פתיחה - מה קורה לגזים לאחר פעפוע
בשיעורים הקודמים דיברנו על המכניקה של הנשימה, על הרכב הגזים בנאדיות, ועל פעפוע גזים בין הנאדיות לדם. מכאן עוברים לשאלה הבאה: אחרי שגזים עברו לתוך הדם או יצאו ממנו, איך הגוף משנע אותם?
שני הגזים המרכזיים מתנהגים באופן שונה:
- $\ce{CO2}$ (פד״ח) מסיס מאוד, ולכן חלק גדול ממנו יכול לעבור עיבוד כימי בתוך הדם.
- $\ce{O2}$ (חמצן) פחות מסיס, ולכן הגוף חייב להשתמש בהמוגלובין כדי לשנע כמות משמעותית שלו.
מדובר במערכת דינמית: גזים מפעפעים לפי מפלי לחצים, נקשרים ומתנתקים מהמוגלובין, עוברים תגובות דו־כיווניות, ומושפעים מהתנאים ברקמות ובריאות.
שינוע פחמן דו־חמצני בדם
פחמן דו־חמצני, $\ce{CO2}$, מיוצר ברקמות כתוצר לוואי (פסולת) של מטבוליזם. הוא עובר מהרקמות לנימים, ומשם ממשיך לפלזמה ולכדוריות הדם האדומות.
בדם, $\ce{CO2}$ משונע בשלוש צורות עיקריות:
| צורת שינוע | חלק יחסי | מה קורה |
|---|---|---|
| $\ce{CO2}$ מומס | כ־7% | נשאר מומס בפלזמה או בציטופלזמה של הכדורית האדומה |
| קרבאמינו־המוגלובין (Carbaminohemoglobin) | כ־23% | $\ce{CO2}$ נקשר לקבוצות אמיניות בהמוגלובין הלא־מחומצן |
| ביקרבונט $\left(\ce{HCO3^-}\right)$ | כ־70% | $\ce{CO2}$ הופך ל־$\ce{HCO3^-}$ בתוך הכדורית האדומה |
למרות של־$\ce{CO2}$ יש מסיסות גבוהה, רק חלק קטן ממנו נשאר כ־$\ce{CO2}$ מומס (כ־7%); רובו הופך לביקרבונט או נקשר להמוגלובין.
ברקמות, $\mathrm{P_{\ce{CO2}}}$ בדם עולה מבערך $40\, \mathrm{mmHg}$ ל־$45\text{–}48\, \mathrm{mmHg}$. לכאורה עלייה קטנה, אך בפועל, בגלל המסיסות הגבוהה של $\ce{CO2}$, שינוי קטן בלחץ החלקי יכול לייצג כמות גדולה יחסית של מולקולות.
התגובה הביוכימית של $\ce{CO2}$ ומים
בתוך כדוריות הדם האדומות נמצא האנזים Carbonic Anhydrase, שמאיץ תגובה דו־כיוונית בין $\ce{CO2}$ ומים לבין חומצה פחמתית:
\[\ce{CO2 + H2O <=>[Carbonic\ Anhydrase] H2CO3 <=> H^+ + HCO3^-}\]השלבים הם:
- $\ce{CO2}$ נכנס לכדורית הדם האדומה
- $\ce{CO2}$ מתחבר למים
- Carbonic Anhydrase מאיץ יצירת $\ce{H2CO3}$, חומצה פחמתית (carbonic acid)
- החומצה הפחמתית מתפרקת ל־$\ce{H^+}$ ול־$\left(\ce{HCO3^-}\right)$
- הפרוטון, $\ce{H^+}$, נקשר להמוגלובין
- הביקרבונט, $\ce{HCO3^-}$, יוצא מהכדורית האדומה לפלזמה
- כדי לשמור על איזון מטענים, יון כלור, $\ce{Cl^-}$, נכנס לכדורית האדומה. זהו Chloride Shift
התגובה מסבירה למה עלייה ב־$\ce{CO2}$ יכולה להוריד pH: גם אם לביקרבונט יש אופי בסיסי, התוצר הסופי של יצירת חומצה פחמתית ופירוקה כולל $\ce{H^+}$, ולכן הכיוון הכללי הוא חומצי.
הקשר הזה עומד מאחורי חמצת נשימתית: כאשר $\ce{CO2}$ מצטבר, חלק גדול יותר ממנו יכול להפוך לחומצה פחמתית ולתרום לירידת pH.
אותו עיקרון מוכר גם ממשקה מוגז: $\ce{CO2}$ מומס במים, חלקו הופך לחומצה פחמתית, ולכן המשקה חומצי יותר ממים רגילים.
