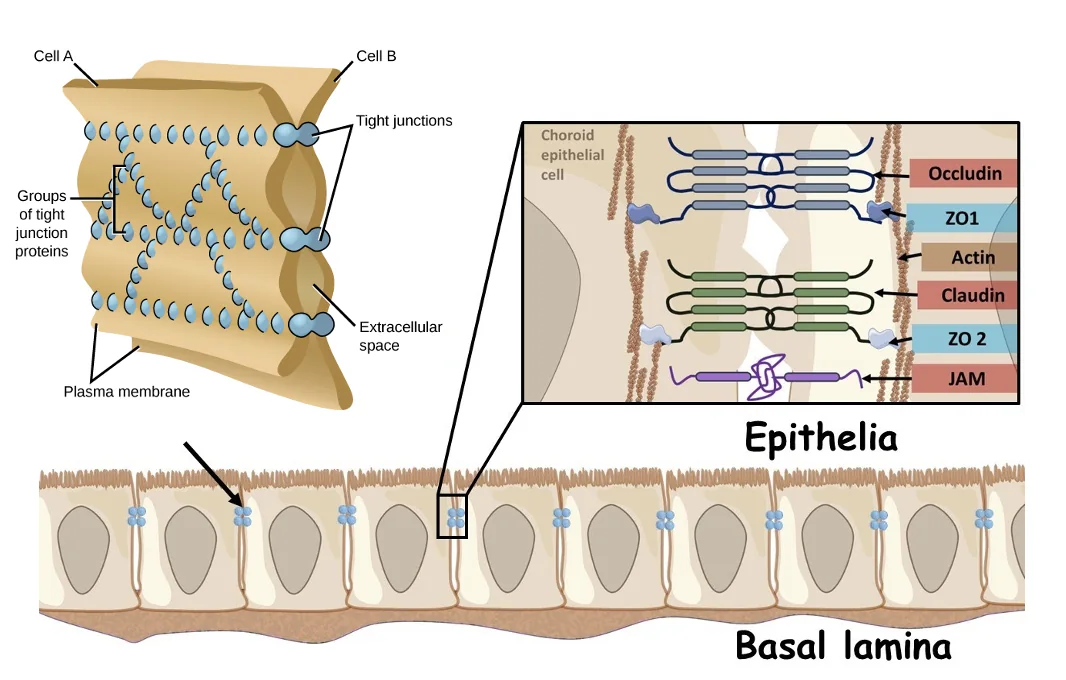
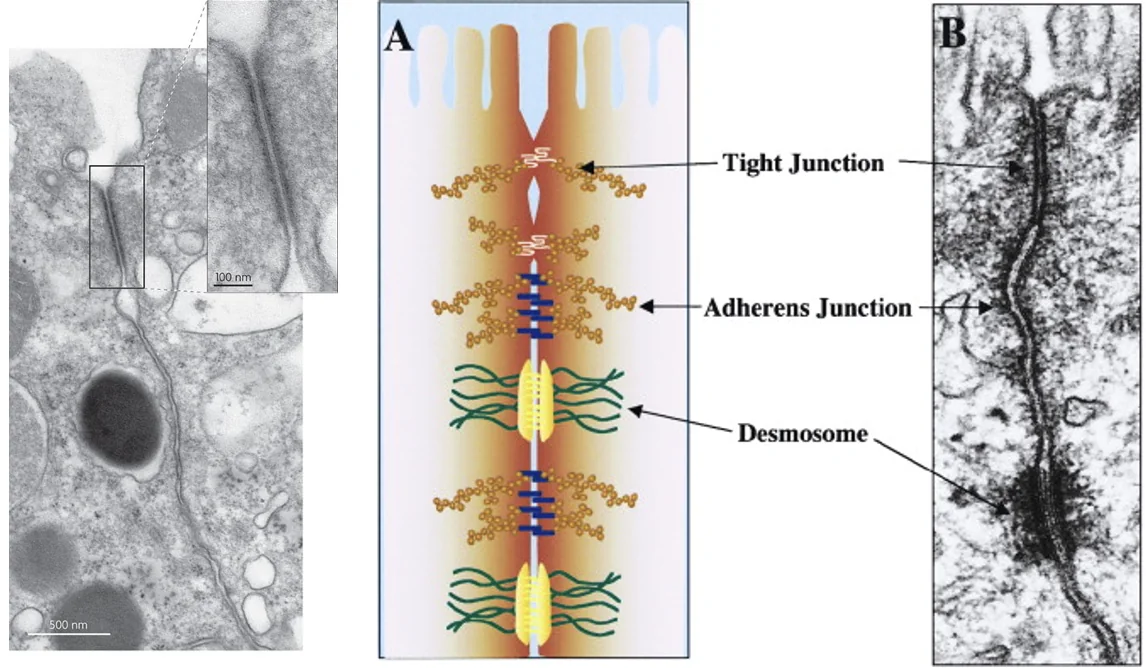
בקצה האפיקלי של תאי אפיתל יכולים להופיע Tight junctions. אלה קומפלקסים של חלבונים שמצמידים את הממברנות של שני תאים שכנים, כך שהמעבר בין התאים נחסם.
ההבדל בין שני סוגי החיבורים:
| מבנה | תפקיד עיקרי בשיעור |
|---|---|
| Adherens junctions | הצמדה מכנית בין תאים, בין היתר דרך E-cadherin |
| Tight junctions | יצירת מחסום שמונע מעבר בין התאים |
הנקודה החשובה היא ש־Tight junctions לא מאפשרים למים, יונים או חומרים אחרים לעבור בין התאים. אם משהו עובר מצד אחד לצד השני, הוא צריך לעבור דרך התא עצמו ולא בין התאים.
יצירת ה־Blastocoel
 | 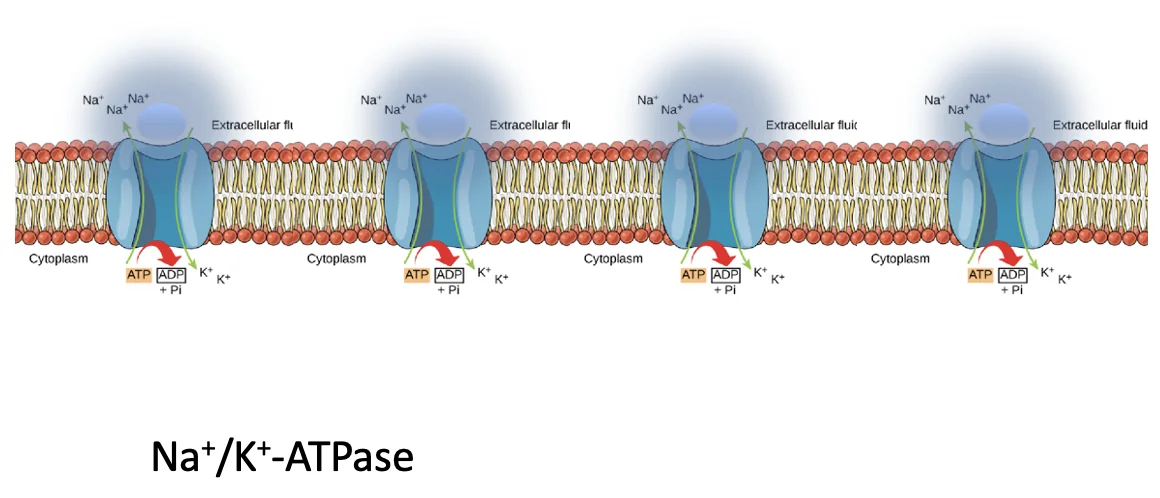 |
בשלב המורולה יש תאים פנימיים ותאים חיצוניים; כל התאים מחוברים זה לזה דרך Adherens junctions, אבל רק התאים החיצוניים מתנהגים כתאים אפיתליאליים ויוצרים Tight junctions בצד האפיקלי שלהם.
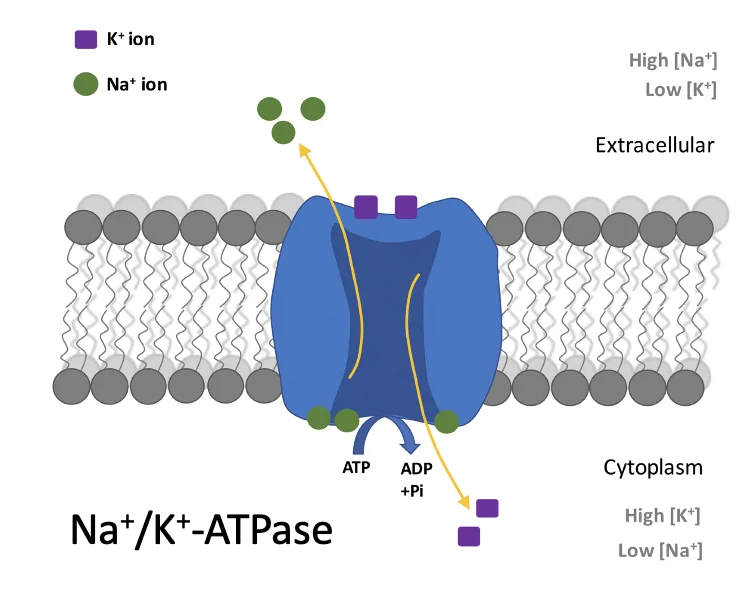
המשאבה בתמונה למעלה היא חלבון טראנסממברנלי (אנטיפורטר), בצד הבזו־לטרלי של התאים (בין תאים סמוכים ולא בצד האפיקלי). היא מוציאה שלושה יוני נתרן ומכניסה שני יוני אשלגן תוך שימוש ב־ATP. החשיבות כאן היא בעיקר בשינוי ריכוז היונים (ולא המטען החשמלי): כשריכוז היונים עולה באזור שבין התאים (מחוץ לתא), נוצר לחץ אוסמוטי שמושך מים מהתא החוצה.
הסדר הוא כזה (ראו תמונה מטה):
- המשאבות יוצרות ריכוז יונים גבוה יותר באזור הבזו־לטרלי, בין התאים (התוצר נטו הוא מינוס יון אחד).
- מים נעים דרך התאים (החוצה) בעקבות הלחץ האוסמוטי.
- נוצרים חללים קטנים של נוזל בין התאים.
- החללים הקטנים נעים, מתאחדים ויוצרים חללים גדולים יותר.
- בסופו של דבר נוצר חלל מרכזי אחד - Blastocoel.
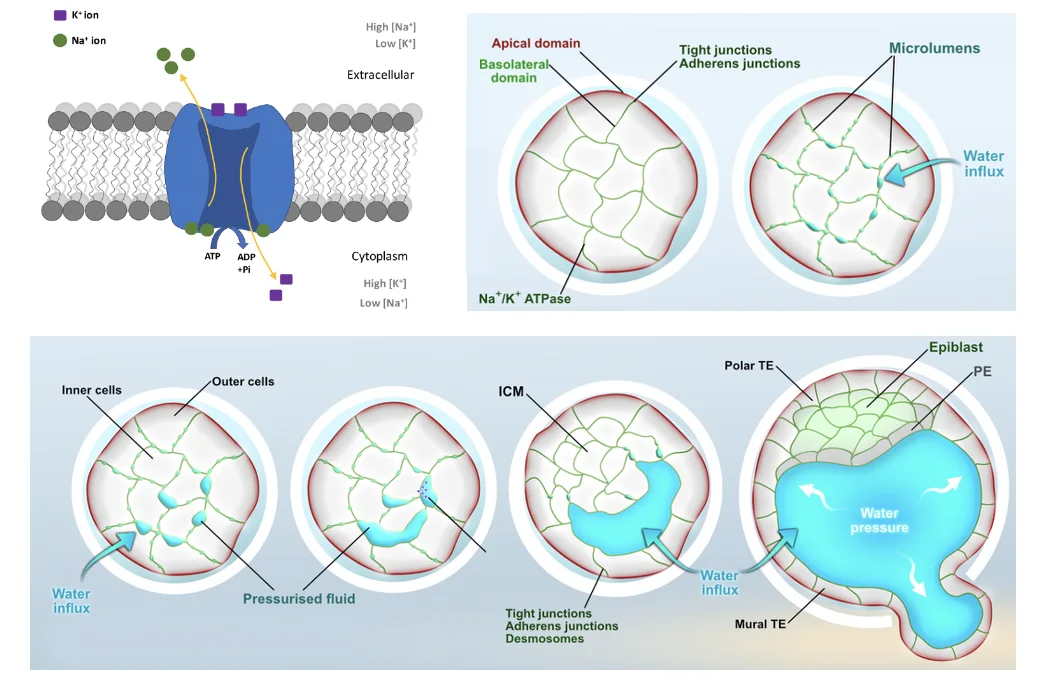
ה־Tight junctions חשובים משום שהם סוגרים את העובר כמערכת. אם לא הייתה סגירה (מחסום), הנוזלים היו יכולים לברוח החוצה, והחלל לא היה נשמר.
למה ה־Blastocoel חשוב?
ככל שה־Blastocoel גדל, הוא יוצר לחץ מבפנים כלפי חוץ. בשלב היציאה מה־Zona Pellucida, העובר מפריש פרוטאזות באזור מסוים ויוצר פתח קטן ב־Zona Pellucida. הפתח לבדו אינו מספיק; הלחץ שנוצר על ידי ה־Blastocoel דוחף את העובר החוצה.
ה־Zona Pellucida קשיחה, אבל מים יכולים לעבור דרכה. מה ששומר עליהם הוא המחסום שיוצרים התאים החיצוניים, בעזרת ה־Tight junctions.
מה קורה ב־Cdx2 KO?
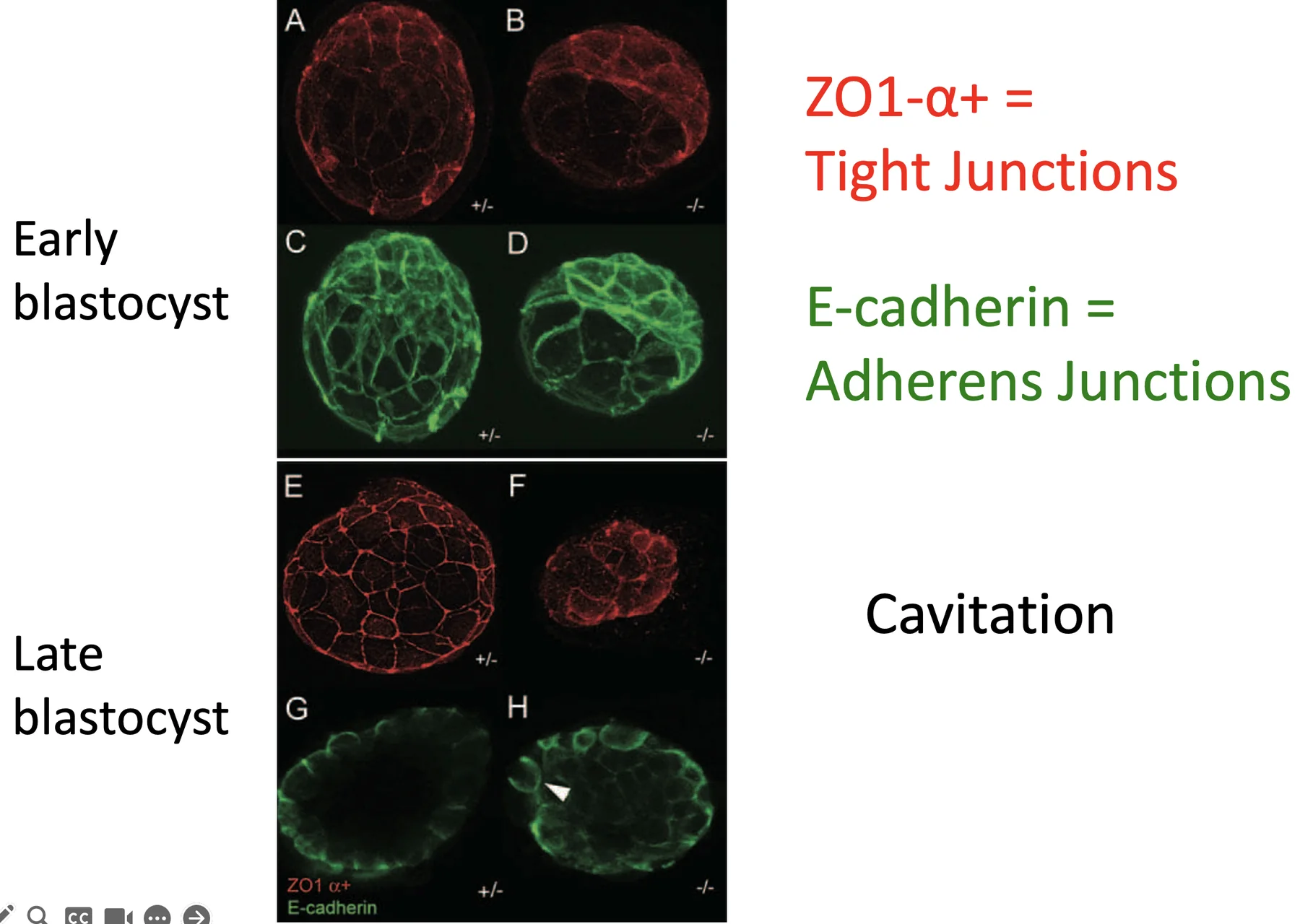
בתמונה, העוברים התקינים משמאל (A, C, E, G), והמוטנטים ל־Cdx2 מימין (B, D, F, H).
בבלסטוציסט מוקדם, גם במוטנט ניתן לראות התחלה של Tight junctions ו־E-cadherin. אבל בבלסטוציסט מאוחר, בעובר התקין יש Blastocoel ברור ומחסום אפיתליאלי יציב, ואילו במוטנט ה־Tight junctions מתפרקים או נעשים לא תקינים.