המצב ברקמות ובריאות
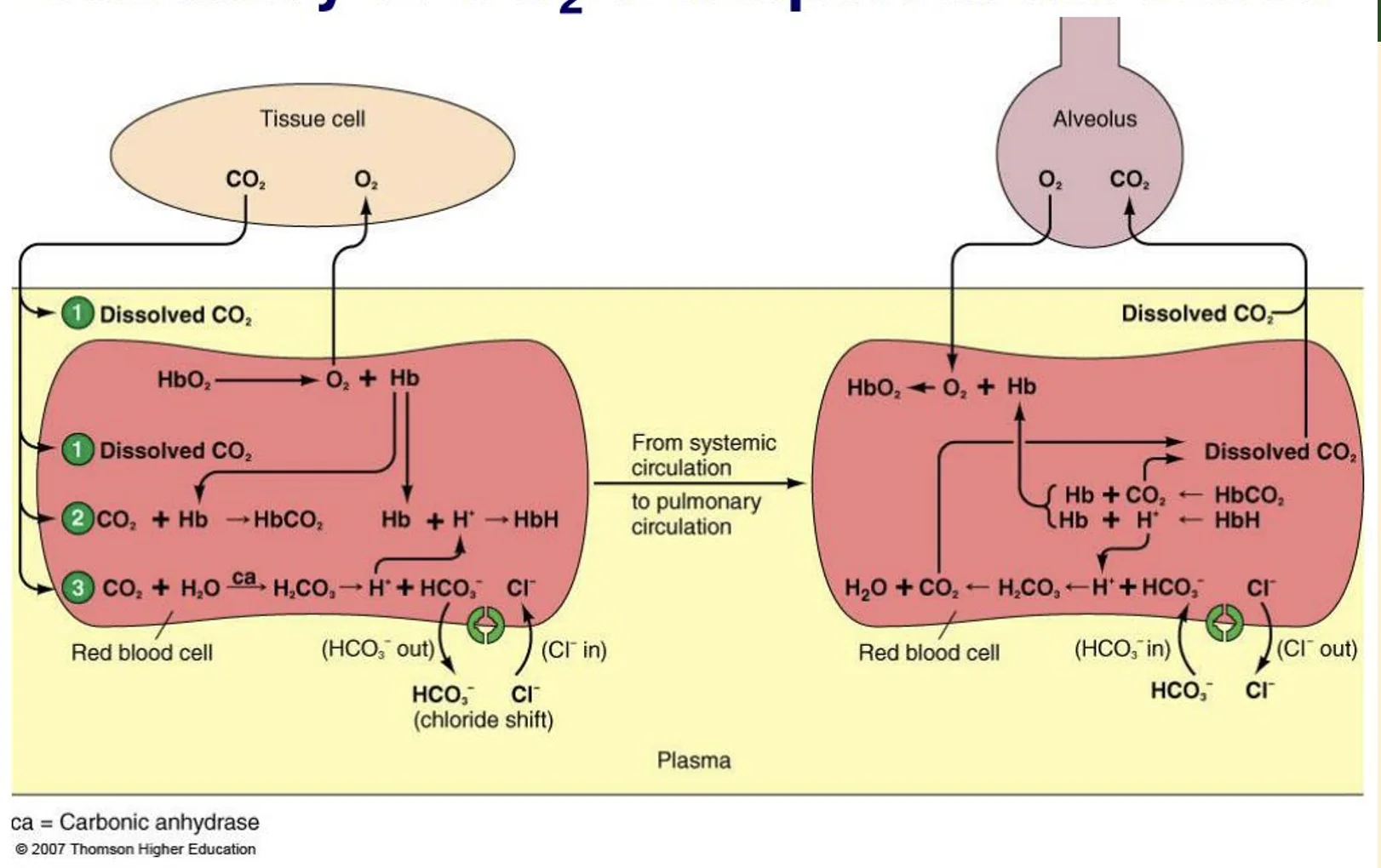
ברקמות
ברקמות יש פעילות מטבולית. לכן התנאים שם שונים מהתנאים בריאות:
- רמת $\ce{CO2}$ גבוהה יותר
- רמת $\ce{O2}$ נמוכה יותר
- הטמפרטורה מעט גבוהה יותר בגלל הפעילות המטבולית
- ה־pH נוטה לרדת בגלל יצירת $\ce{H^+}$ מתהליך $\ce{CO2}$-ביקרבונט
בתנאים האלה $\ce{CO2}$ מפעפע מהרקמות אל הפלזמה, ומשם לתוך הכדוריות האדומות.