המשמעות היא שה־Blastocoel מתחיל להיווצר, אבל אינו מוחזק. ברגע שה־Tight junctions לא יציבים, הנוזלים אינם נשמרים, והחלל קורס.
ללא Cdx2, התאים החיצוניים (outer cells) מאבדים את התכונות האפיתליאליות הדרושות לשימור ה־Blastocoel.
ניסוי תרבית: Outgrowth של TE
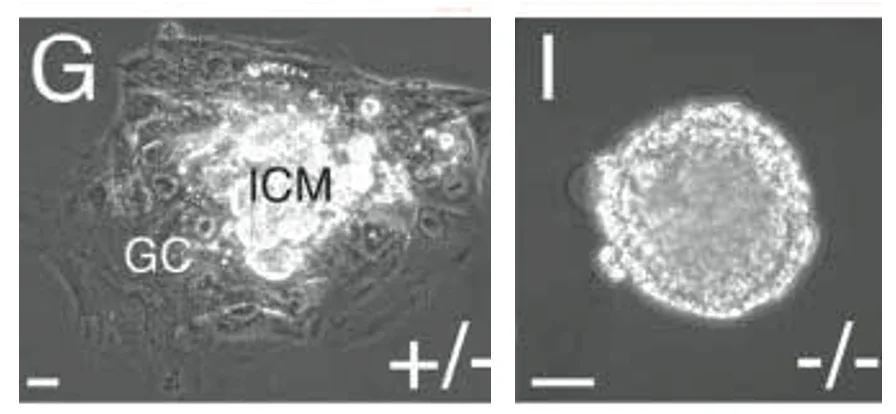
בניסוי נוסף לוקחים את העובר ושמים אותו בצלחת בתנאי תרבית.
בעובר תקין, ה־TE נפרס על הצלחת, וה־ICM יושב מעליו. במוטנט ל־Cdx2, נראה שאין TE מתפקד, אלא בעיקר גוש תאים שנראה כמו ICM (צד ימין בתמונה למעלה).
Cdx2 מדכא Oct4 ו־Nanog ב־TE
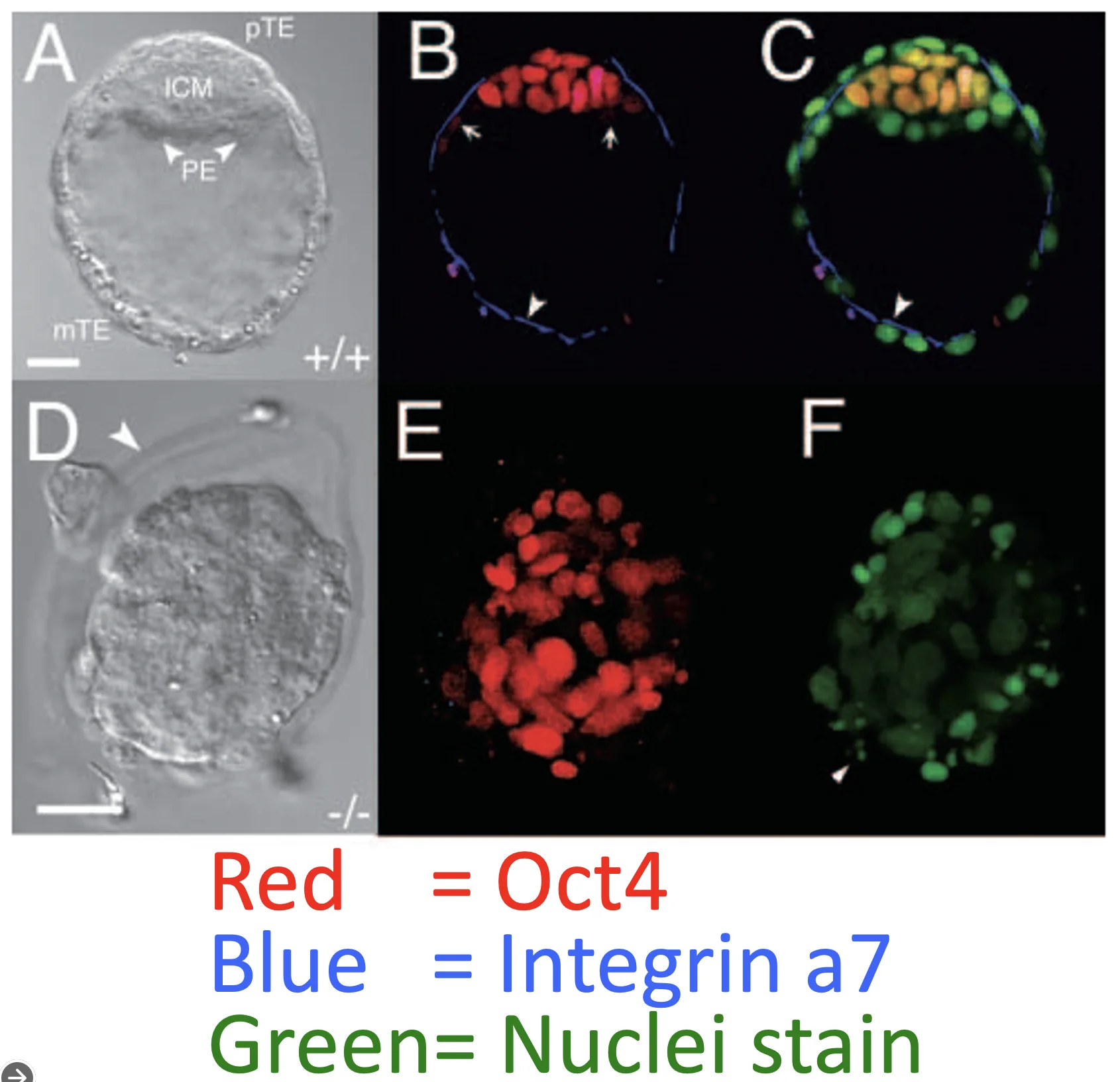
בצביעה שבתמונה השתמשו ב־Integrin α7 (צבע כחול) כמרקר של TE, וב־Oct4 (צבע אדום) כמרקר שקשור ל־ICM.
במוטנטים ל־Cdx2, כמעט כל התאים מבטאים Oct4 (צבע אדום בתמונה E), בעוד שהמרקר ל־TE אינו מופיע כמו בעובר התקין (בתמונה E לא רואים את העיגול הכחול שרואים ב־B). המשמעות היא שאין הפרדה תקינה בין תאי ICM לבין TE.
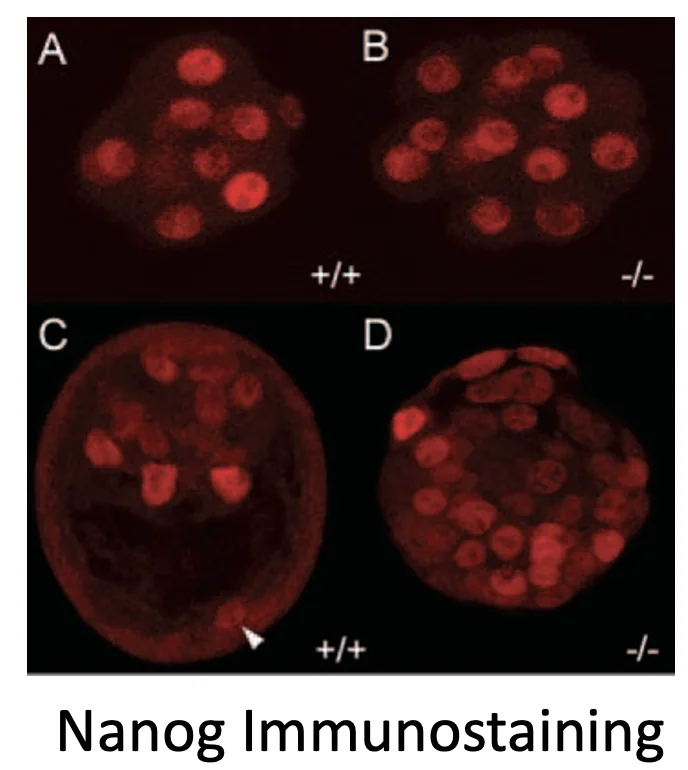
גם בבחינה של Nanog מתקבלת תמונה דומה: במוטנט, תאים רבים מבטאים Nanog.
המסקנה החשובה: Cdx2 הוא רפרסור של Oct4 ושל Nanog בתאי ה־TE.
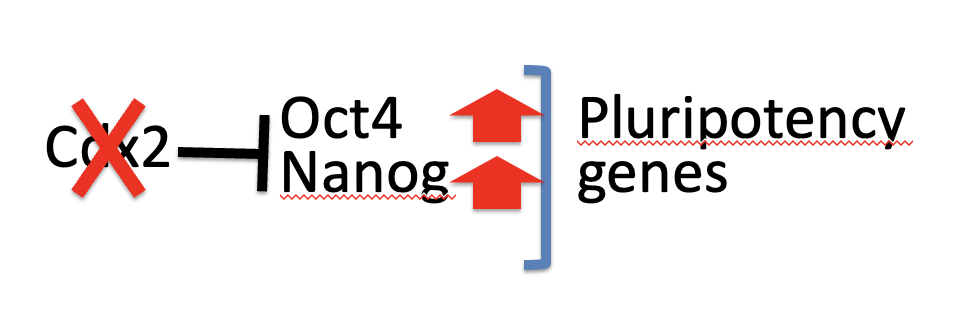
כלומר, כדי שתא חיצוני יהפוך ל־TE, צריך למנוע בו ביטוי של Oct4 ו־Nanog. לעומת זאת, כדי שתא יהיה ICM, צריך להוריד את Cdx2 ולאפשר ל־Oct4 ול־Nanog להתבטא (כמו בתמונה למעלה).
Compaction
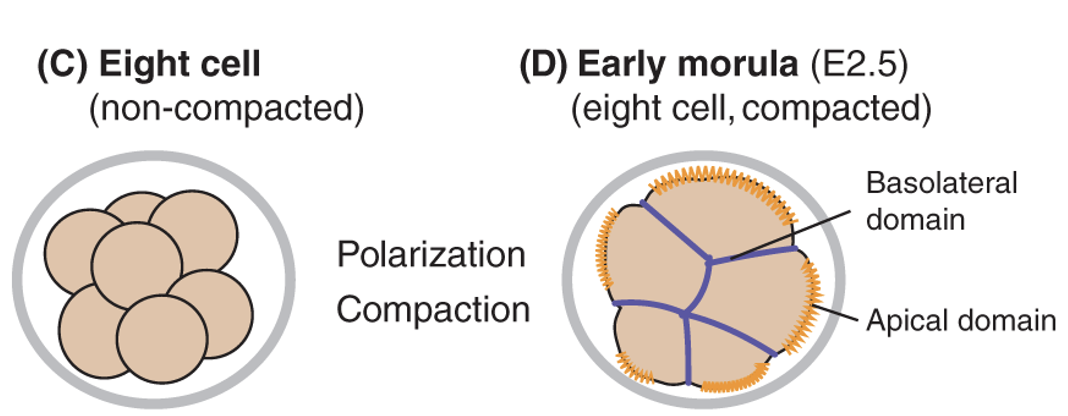
במעבר סביב שלב שמונת התאים מתרחשים שני תהליכים חשובים:
- Compaction - התאים נצמדים זה לזה ונראים פחות כבלסטומרים נפרדים.
- Polarization - התאים מקבלים צד אפיקלי וצד בזולטרלי.
ב־Compaction, הבלסטומרים נצמדים זה לזה בעיקר דרך Adherens junctions, שבהם משתתף E-cadherin.
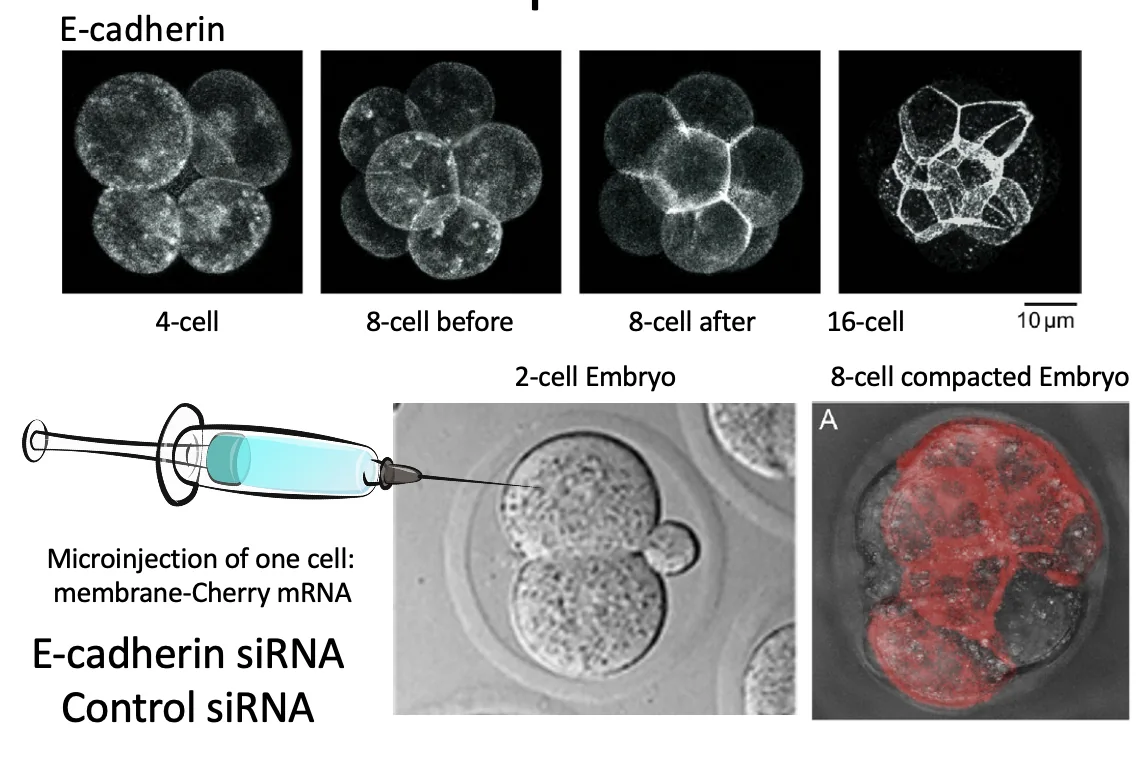
בתחילת הדרך E-cadherin נמצא על פני הממברנה, אבל לאחר Compaction הוא מתרכז באזורי המגע בין התאים. עצם הנוכחות של E-cadherin על הממברנה לא מספיקה; צריך שייווצרו בין התאים Adherens junctions פעילים.
siRNA וניסוי E-cadherin
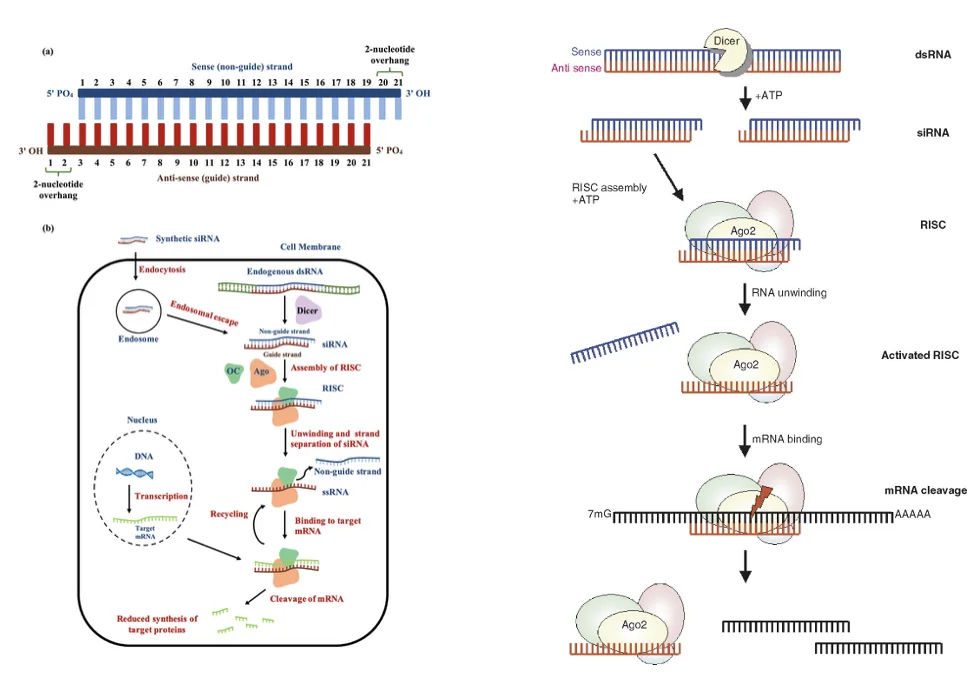
העיקרון של siRNA: מולקולות RNA דו־גדיליות קצרות יכולות להוביל לפירוק RNA מטרה ספציפי, וכך להוריד ביטוי של גן מסוים. המנגנון הביולוגי כולל חיתוך dsRNA על ידי Dicer, ובהמשך שימוש בקומפלקס שמזהה את ה־RNA המתאים ומוביל לפירוקו.
בניסוי הזה מזריקים לתא אחד בלבד בעובר שנמצא בשלב של שני תאים mRNA שמקודד לסמן אדום. מכיוון שרק תא אחד קיבל את ה־mRNA, רק הצאצאים שלו יהיו מסומנים באדום.
אחרי שהעובר ממשיך להתחלק, בודקים איפה נמצאים התאים האדומים. אם רואים שחצי מהעובר אדום וחצי מהעובר לא אדום, המשמעות היא שהצאצאים של כל אחד משני התאים המקוריים נשארו יחסית יחד. כלומר, התאים לא התערבבו באקראיות מלאה בתוך העובר.
במילים פשוטות: הסמן האדום משמש למעקב אחרי “משפחה” אחת של תאים, ומראה שבשלבים האלה עדיין אפשר לזהות מאיזה תא מוקדם הם הגיעו.
השלב הבא הוא להזריק גם siRNA נגד E-cadherin.
הביקורת חשובה: ייתכן שההזרקה עצמה (הסמן האדום), או עצם הכנסת siRNA גורמים להשפעה. לכן משתמשים גם ב־siRNA אקראי שאינו מכוון נגד גן ספציפי.
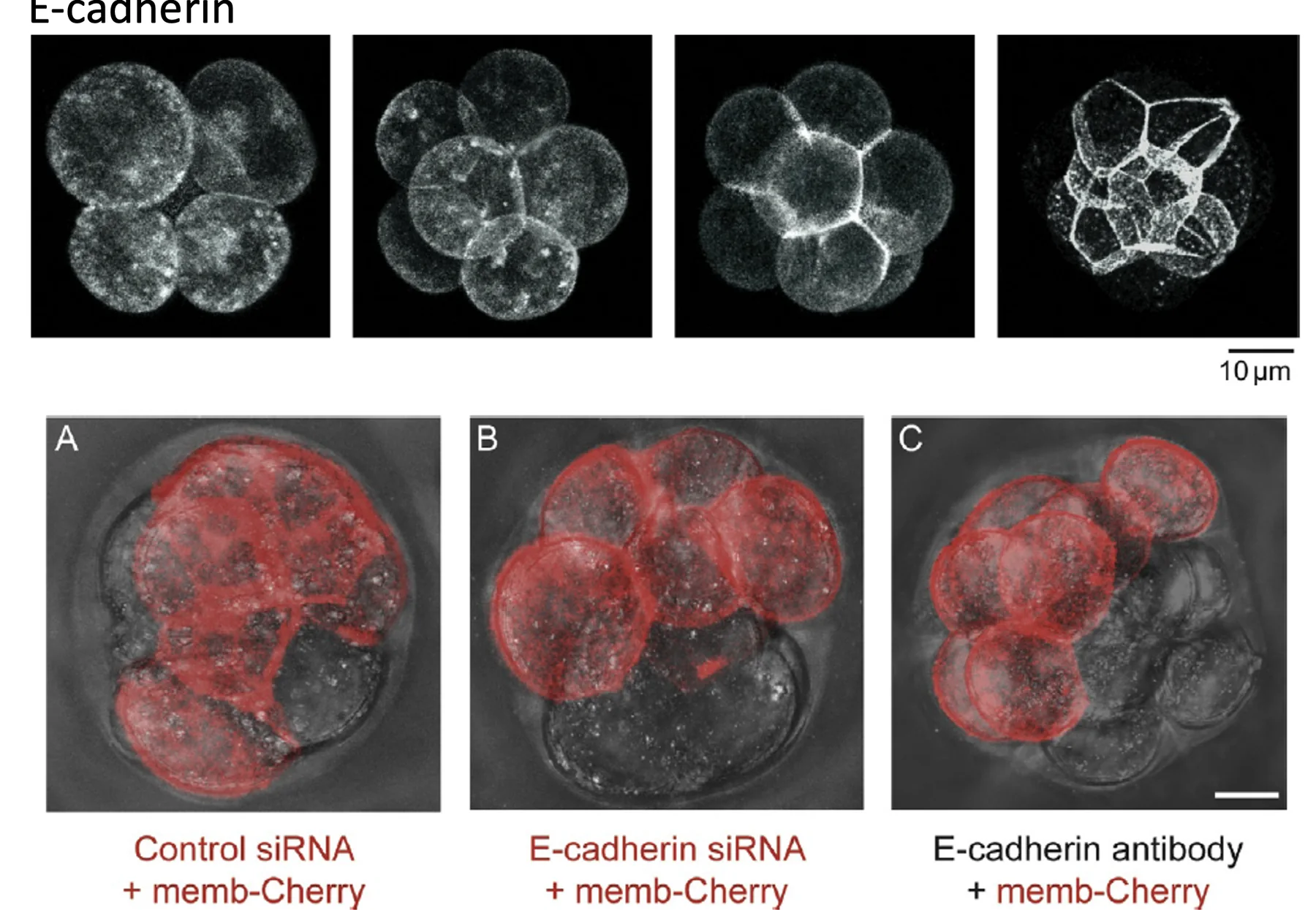
התוצאות:
| טיפול | מה רואים? | מסקנה |
|---|---|---|
siRNA ביקורת  | התאים המסומנים משתלבים בעובר שעובר Compaction | עצם ההזרקה אינה מספיקה כדי לשבש Compaction |
siRNA נגד E-cadherin 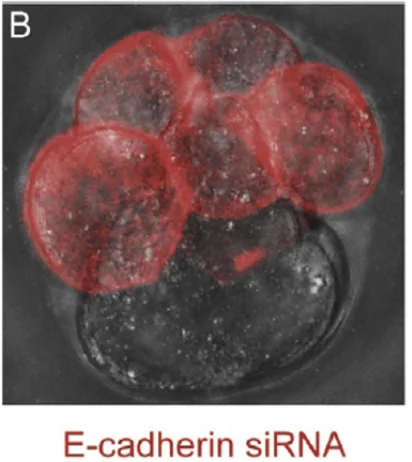 | התאים שבהם הורד E-cadherin נשארים עגולים ואינם משתלבים היטב | E-cadherin דרוש ל־Compaction |
נוגדן חוסם נגד E-cadherin 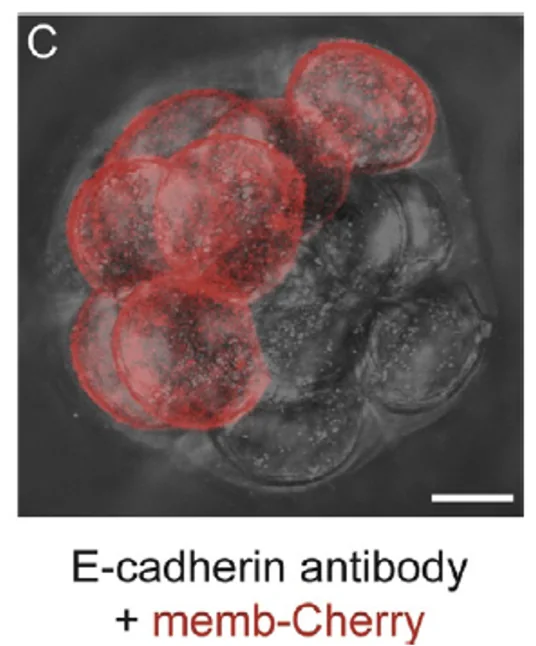 | כל התאים נפגעים ולא עוברים Compaction תקין | הפעילות הישירה של E-cadherin חשובה לתהליך |
איך Compaction נוצר בפועל?
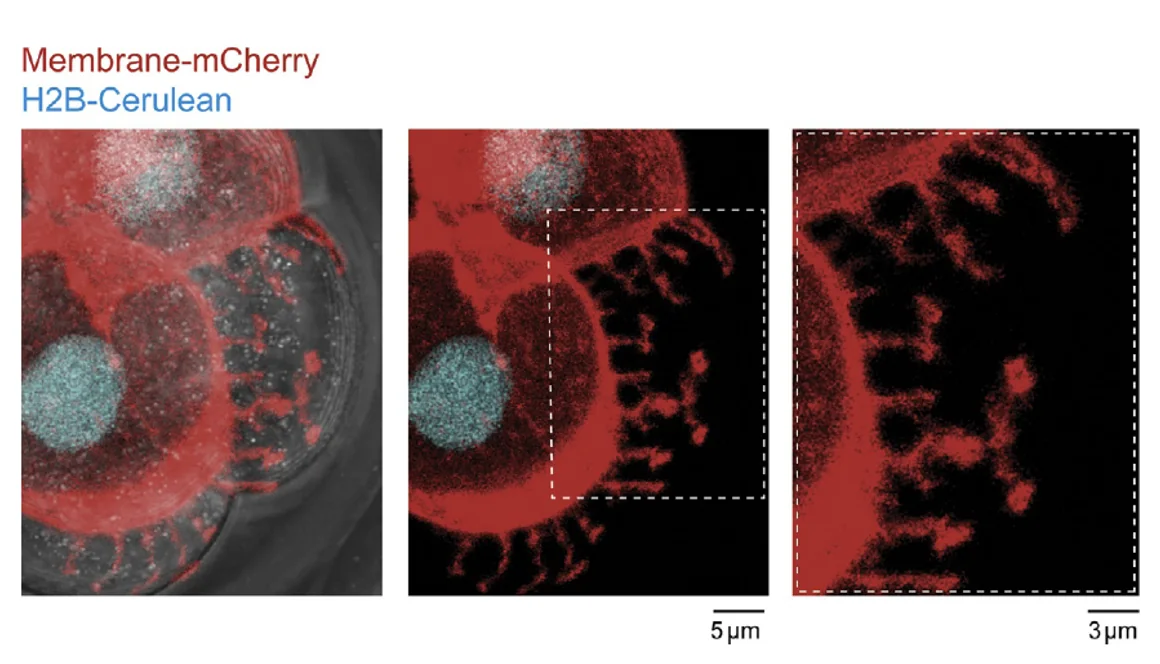
התאים שולחים שלוחות דמויות זרועות, filopodia, שתופסות את הממברנה של תאים שכנים ומקרבות אותם. התהליך תלוי ב־E-cadherin.
Polarization
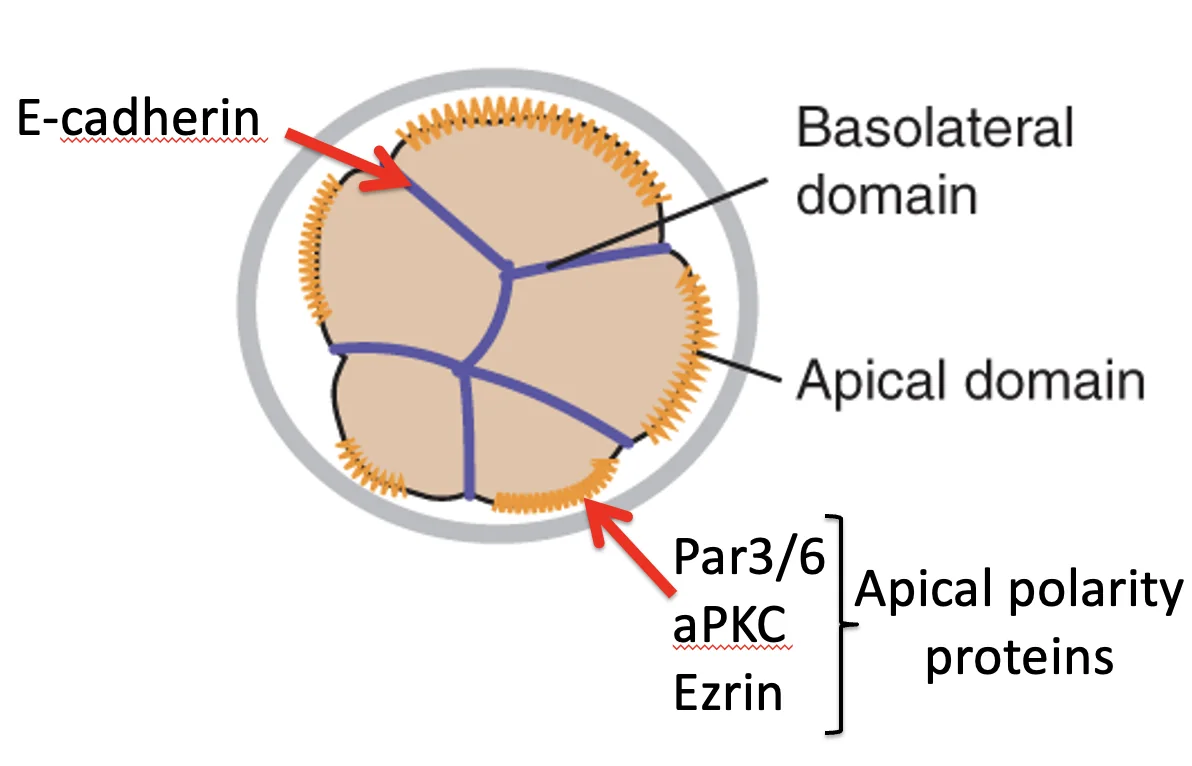
במקביל ל־Compaction, התאים עוברים Polarization. המשמעות היא שהתא כבר אינו אחיד מכל צדדיו: בצד אחד יש קומפלקס אפיקלי, ובצד אחר יש מגעים עם תאים שכנים.
בצד האפיקלי מופיעים חלבונים כמו Par3, Par6, aPKC ו־Ezrin. הם יוצרים את ה־Apical polarity complex.
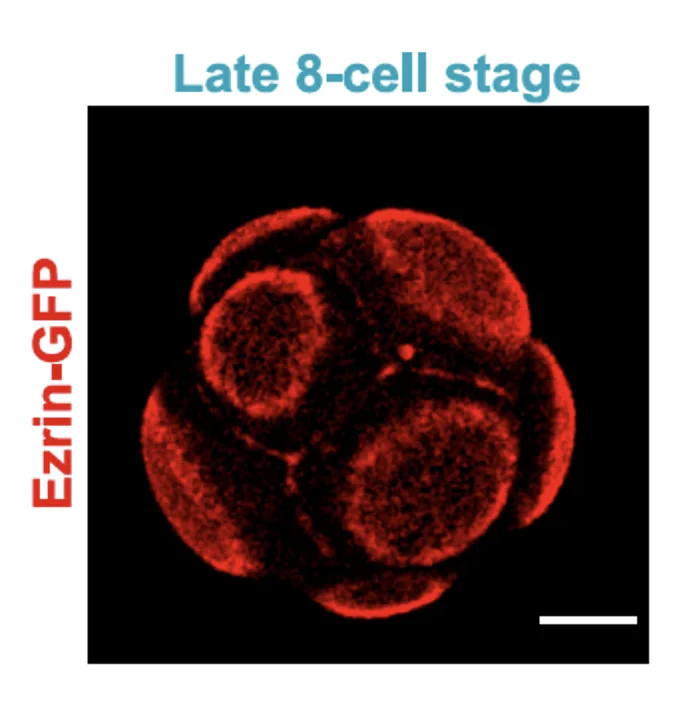
בצביעה של Ezrin ניתן לראות מעין כיפה אפיקלית, שמייצגת את האזור שבו מרוכזים חלבוני הפולריות האפיקלית.
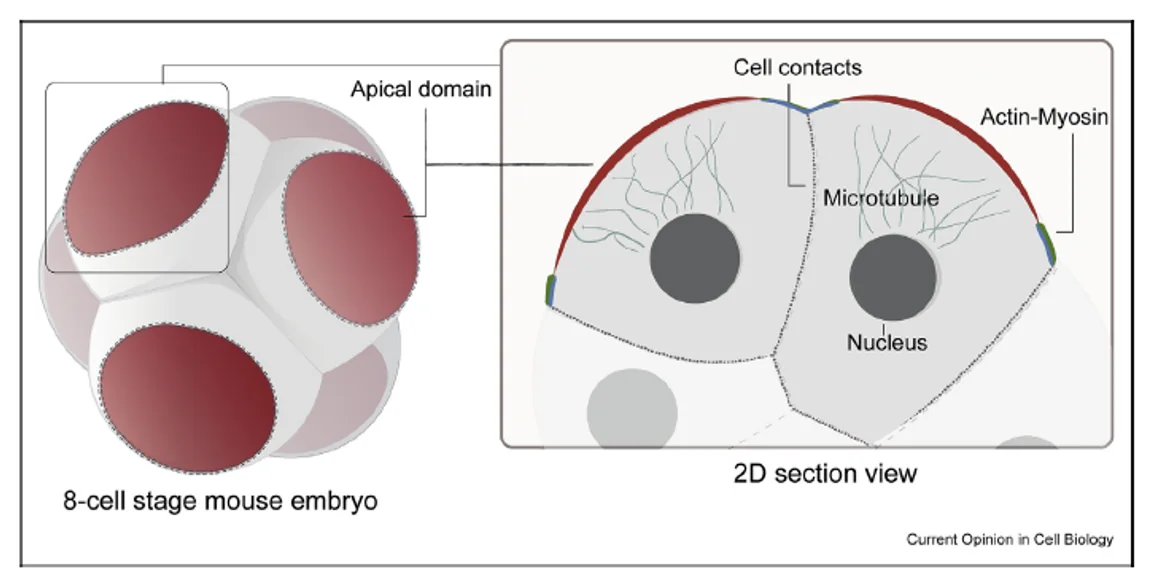
הכיפה האפיקלית מוקפת בטבעת אקטין-מיוזין. בנוסף לכך, המיקרוטובולים מכוונים מהגרעין לכיוון האזור האפיקלי.