בתוך כדורית הדם האדומה יש ל־$\ce{CO2}$ שלושה מסלולים (ראו בצד השמאלי של התמונה למעלה):
- להישאר מומס בציטופלזמה
- להיקשר להמוגלובין הלא־מחומצן וליצור Carbaminohemoglobin
- להפוך לביקרבונט דרך Carbonic Anhydrase
הפרוטון שנוצר בתגובה נקשר להמוגלובין, ובכך ההמוגלובין ממתן חלק מההשפעה החומצית. הביקרבונט יוצא לפלזמה דרך Chloride Shift.
בריאות
בריאות מתרחש אותו תהליך בכיוון ההפוך.
הדם מגיע לנימי הריאה, ובנאדיות יש ריכוז חמצן גבוה יחסית. ההמוגלובין נקשר לחמצן. כאשר ההמוגלובין נקשר לחמצן, הוא פחות “אוהב” להיות קשור ל־$\ce{CO2}$ ול־$\ce{H^+}$, ולכן הוא משחרר אותם.
במקביל:
- $\ce{CO2}$ מומס בפלזמה מפעפע אל הנאדיות
- ירידת $\ce{CO2}$ בפלזמה יוצרת מפל שמושך $\ce{CO2}$ מתוך הכדורית האדומה אל הפלזמה
- ביקרבונט נכנס חזרה מהפלזמה אל הכדורית האדומה
- כלור יוצא מהכדורית האדומה
- $\ce{HCO3^-}$ מתחבר ל־$\ce{H^+}$ ויוצר שוב $\ce{H2CO3}$
- Carbonic Anhydrase הופך $\ce{H2CO3}$ ל־$\ce{CO2}$ ולמים
- $\ce{CO2}$ מפעפע החוצה אל הפלזמה ומשם אל הנאדית
מכיוון שכל התגובות האלה דו־כיווניות, הכיוון בפועל נקבע לפי מפלי הריכוזים והלחצים. ברקמות, $\ce{CO2}$ נכנס לדם וחמצן משתחרר. בריאות, חמצן נכנס לדם ו־$\ce{CO2}$ משתחרר אל הנאדיות.
שינוע חמצן והמוגלובין
חמצן משונע בדם בשתי צורות:
- חמצן מומס בפלזמה ובציטופלזמה של הכדורית האדומה
- חמצן קשור להמוגלובין
התגובה הבסיסית היא:
\[\ce{O2 + Hb <=> HbO2}\]חמצן פחות מסיס מ־$\ce{CO2}$, ולכן קשה לשנע כמות גדולה שלו רק בצורה מומסת. הפתרון הוא המוגלובין.
כשרמת ההמוגלובין תקינה, בערך 98% מהחמצן בדם קשור להמוגלובין, ורק 2% נמצא בצורה מומסת. למרות שהחלק המומס קטן, הוא חשוב מאוד, כי הוא זה שקובע את $\mathrm{P} _\ce{O2}$ והוא זה שמחזיק את שיווי המשקל עם החמצן הקשור להמוגלובין.
- אם $\mathrm{P} _\ce{O2}$ המומס יורד, התגובה נעה לכיוון פירוק $\ce{HbO2}$ (חמצן שקשור להמוגלובין), והמוגלובין משחרר חמצן
- אם $\mathrm{P} _\ce{O2}$ עולה, יותר חמצן נקשר להמוגלובין
לכן אין הפרדה אמיתית בין החלק המומס לחלק הקשור; החלק המומס קובע את הלחץ החלקי ואת כיוון שיווי המשקל.
סטורציה ו־$\mathrm{P} _\ce{O2}$ מומס
סטורציה $\left(\mathrm{Sa} _\ce{O2}\right)$ היא ריווי ההמוגלובין בחמצן: כמה אתרי קישור מתוך כל האתרים של ההמוגלובין תפוסים על ידי חמצן.
מולקולת המוגלובין אחת יכולה לקשור עד ארבע מולקולות חמצן; יש בה ארבע שרשראות גלובין, וכל שרשרת מכילה טבעת פורפירין אחת שיכולה לקשור מולקולת חמצן אחת.
לכן:
- המוגלובין שקשר מולקולת חמצן אחת מתוך ארבע הוא בריווי של 25%
- המוגלובין שקשר שתי מולקולות חמצן הוא בריווי של 50%
- המוגלובין שקשר שלוש מולקולות חמצן הוא בריווי של 75%
- המוגלובין שקשר ארבע מולקולות חמצן הוא בריווי של 100%
אם מסתכלים על כלל ההמוגלובין בדם, סטורציה של 75% פירושה ש־75% מכל אתרי הקישור האפשריים תפוסים על ידי חמצן.