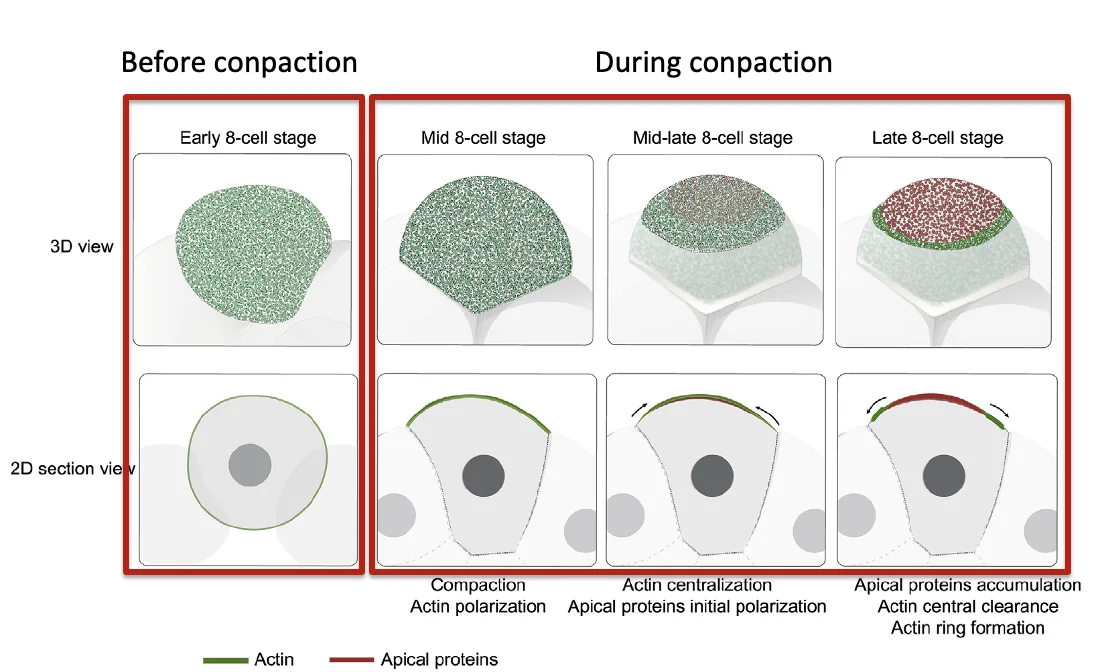
בהתחלה, אקטין מתרכז באזור החופשי ממגע עם תאים אחרים. בהמשך, חלבוני הפולריות האפיקלית מתרכזים במרכז האזור, והאקטין נדחק לפריפריה ויוצר טבעת.
Compaction ו־Polarization קשורים זה לזה, אבל אינם תלויים לחלוטין זה בזה.
First cell fate decision: תאים חיצוניים מול תאים פנימיים
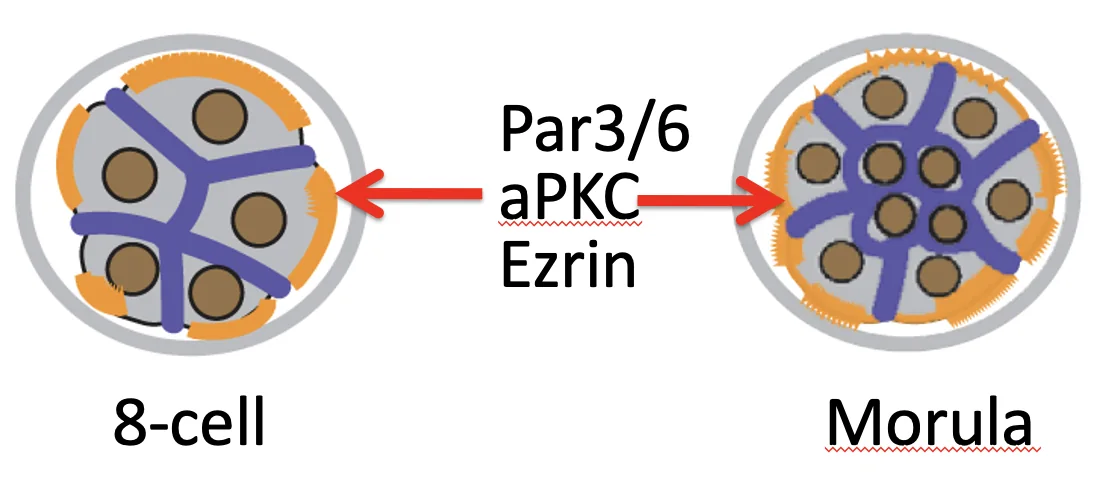
בעובר בן שמונה תאים, לפני ההבחנה בין תאים פנימיים וחיצוניים, כל התאים פולריים (כל התאים פונים מצד אחד ל־ZP ומשאר הצדדים לתאים סמוכים). לאחר מכן מתקבל מצב שבו:
| אוכלוסייה | מצב פולריות | גורל עיקרי |
|---|---|---|
| תאים חיצוניים | פולריים | TE |
| תאים פנימיים | לא פולריים | ICM |
השאלה המרכזית היא איך עוברים ממצב סימטרי יחסית, למצב שבו יש שתי אוכלוסיות תאים שונות.
שתי היפותזות
| היפותזה | רעיון מרכזי |
|---|---|
| Inside-outside | המיקום קובע את הפולריות: תא שבחוץ יהיה פולרי, תא שבפנים יהיה לא פולרי |
| Polarity | הפולריות קובעת את המיקום: תא פולרי יישאר בחוץ, תא לא פולרי ייכנס פנימה |
ניסויים שמראים תמיכה בשני הכיוונים:
Reciprocal positional change (לתמיכה בהיפותזת Inside-outside):
- אם מזיזים פיזית תא חיצוני פנימה, הוא יכול לאבד פולריות.
- אם מזיזים תא פנימי החוצה, הוא יכול להפוך לפולרי.
Downregulation of Par/6 and aPKC in one cell (לתמיכה בהיפותזת Polarity):
- אם מורידים חלבון של הקומפלקס האפיקלי (למשל בעזרת siRNA), למשל Par3, התא יכול לעזוב את השכבה החיצונית ולהיכנס פנימה.
- אם גורמים לתא פנימי לבטא מרכיבי פולריות, הוא יכול להפוך לפולרי ולצאת החוצה.
המסקנה: שליטה בפולריות מאפשרת שליטה במיקום התא, ולכן היא חלק מרכזי ב־First cell fate decision. שתי ההיפותזות נכונות.
מישור החלוקה: חלוקה סימטרית וא-סימטרית
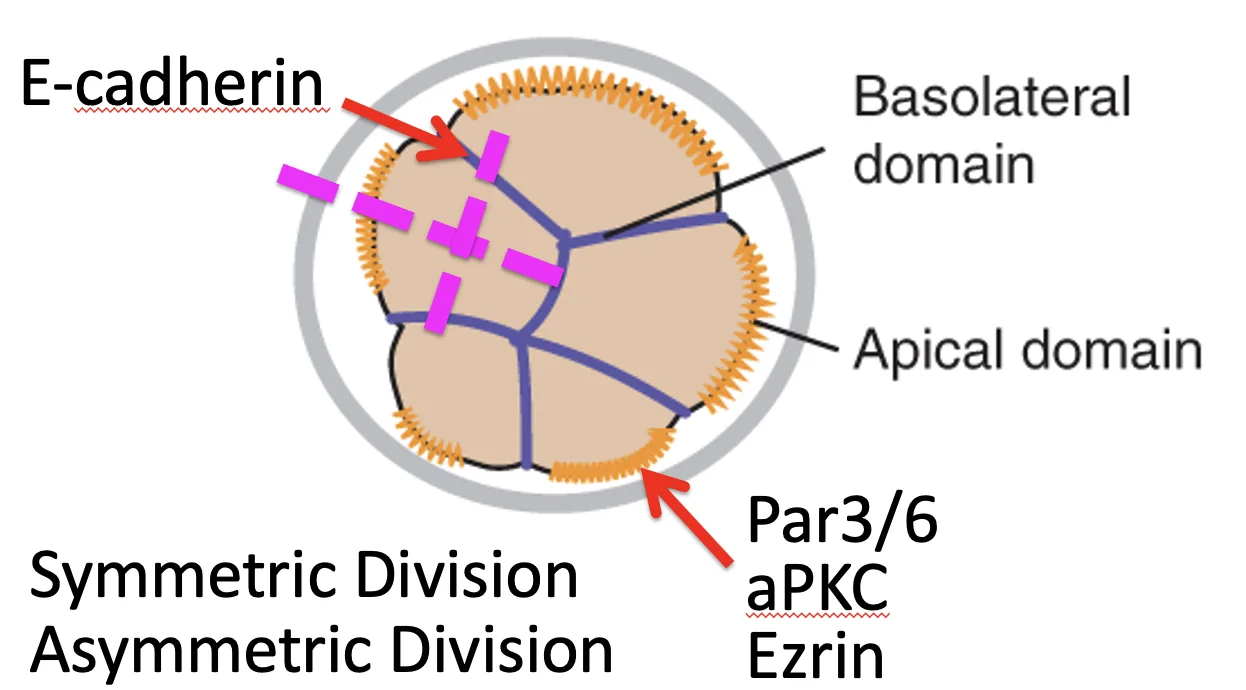
מלבד פולריות, יש פרמטר נוסף לשליטה במיקום (ומכאן בסימטריה, או ליתר דיוק בהחלטת הגורל הראשונה ששוברת אותה): מישור החלוקה.
| סוג חלוקה | תוצר |
|---|---|
חלוקה סימטרית 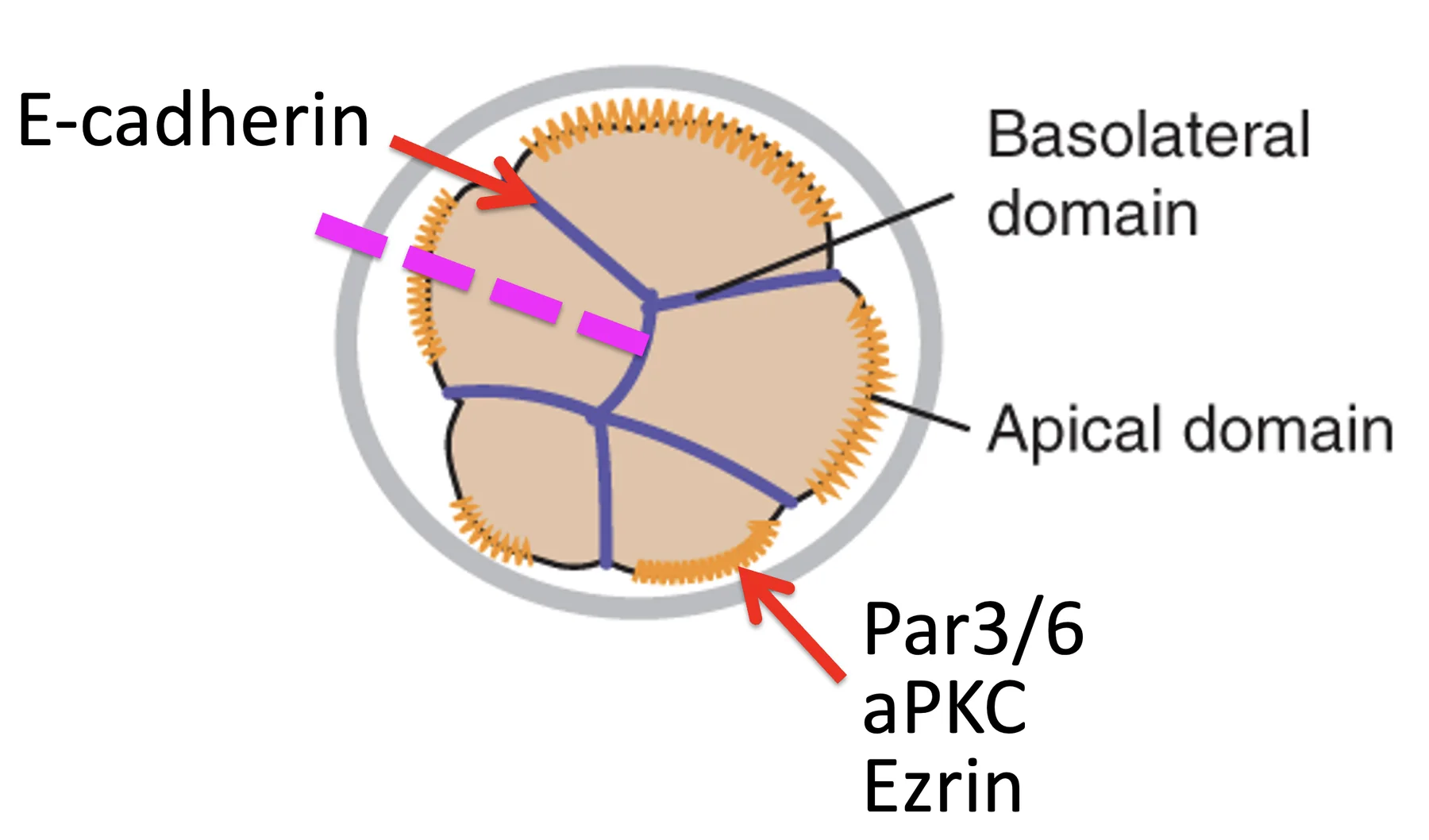 | שני תאי בת דומים, שניהם נשארים חיצוניים ופולריים |
חלוקה א-סימטרית 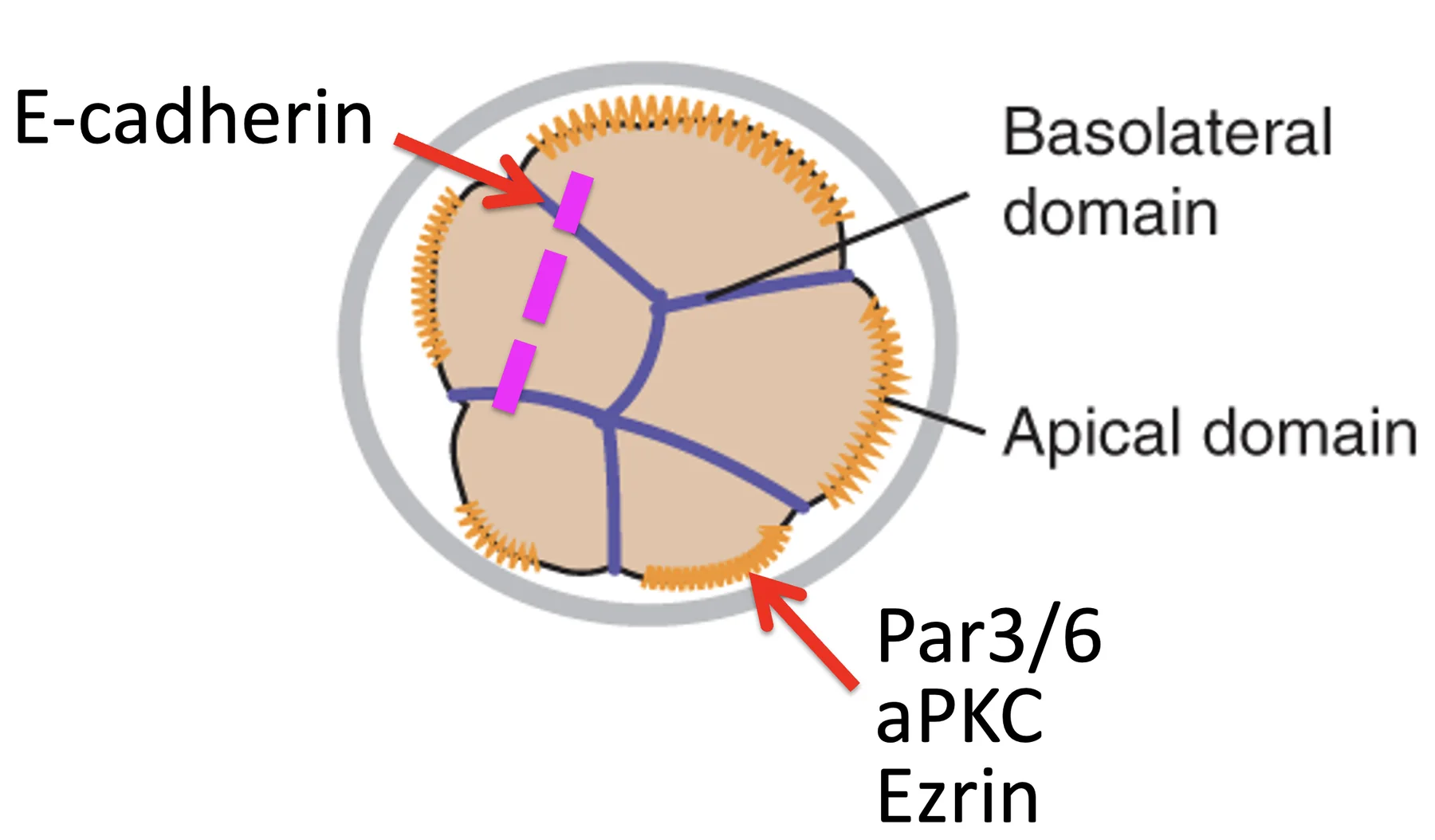 | תא בת אחד מקבל את הקומפלקס האפיקלי ונשאר חיצוני, ותא בת שני אינו מקבל אותו ונכנס פנימה |
לכן, כדי לבצע First cell fate decision צריך לשלוט בשני פרמטרים:
- פולריזציה
- מישור החלוקה
Cdx2, פולריות ולוקליזציית mRNA
החלק הבא של הדיון עוסק בשאלה איך ניתן לשלוט בפולריות.
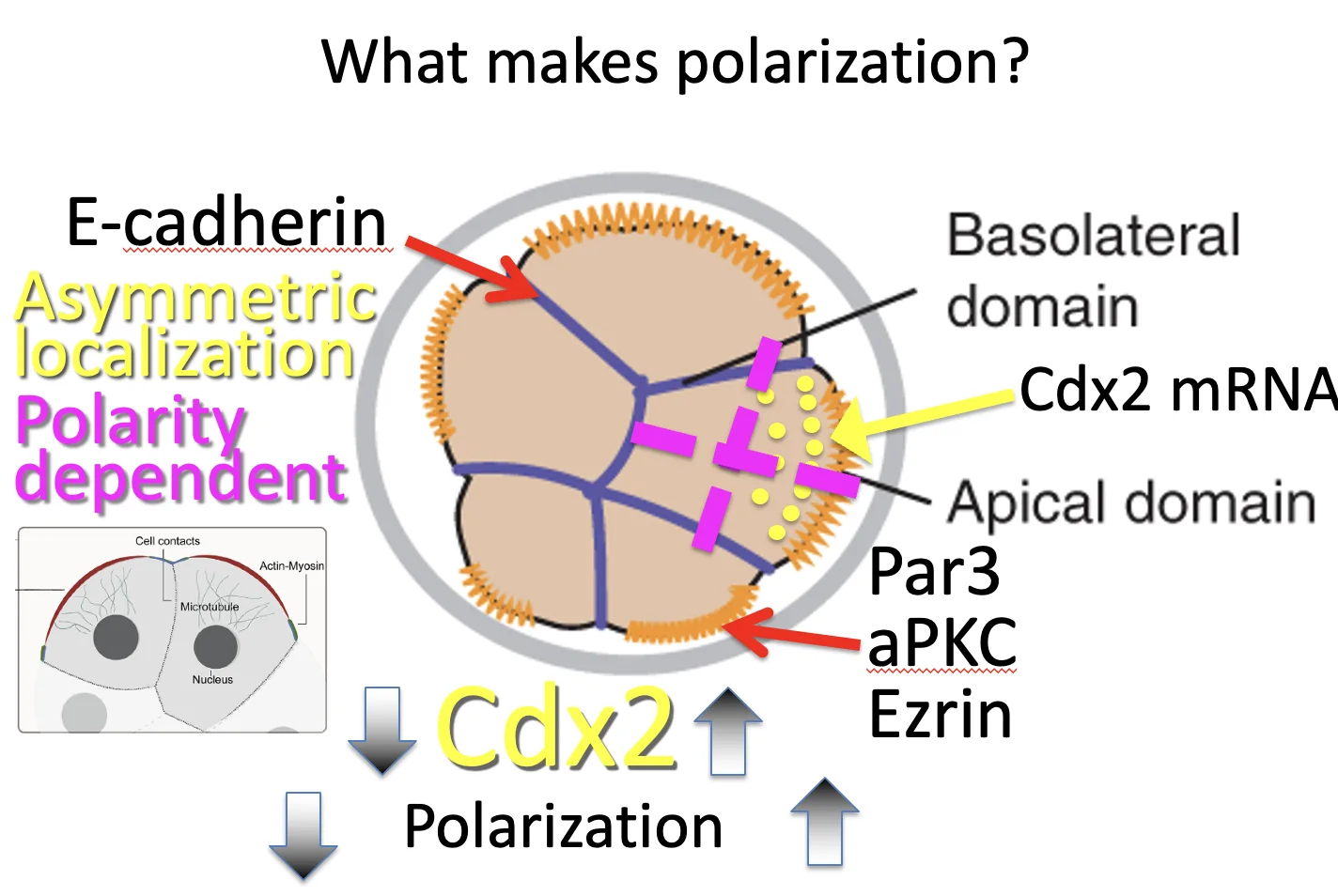
יש קשר בין רמות Cdx2 ובין מידת הפולריות של התא:
- יותר Cdx2 ← יותר פולרי
- פחות Cdx2 ← פחות פולרי
לא צריך לחשוב על פולריות כמצב בינארי (״פולרי״ או ״לא פולרי״), אלא כספקטרום. ככל שיהיה לתא יותר Cdx2, הוא יהיה יותר פולרי (וכתוצאה מכך יישאר בחוץ בסבירות גבוהה יותר, להבנתי). ככל שיהיה לו פחות Cdx2, הוא יהיה פחות פולרי, ויותר סיכוי שייכנס פנימה.
בנוסף לכך, ה־mRNA של Cdx2 לא מפוזר בציטופלזמה באופן אחיד, אלא ממוקם באזור האפיקלי של התא. זו לוקליזציה א-סימטרית (Asymmetric localization) של mRNA. בתמונה למעלה ניתן לראות את Cdx2 בנקודות הצהובות.
כשיש שעתוק נוצר RNA, לאחר מכן מתבצע שחבור (splicing) ואז ה־RNA יוצא מהגרעין, אל הציטופלזמה. אבל מסתבר שהפיזור שלו בציטופלזמה לא בהכרח אחיד.
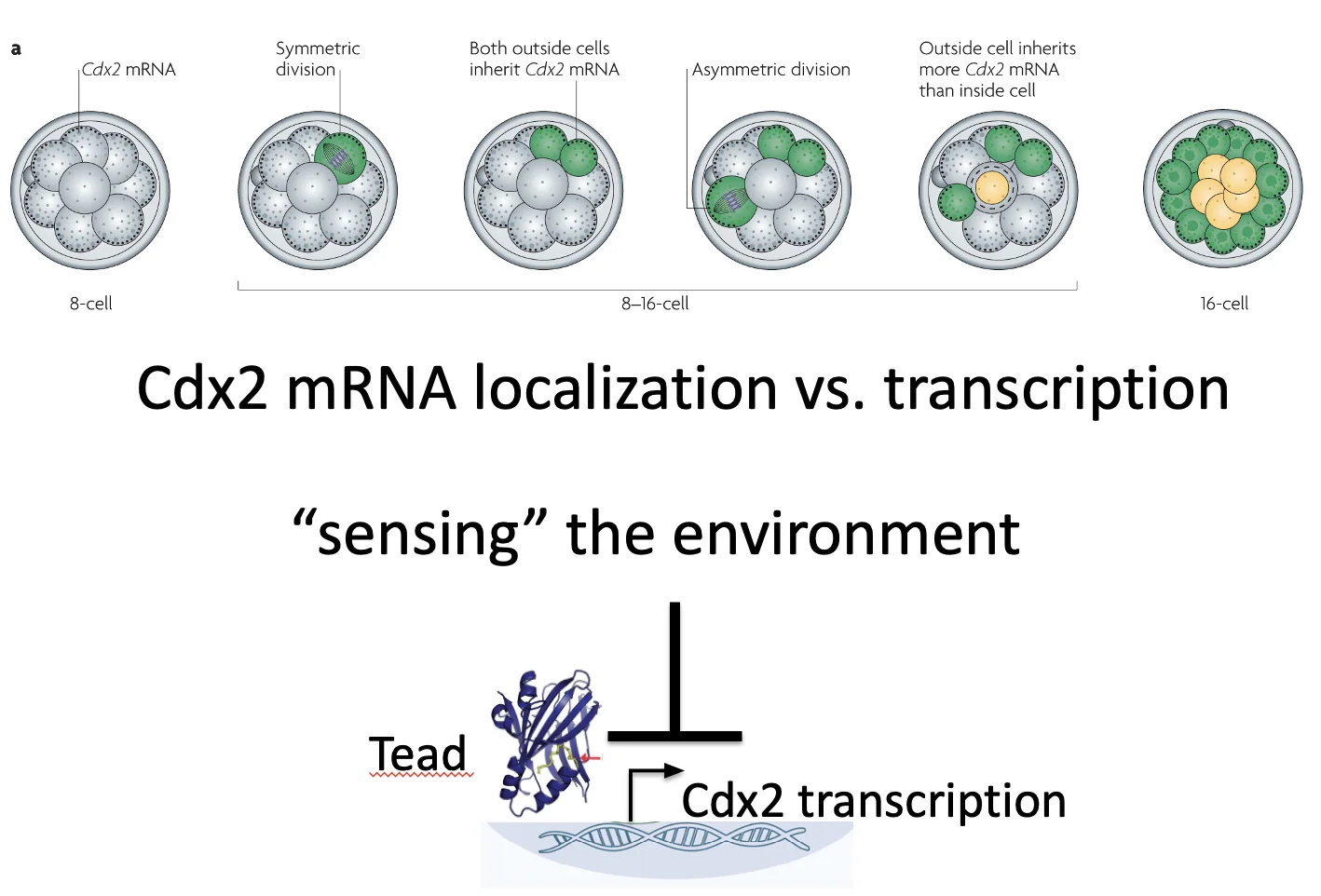
חשוב להפריד בין שני דברים: מיקום ה־mRNA של Cdx2, ושעתוק Cdx2.
| רכיב | איפה הוא רלוונטי? | משמעות |
|---|---|---|
| Cdx2 mRNA | ממוקם בצד האפיקלי (בציטופלזמה) | משפיע על מה כל תא בת יקבל בחלוקה |
| Cdx2 protein | פועל בגרעין | פקטור שעתוק שמפעיל תוכנית של TE |
כשתא פולרי מתחלק א-סימטרית, תא הבת החיצוני מקבל יותר Cdx2 mRNA, ואילו תא הבת הפנימי מקבל מעט מאוד ממנו. כך נוצר חלון זמן שבו בתא הפנימי רמות Cdx2 נמוכות.
אבל זה לא מספיק: אם התא הפנימי ימשיך לשעתק Cdx2, הרמות יעלו שוב והתא עלול להפוך לפולרי ולחזור החוצה (לולאת משוב חיובי בין פולריזציה ו־Cdx2 שמובילה ליצירת TE). לכן, התא הפנימי צריך גם לכבות את השעתוק של Cdx2 (דרך עיכוב TEAD, כפי שנראה בהמשך), כדי לשמור על רמות Cdx2 נמוכות ולהישאר פנימי.
Hippo signaling: איך תא יודע שהוא בפנים?
השאלה הבאה היא איך תא פנימי יודע שהוא נמצא בתוך העובר, ולכן צריך לכבות את השעתוק של Cdx2.
כדי שתא פנימי לא יבטא מחדש את Cdx2, חייבת להיות בו מערכת שמזהה את המיקום שלו, ומעכבת את הפעילות של פקטורי השעתוק שמפעילים את Cdx2 (הפקטור TEAD). המערכת המרכזית שמבטלת את מסלול Cdx2 היא Hippo signaling.
ביטוי Cdx2 מופעל דרך פקטורי שעתוק ממשפחת TEAD1-4.
TEAD יכול להיקשר לפרומוטר של Cdx2, אבל כדי להפעיל שעתוק בצורה יעילה הוא צריך שיתוף פעולה עם YAP/TAZ, שהם קו־אקטיבטורים של שעתוק (transcriptions factors, לפי השיעור).
לכן, אם תא רוצה לעצור שעתוק של Cdx2, הוא צריך למנוע מ־YAP/TAZ להגיע לגרעין ולהפעיל את TEAD.
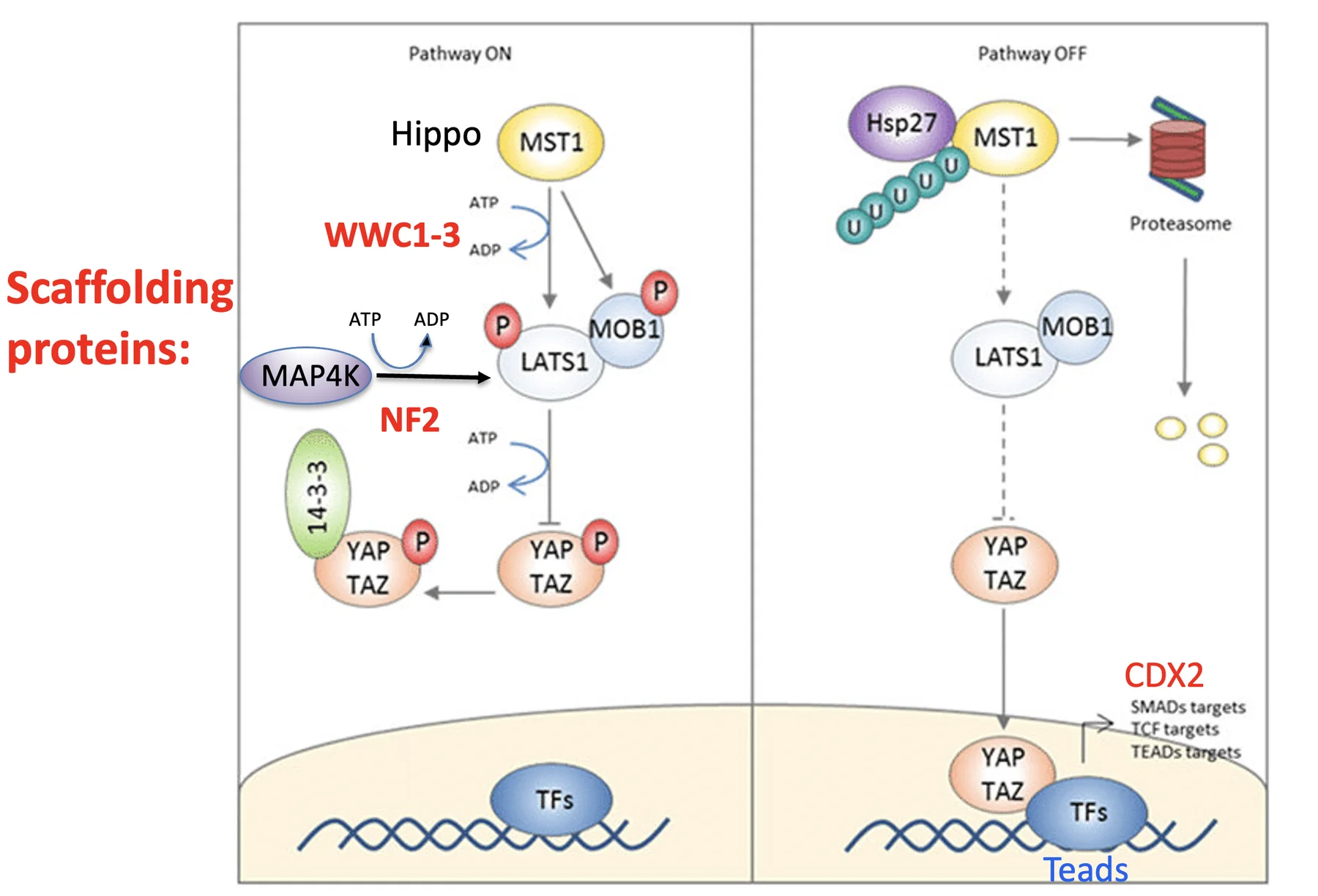
כאשר מסלול Hippo פעיל (ביטול Cdx2):
- LATS1/2 (קינאז) פעיל
- LATS מזרחן את YAP/TAZ
- YAP/TAZ נקשרים לחלבון 14-3-3
- הקישור ל־14-3-3 מקבע את YAP/TAZ בציטופלזמה, ומונע מהם להיכנס לגרעין
- TEAD לא מקבל את הקו־אקטיבציה הדרושה, ולא יכול להפעיל שעתוק של Cdx2
זה מצב שמתאים לתאים פנימיים, שבהם צריך לדכא את הביטוי של Cdx2.
כאשר מסלול Hippo כבוי (הפעלת Cdx2):
- LATS אינו פעיל
- YAP/TAZ אינם מזורחנים בצורה שמחזיקה אותם בציטופלזמה
- YAP/TAZ נכנסים לגרעין
- הם פועלים יחד עם TEAD
- TEAD מפעיל שעתוק של Cdx2
זה מצב שמתאים לתאים חיצוניים, שבהם יש ביטוי גבוה של Cdx2 ומתפתח גורל של TE.
הנקודה המבלבלת במסלול:
- כאשר Hippo פעיל, התוצאה היא דווקא חסימת שעתוק.
- כאשר Hippo כבוי, YAP/TAZ נכנסים לגרעין ומאפשרים שעתוק.
כלומר, "Hippo ON" פירושו עיכוב של YAP/TAZ, ולא הפעלה של שעתוק.
הפעלת LATS במסלול Hippo
השלב המרכזי במסלול הוא הפעלת הקינאז LATS1/2, שכברירת מחדל אינו פעיל ודורש זרחון לשפעול. יש שתי דרכים עיקריות לעשות זאת, ובשתיהן, נדרשים גם חלבוני פיגום (scaffolding proteins) שמקרבים בין הקינאז ו־LATS, וכך מאפשרים את הפעלתו:
-
דרך MST1/2 (נקרא גם Hippo בצורה דרוזופילאית) - דורש חלבון פיגום בשם WWC1 (ראו באותיות אדומות באיור למעלה, מצד שמאל).
-
דרך MAP4K - דורש חלבון פיגום בשם NF2/Merlin (ראו שוב באיור למעלה), חשוב בהקשר של Adherens junctions (ראו להלן).
כשהמסלול כבוי, למשל בעקבות דגרדציית MST1 או היעדר MAP4K, ה־LATS לא מזורחן ואינו פעיל. במצב כזה YAP/TAZ לא מוחזקים בציטופלזמה, נכנסים לגרעין, נקשרים ל־TEAD ומאפשרים שעתוק של Cdx2.
לשלמות התמונה, TEAD/YAP/TAZ לא מפעילים רק את Cdx2, אלא גם גנים נוספים. בהקשר של העובר המוקדם, Cdx2 חשוב במיוחד משום שהוא קשור להתמיינות לכיוון TE.
Amot, NF2 והקשר ל־Adherens Junctions
מסלול Hippo יכול להיות מופעל בדרכים רבות, ובתאים שונים מנגנון ההפעלה יכול להיות שונה. בהקשר של העובר המוקדם, אחת הדרכים החשובות להפעלת Hippo היא Adherens Junctions (שנמצאים בין תאים סמוכים, בצד הבזו־לטרלי של התא, קרי, בתאים פנימיים).
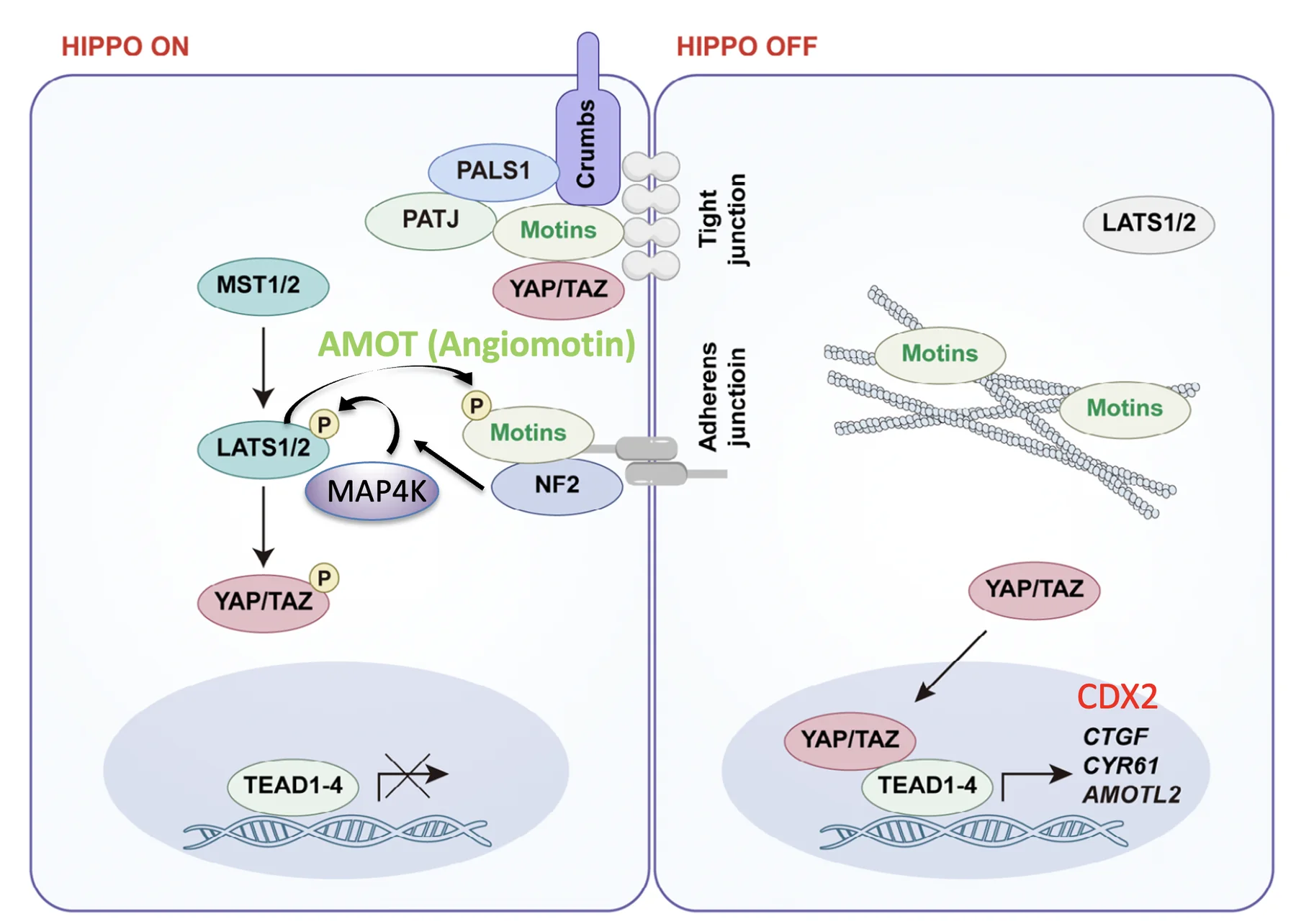
כדי להבין את ההפעלה דרך junctions צריך להבין תחילה את NF2/Merlin.
כאמור, NF2 הוא חלבון פיגום שמאפשר להפגיש את MAP4K ו־LATS, וכך מתאפשר זרחון ושפעול של LATS. אבל NF2 לא תמיד פעיל; הוא נמצא באזורי ה־junctions ויכול להיות במצב auto-inhibited (מעוכב ולא מפעיל את המסלול).
כדי להפעיל את NF2, דרושה משפחת חלבונים בשם Motins. החלבון המרכזי בה, בהקשר של העובר, הוא Amot (Angiomotin).
Amot נמצא בשיווי משקל בין שני מצבים:
- קשור ל־F-actin (שלהבנתי מרוכז באזור האפיקלי), ופחות זמין להפעלת NF2
- חופשי יותר ומשתתף בהפעלת Hippo באזורי ה־junctions
כאשר Amot משתתף בגיוס ובהפעלה של NF2 באזורי ה־Adherens Junctions, NF2 יכול לקרב את MAP4K ו־LATS. כך MAP4K מזרחן את LATS, LATS מופעל, ואז מזרחן את YAP/TAZ ומונע מהם להיכנס לגרעין (ולכן TEAD אינו פעיל, ואין שעתוק של Cdx2).
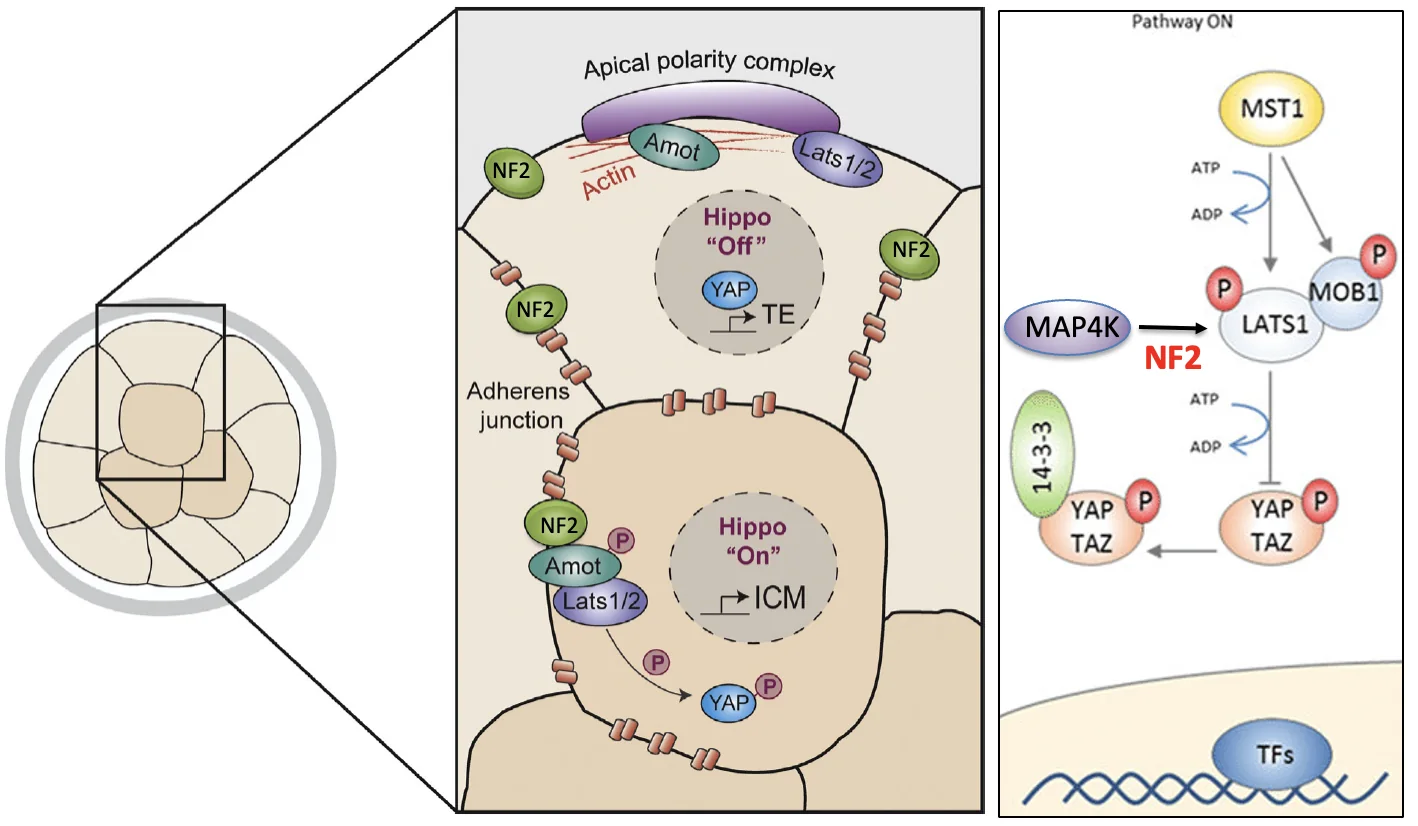
בתאים פנימיים יש הרבה מגעים בין תאים ו־Adherens Junctions. ה־junctions מאפשרים הפעלה של Amot, NF2 ו־LATS, ולכן Hippo פעיל. התוצאה היא ש־YAP/TAZ נשארים בציטופלזמה, TEAD לא מפעיל שעתוק של Cdx2, והתא מתקדם לכיוון ICM.
בתאים חיצוניים יש פולריות אפיקלית ומבנה שונה של מגע בין תאים. Hippo נשאר כבוי, YAP/TAZ כן נכנסים לגרעין, TEAD מפעיל שעתוק Cdx2, והתא מתקדם לכיוון TE.
ההבדל בין תאים פנימיים וחיצוניים:
| תא | מצב Hippo | תוצאה |
|---|---|---|
| פנימי | Hippo פעיל | YAP לא נכנס לגרעין ← אין TEAD פעיל ← אין שעתוק Cdx2 |
| חיצוני | Hippo כבוי | YAP נכנס לגרעין ← TEAD פעיל ← יש שעתוק Cdx2 |
בתאים פנימיים יש הרבה מגעים בין תאים ו־Adherens junctions, ולכן יש תנאים להפעלת Hippo. בתאים חיצוניים, הקומפלקס האפיקלי קושר רכיבים כמו Amot ו־LATS והופך אותם לפחות זמינים להפעלת Hippo. לכן, בתאים החיצוניים Hippo כבוי, ו־Cdx2 יכול להתבטא.
חיבור המודל: Cdx2, פולריות ו־Hippo
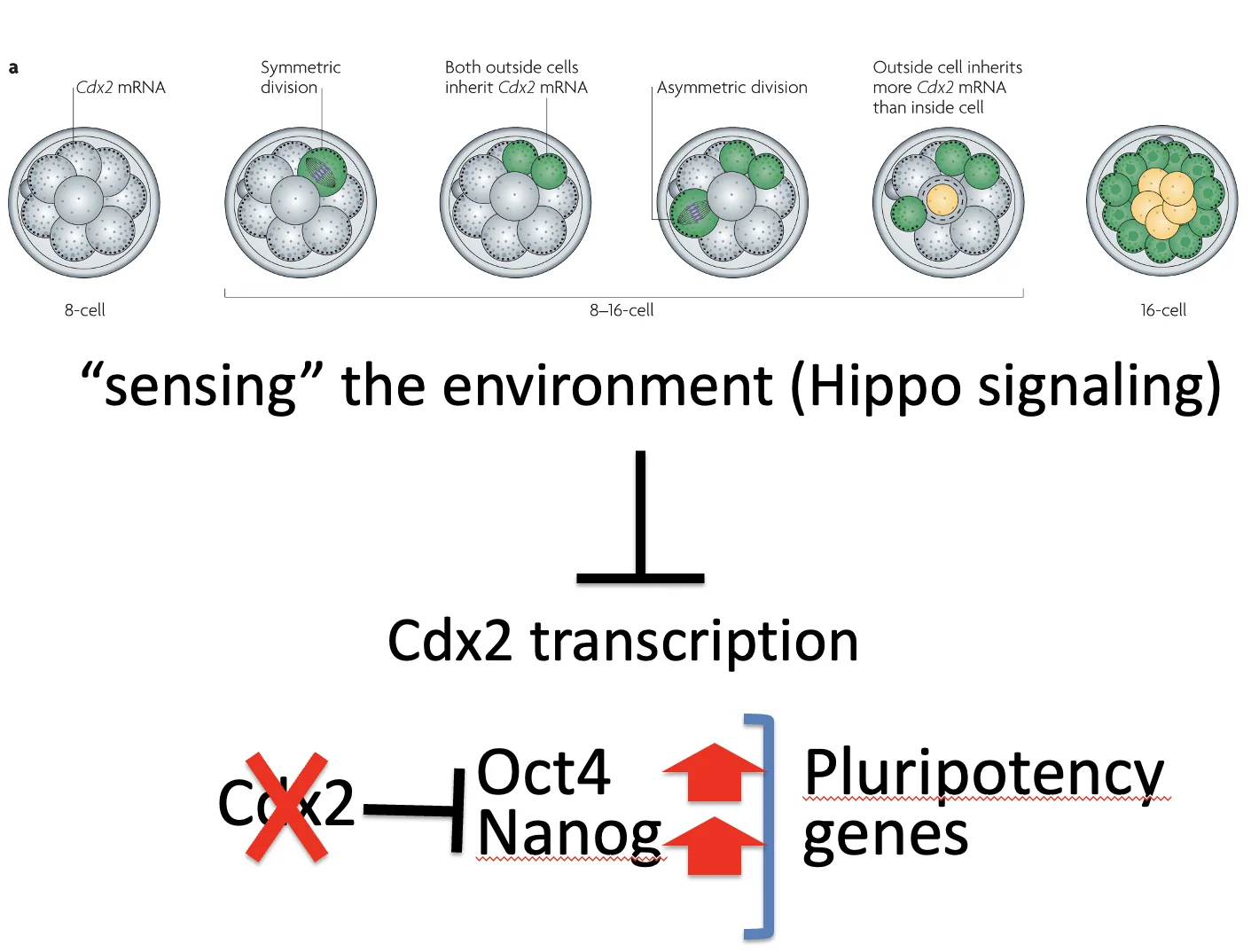
המודל הכללי של First cell fate decision:
- בעובר בן שמונה תאים יש פולריות, ולוקליזציה אפיקלית של Cdx2 mRNA
- בחלוקה סימטרית, שני תאי־הבת נשארים חיצוניים, פולריים, וממשיכים לבטא Cdx2
- בחלוקה א-סימטרית, תא אחד נכנס פנימה ומקבל מעט Cdx2 mRNA
- בתא הפנימי מופעל Hippo
- Hippo מונע כניסת YAP/TAZ לגרעין
- TEAD אינו מפעיל שעתוק של Cdx2
- רמות Cdx2 נשארות נמוכות
- Oct4 ו־Nanog יכולים להתבטא, והתא נשמר בכיוון ICM
Cdx2 ומישור החלוקה
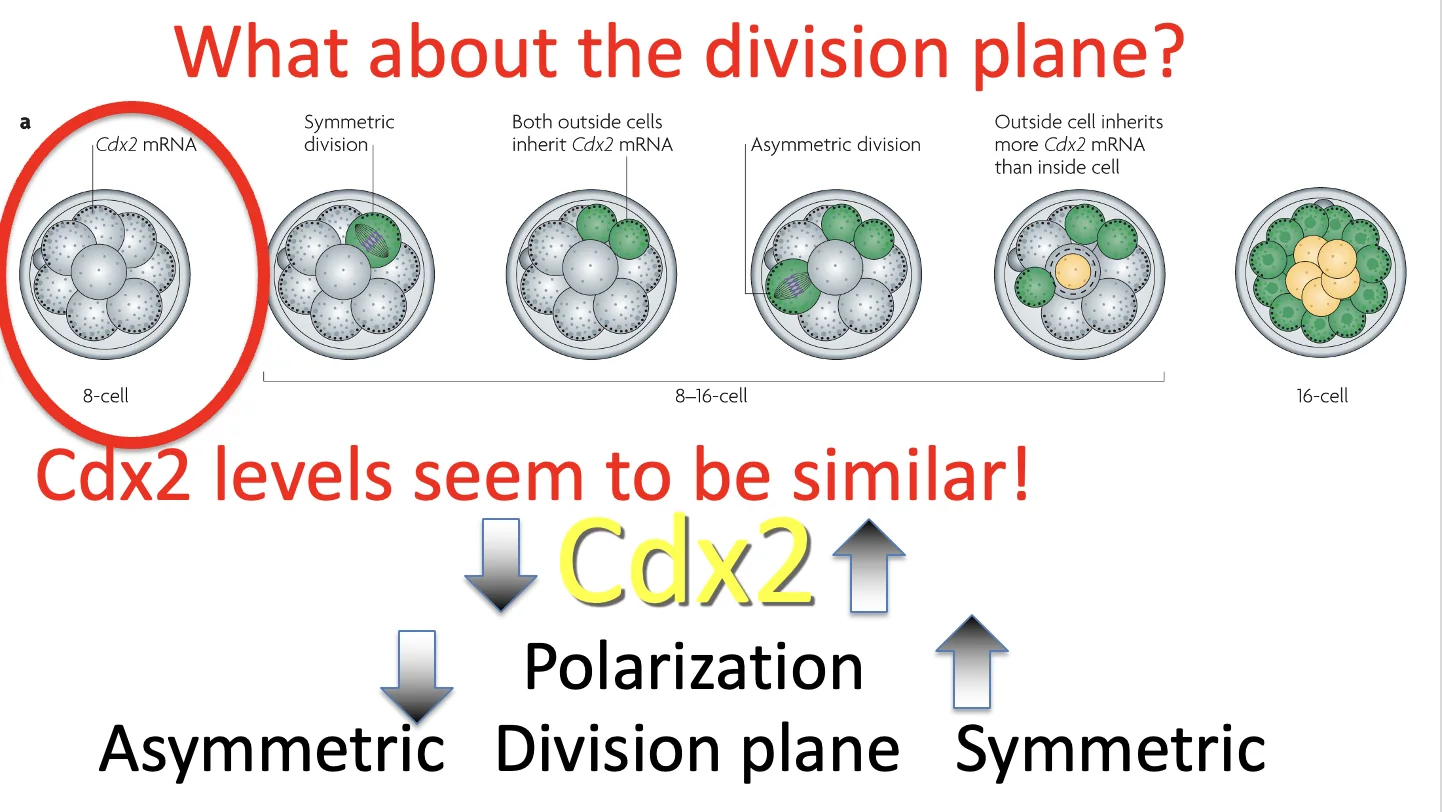
יש גם קשר בין רמות Cdx2 לבין סוג החלוקה:
- רמות גבוהות יותר של Cdx2 ← סיכוי גבוה יותר לחלוקה סימטרית (שני תאי בת פולריים). מפני שכאמור, Cdx2 מחזק את הפולריות, ולכן תא עם הרבה Cdx2 נוטה להישאר חיצוני (פולרי).
- רמות נמוכות יותר של Cdx2 ← סיכוי גבוה יותר לחלוקה א-סימטרית (תא בת אחד פולרי, תא בת שני לא פולרי)
חשוב: מדובר בסיכויים, לא בכלל מוחלט.
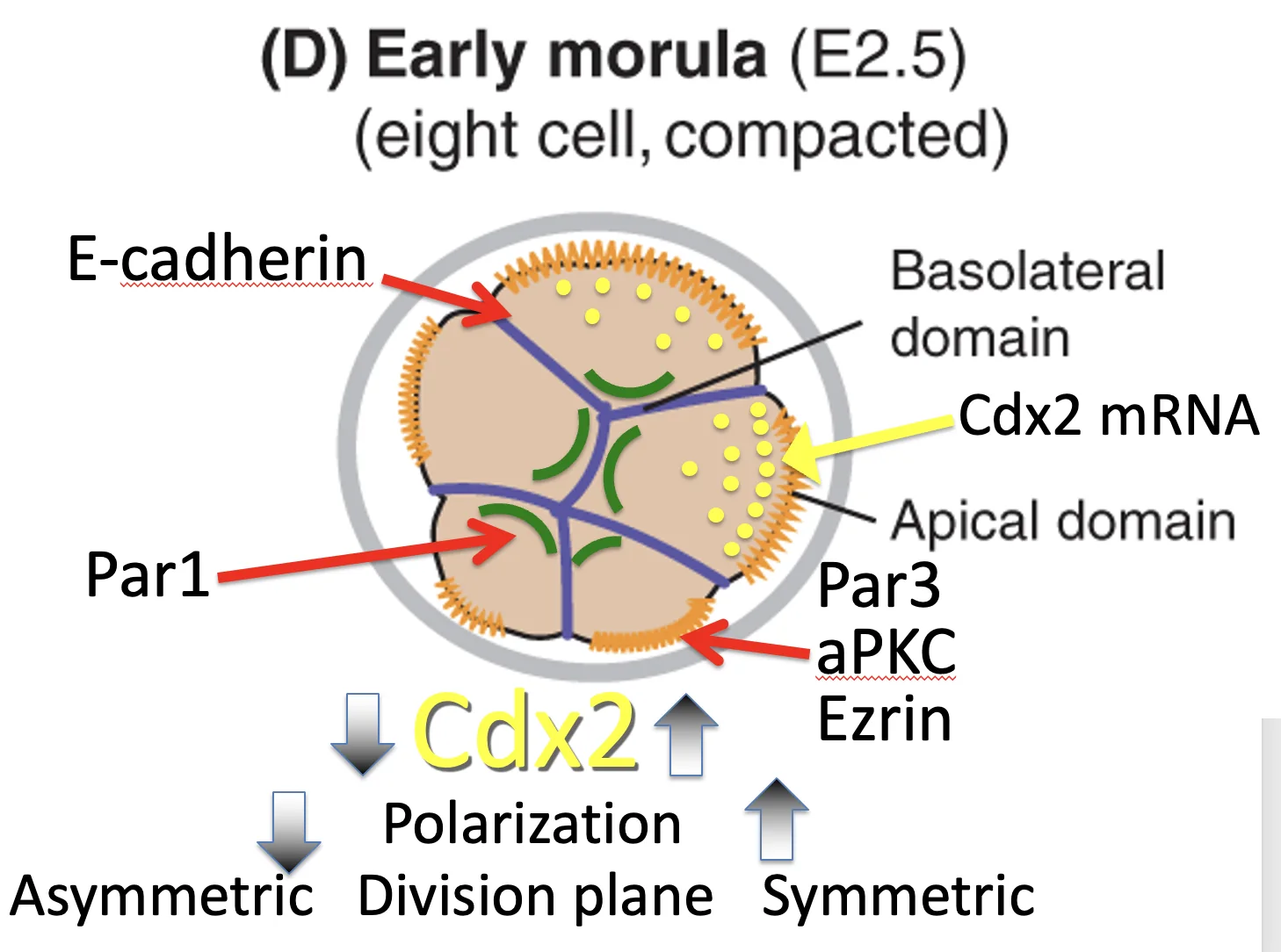
הסכמה בתמונה למעלה, שבה כל התאים בשלב שמונת התאים מבטאים אותה כמות של Cdx2, אינה מדויקת. לפי השיעור, כבר בשלבים האלה יש בין תאים שונות ברמות Cdx2, והשונות יכולה להשפיע על מישור החלוקה ועל יצירת תאים פנימיים.
Cdx2 בעיקר מוביל ל־TE, ומשפיע על כל התהליכים הבאים:
- פולריות
- מישור החלוקה
- first cell fate decision
- compaction
בדיקה עצמית: הסבר איך Cdx2 משפיע על כל אחד מהתהליכים.
תשובה
- פולריות: Cdx2 מחזק את הפולריות האפיקלית, יותר Cdx2 ← יותר פולרי.
- מישור החלוקה: תא עם יותר Cdx2 נוטה לחלוקה סימטרית, תא עם פחות Cdx2 נוטה לחלוקה א-סימטרית.
- First cell fate decision: תא פנימי עם פחות Cdx2 יפעיל Hippo, TEAD לא יפעיל Cdx2, ולכן התא יישאר בכיוון ICM. תא חיצוני עם יותר Cdx2 יבטא Cdx2 ויתקדם לכיוון TE.
- compaction: מוזמנים להשלים לבד, וייתכן שגם cavitation.
Zygotic Genome Activation וגלי כניסה ל־ICM
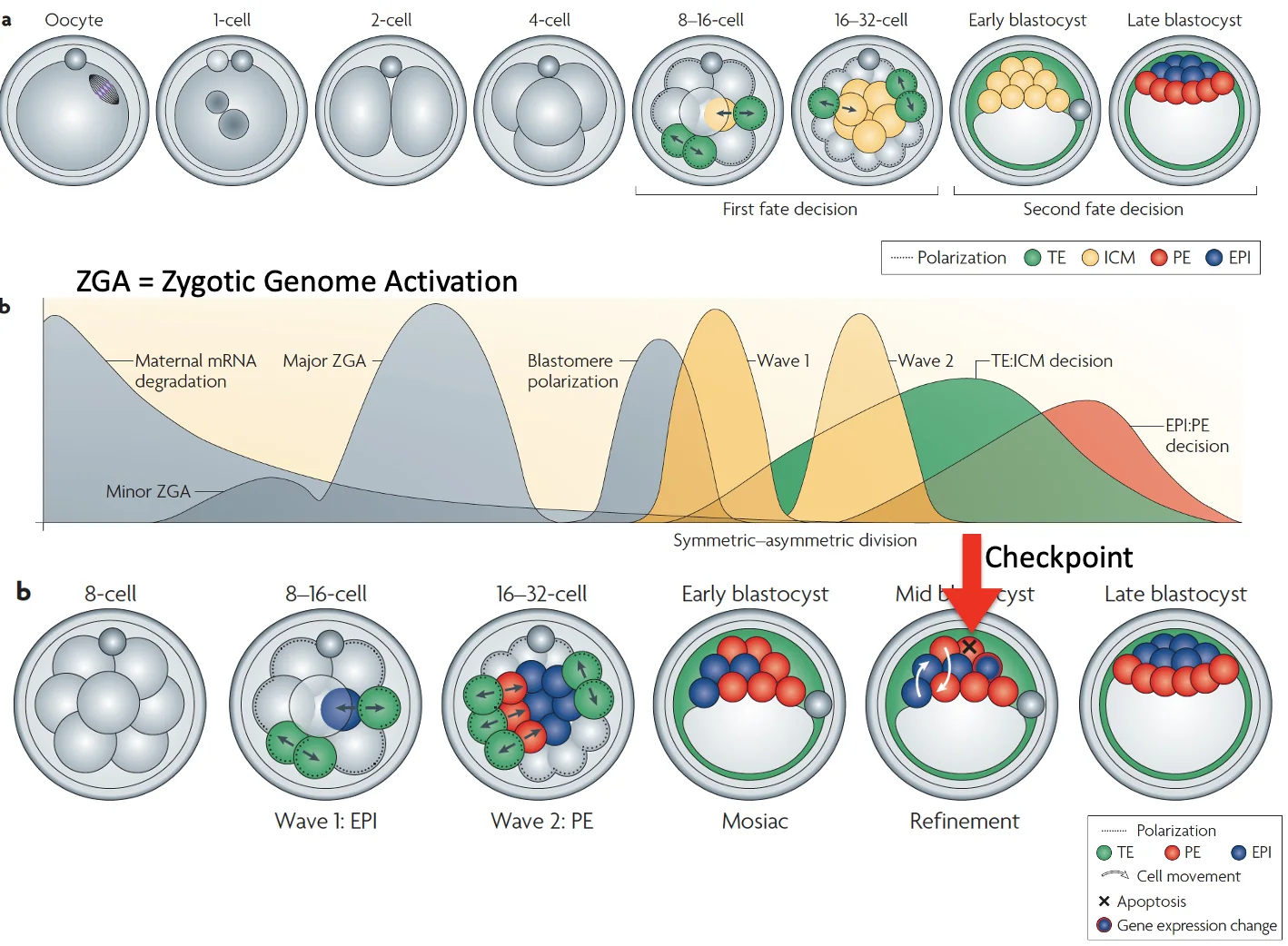
בתחילת ההתפתחות, עד לשלב שני התאים, הגנום העוברי מושתק, וה־RNA שמניע את השלבים הראשונים מגיע מהביצית. בשלב שני התאים מתרחשת Zygotic Genome Activation (ZGA), כלומר התחלת הפעילות של הגנום העוברי.
נקודה חשובה: תאי ה־ICM לא נוצרים בבת אחת; יש שני גלים של חלוקות א-סימטריות:
| גל | מה קורה? | גורל מועדף |
|---|---|---|
| 1 | תאים נכנסים מוקדם יותר פנימה | לרוב Epiblast |
| 2 | תאים נכנסים מאוחר יותר פנימה | לרוב Primitive endoderm |
לאחר מכן מתרחש sorting: תאים שאמורים להיות Primitive endoderm מסתדרים באזור המתאים, ותאים שאמורים להיות Epiblast נשארים באזור אחר.
אם תא מבטא תוכנית שאינה מתאימה למיקום שלו, למשל, תא שמבטא Gata4 (שמוביל ל־Primitive endoderm) אך נשאר במקום לא מתאים, העובר יכול להפעיל תהליך אפופטוזיס. זאת מעין נקודת ביקורת חשובה לפני יצירת האפיבלסט, משום שהאפיבלסט ייתן בהמשך את העובר כולו.
Positive feedback loops ברשת פקטורי השעתוק
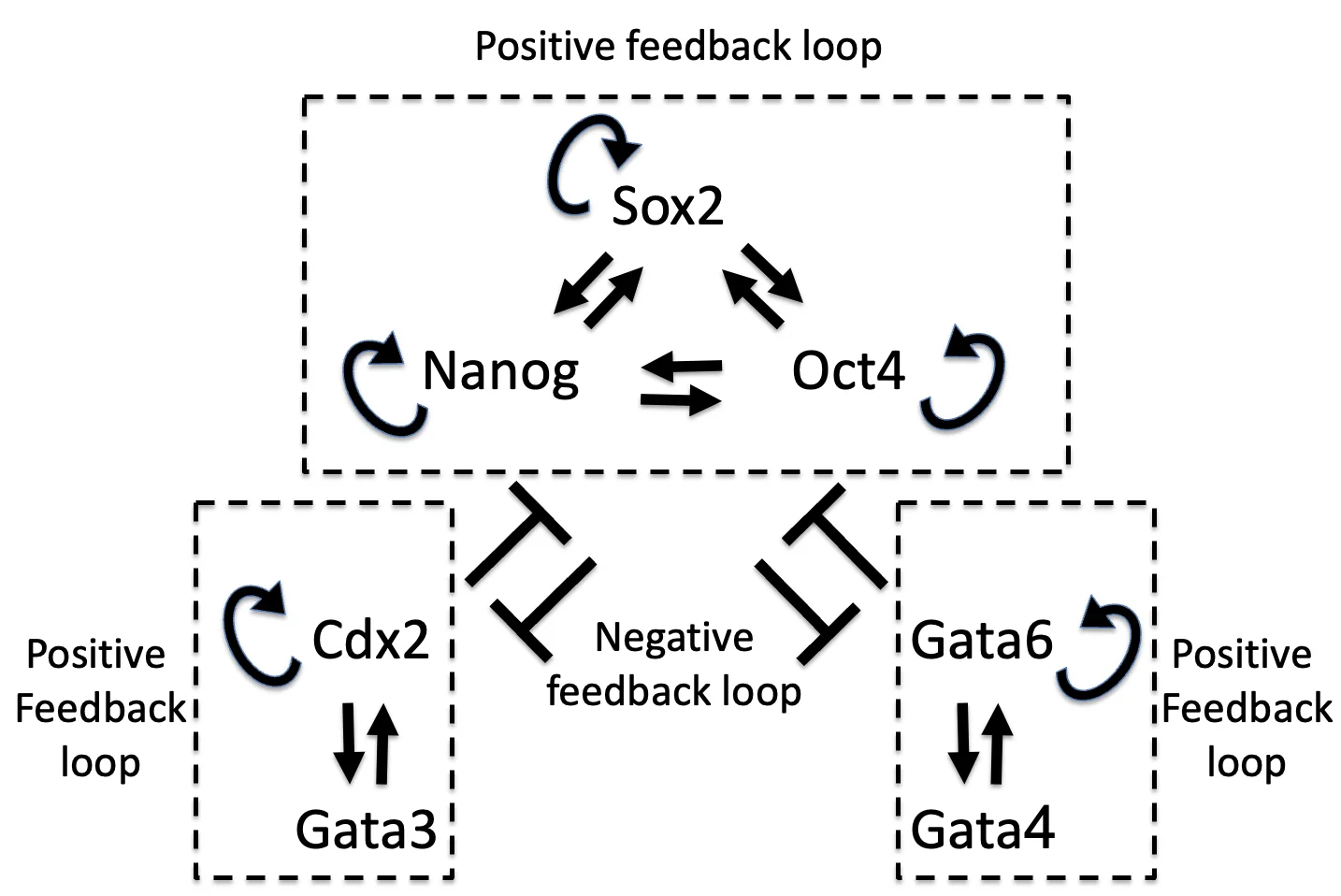
בתמונה למעלה מודל של רשת פקטורי שעתוק. המודל לא נדרש ברמת המשוואות, אלא ברמת ההיגיון: שינוי קטן ברמות של פקטור מסוים יכול לעבור הגברה דרך positive feedback loop.
שלוש קבוצות מרכזיות:
| קבוצה | פקטורים | משמעות |
|---|---|---|
| Pluripotency maintaining genes | Oct4, Nanog, Sox2 | שימור כיוון פלוריפוטנטי / Epiblast |
| TE genes | Cdx2, Gata3 | קידום גורל TE |
| Primitive endoderm genes | Gata6, Gata4 | קידום גורל Primitive endoderm |
בתוך כל קבוצה יש קשרי הגברה חיוביים. בין קבוצות שונות יש קשרי דיכוי. לכן, כשקבוצה אחת מתחילה לקבל יתרון, היא יכולה להגביר את עצמה ולדכא את האחרות.
לדוגמה:
- אם Nanog עולה, הוא יכול לחזק את הרשת של הפלוריפוטנטיות (Oct4/Nanog/Sox2) ולדכא את Cdx2 ו־Gata6.
- אם Cdx2 דומיננטי, הוא מחזק את כיוון ה־TE ומדכא את Oct4/Nanog.
- אם Gata6 עולה, הוא מקדם את כיוון ה־Primitive endoderm ומדכא את הרשת הפלוריפוטנטית.
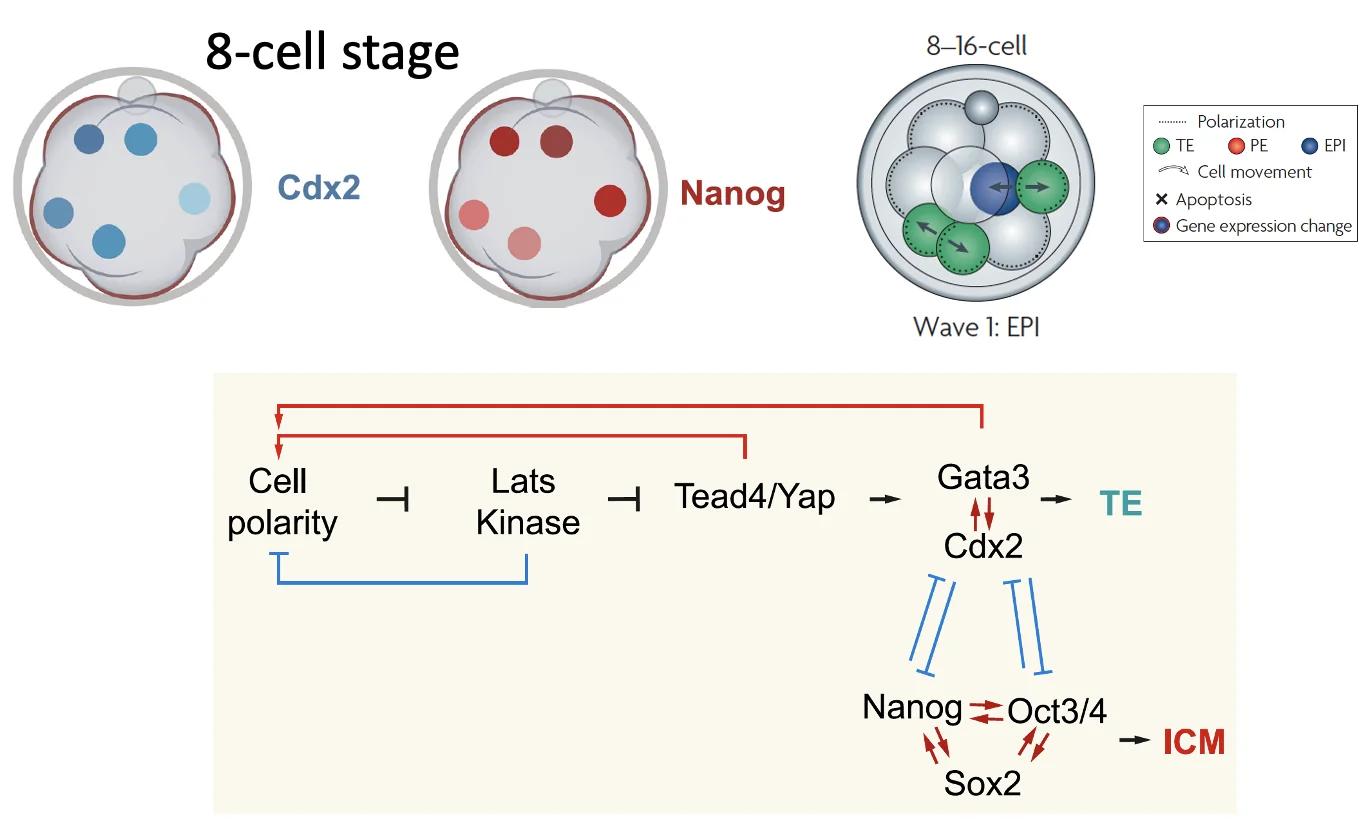
במודל של First cell fate decision, חלוקה א-סימטרית יוצרת חלון זמן שבו תא פנימי מקבל פחות Cdx2. אם בתא הזה יש מעט Nanog, ו־Cdx2 נשאר נמוך בגלל Hippo, רשת הפלוריפוטנטיות יכולה לעבור הגברה ולהשתלט. כך התא נשאר בכיוון ICM.
Second cell fate decision ו־FGF signaling
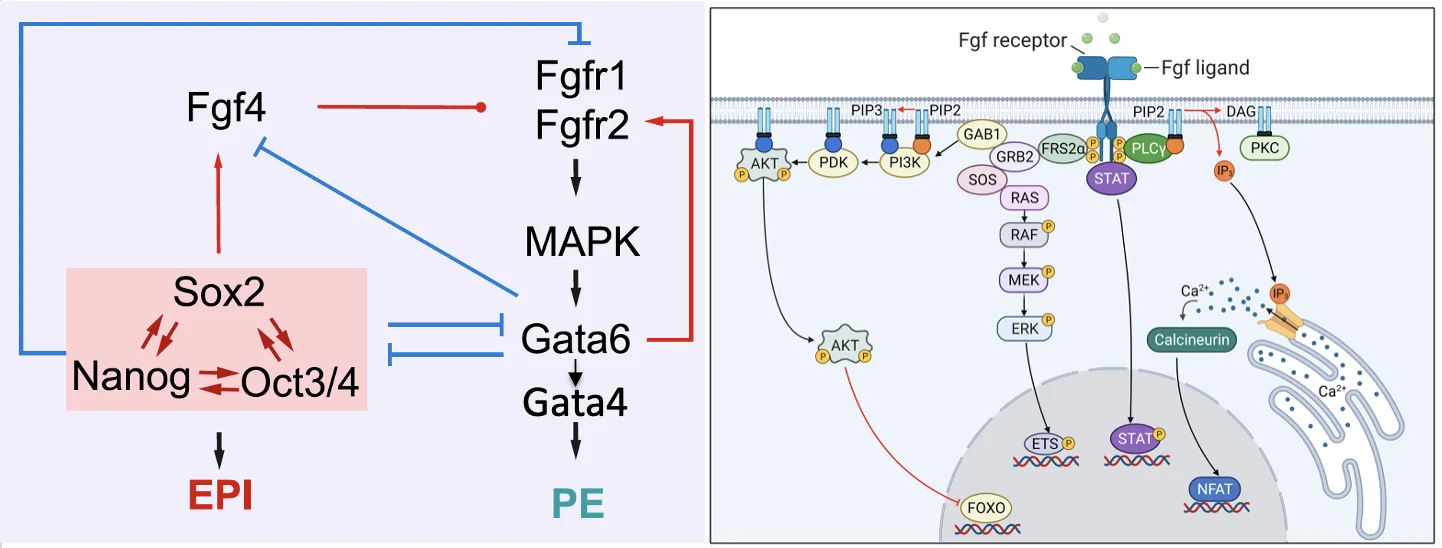
בשלב ה־Second cell fate decision, תאי ה־ICM מתפצלים לשני כיוונים:
| אוכלוסייה | פקטורים מרכזיים | גורל |
|---|---|---|
| תאים עם Nanog / Oct4 / Sox2 | Pluripotency maintaining genes | Epiblast |
| תאים עם Gata6 / Gata4 | Primitive endoderm genes | Primitive endoderm |
כאן נכנס FGF signaling.
- תא שמבטא את קבוצת הפלוריפוטנטיות (Oct4/Nanog/Sox2) מבטא את הליגנד במסלול FGF (ה־FGF4).
- אותו תא, מדכא את ביטוי הרצפטורים של המסלול (FGFR1/FGFR2), כך שבסופו של דבר, הוא בעיקר מייצר את הליגנד ולא מגיב לו (אין לו רצפטורים לליגנד).
- תא שהולך לכיוון Gata6 מבטא את הרצפטורים ל־FGF (ה־FGFR1/FGFR2), אבל אינו מבטא את הליגנד FGF4.
- כאשר הליגנד FGF4 נקשר לרצפטור בתא מתאים, מופעל מסלול שמעלה עוד יותר את Gata6, ובהמשך את Gata4, ומקדם Primitive endoderm.
הערה: להבנתי, זה דומה למסלול שתואר בשיעור הקודם ביחס לפעילות של Nanog אל מול Gata4.
הגל הראשון
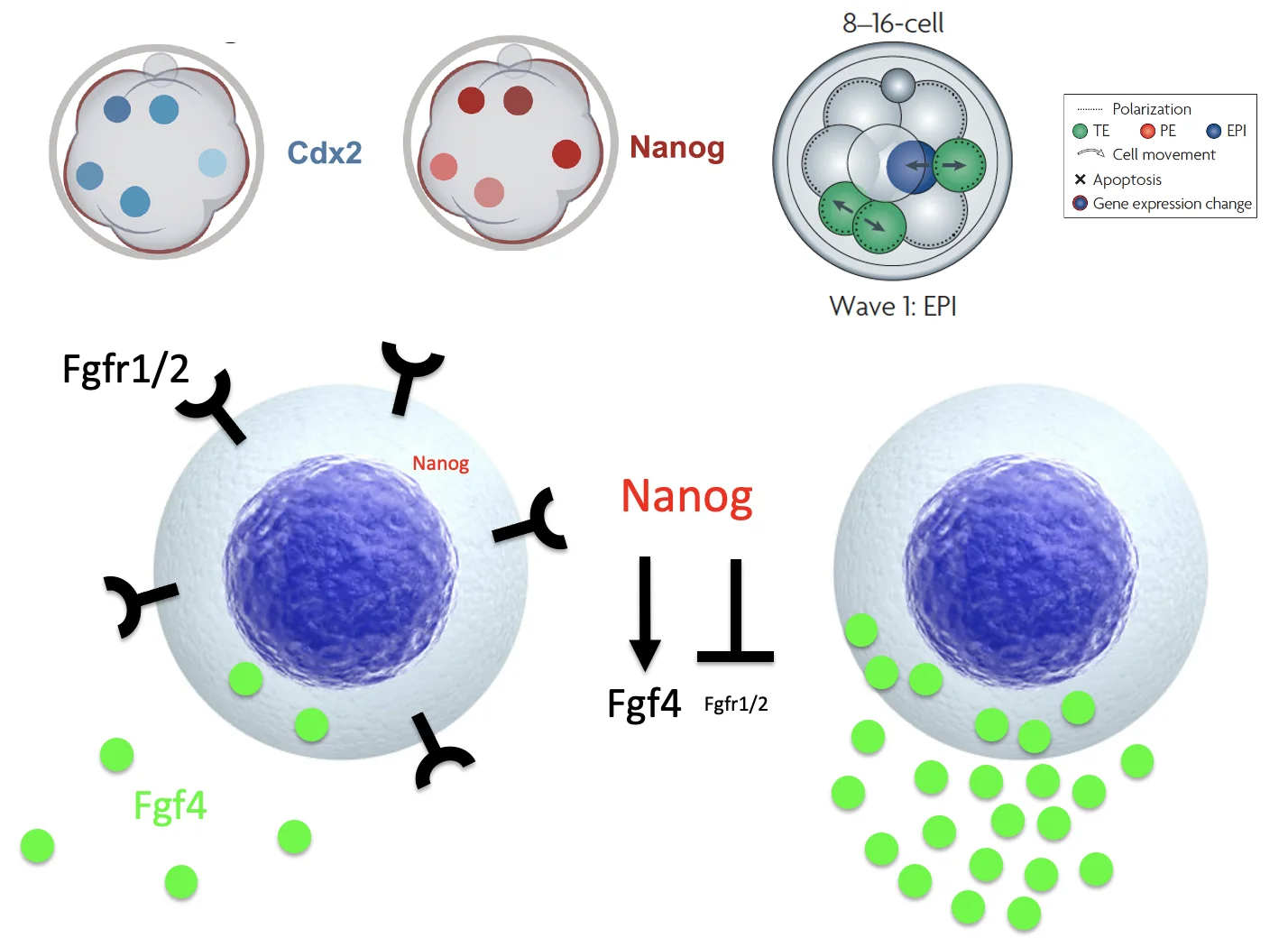
בגל הראשון, התאים שנכנסים פנימה מעלים Nanog. בעקבות זאת הם מבטאים יותר FGF4 ומדכאים את הרצפטורים ל־FGF. לכן הם בעיקר מקור לליגנד, ונוטים להישאר בכיוון Epiblast.
הגל השני
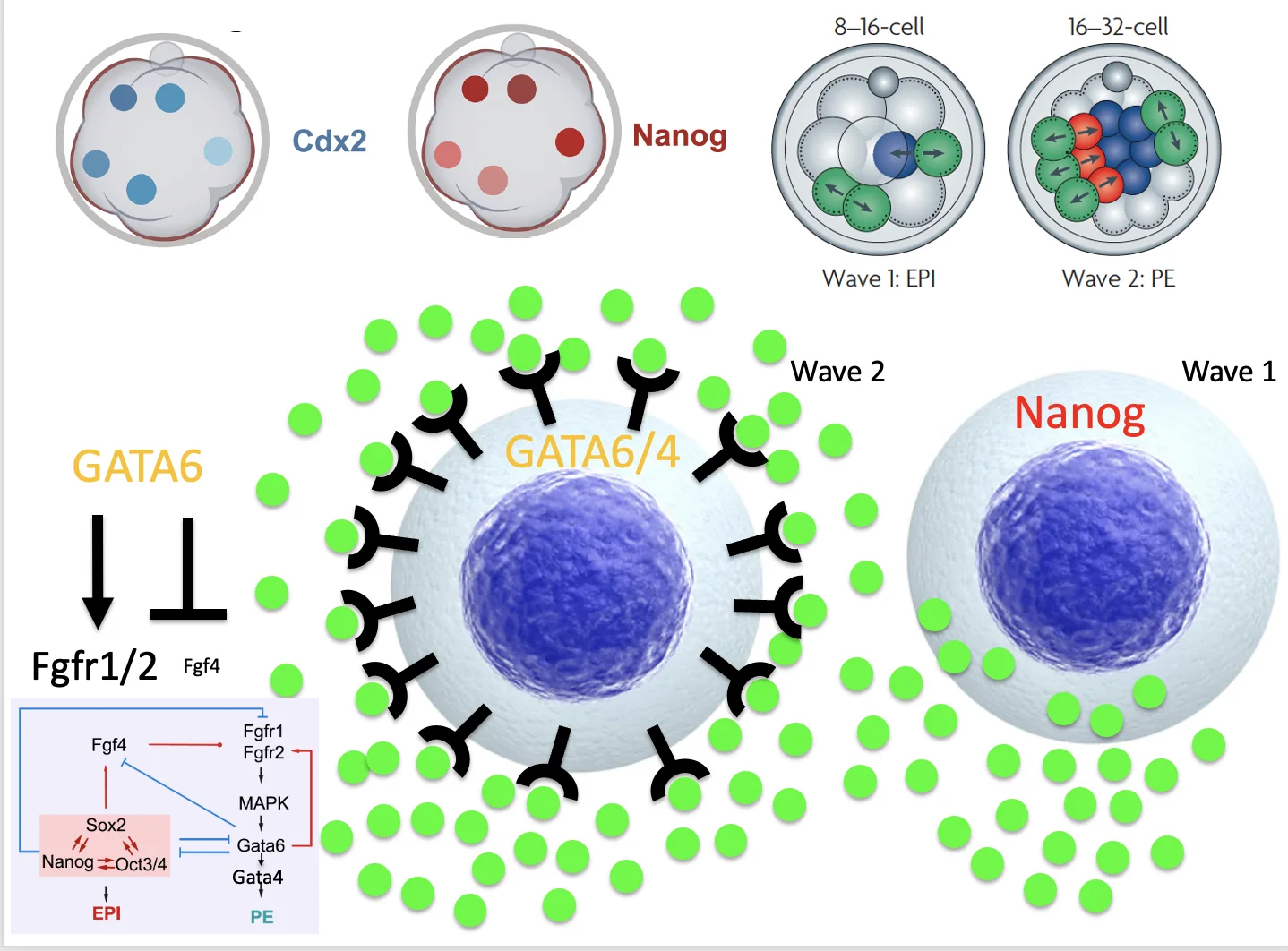
בגל השני, תאים שנכנסים פנימה מגיעים לסביבה שבה כבר יש הרבה FGF4 שהופרש מהתאים של הגל הראשון. גם אם הרצפטורים שלהם בתחילה ברמה נמוכה, כמות הליגנד בסביבה יכולה להפעיל FGF signaling. בעקבות זאת עולה Gata6, שמעלה את רמות הרצפטורים ומדכא את רשת Nanog/Oct4/Sox2. כך התאים נוטים לכיוון Primitive endoderm.
זה ההסבר לכך שתאי Wave 1 נוטים להפוך ל־Epiblast, בעוד שתאי Wave 2 נוטים להפוך ל־Primitive endoderm.
רצף האירועים המרכזי
Zygote
↓
2-cell embryo
↓
Zygotic Genome Activation
↓
4-cell embryo
↓
8-cell embryo
↓
Compaction + Polarization
↓
First cell fate decision
├─ Outer cells
│ ↓
│ Hippo OFF
│ ↓
│ YAP/TAZ + TEAD active
│ ↓
│ Cdx2 ON
│ ↓
│ Trophectoderm
│
└─ Inner cells
↓
Hippo ON
↓
YAP/TAZ kept out of nucleus
↓
Cdx2 OFF
↓
Oct4/Nanog/Sox2 allowed
↓
ICM
↓
Second cell fate decision
├─ Nanog / Oct4 / Sox2
│ ↓
│ FGF4 ligand
│ ↓
│ Epiblast
│
└─ FGF response → Gata6 / Gata4
↓
Primitive endoderm
משפטי מפתח
- Cdx2 חיוני לשימור זהות ה־TE וליצירת מחסום אפיתליאלי יציב.
- ללא Cdx2, ה־Blastocoel יכול להתחיל להיווצר, אבל אינו נשמר.
- Tight junctions נמצאים בתאים החיצוניים וחשובים לשימור ה־Blastocoel.
- E-cadherin דרוש ל־Compaction.
- פולריות ומישור חלוקה הם שני מנגנונים מרכזיים ביצירת inner cells ו־outer cells.
- Hippo פעיל בתאים פנימיים ומונע שעתוק של Cdx2.
- Hippo כבוי בתאים חיצוניים ומאפשר שעתוק של Cdx2.
- Cdx2 מדכא את Oct4 ו־Nanog בתאי ה־TE.
- בתאי ICM, הורדת Cdx2 מאפשרת ל־Oct4/Nanog/Sox2 לשמר פלוריפוטנטיות.
- ב־Second cell fate decision, ה־FGF4 שמופרש מתאים בכיוון Epiblast משפיע על תאים אחרים ומקדם בהם Gata6/Gata4 ו־Primitive endoderm.