סטורציה היא לא “אחוז החמצן בדם” - היא אחוז אתרי הקישור של ההמוגלובין שתפוסים על ידי חמצן.
כשמודדים סטורציה באצבע, לרוב מודדים סטורציה של דם עורקי. בבדיקת גזים אפשר למדוד גם $\mathrm{P} _\ce{O2}$ מומס וגם סטורציה בדגימה עורקית, ורידית או קפילרית.
במצב מנוחה, הגוף משתמש בערך ב־25% מתכולת החמצן שמגיעה לרקמות. לכן אם הדם העורקי (oxygenated blood) קרוב ל־100% סטורציה, הדם הוורידי (deoxygenated blood) יהיה בערך 75% סטורציה, כי רוב החמצן שנצרך ירד מהחלק שקשור להמוגלובין.
עקומת הדיסוציאציה של חמצן והמוגלובין
הקשר בין $\mathrm{P} _\ce{O2}$ המומס לבין סטורציית ההמוגלובין הוא לא ליניארי. העקומה סיגמואידית.
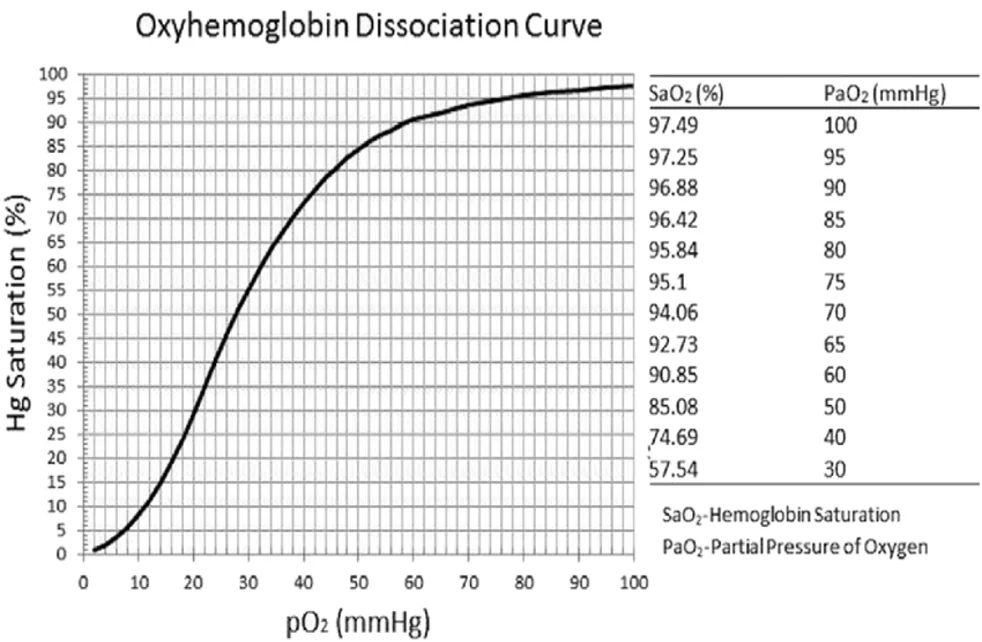
בציר ה־x נמצא ה־$\mathrm{P} _\ce{O2}$ המומס. זהו החמצן המומס, כלומר החלק הקטן יחסית של החמצן בדם (רק ה־2% שלא קשור להמוגלובין).
בציר ה־y נמצאת הסטורציה של ההמוגלובין.
המספרים המרכזיים (באדום ערכים שמוצע לשנן):
| $\mathrm{P} _\ce{O2}$ בדם עורקי או ורידי | סטורציה משוערת |
|---|---|
| $100\, \mathrm{mmHg}$ | 98% |
| $90\, \mathrm{mmHg}$ | 97% |
| $80\, \mathrm{mmHg}$ | 96% |
| $70\, \mathrm{mmHg}$ | 94% |
| $60\, \mathrm{mmHg}$ | 91% |
| $50\, \mathrm{mmHg}$ | 85% |
| $40\, \mathrm{mmHg}$ | 75% |
| $20\, \mathrm{mmHg}$ | 30% |
החלק החשוב בעקומה הוא המדרון בין בערך $20$ ל־$40\, \mathrm{mmHg}$. בטווח הזה שינוי קטן יחסית ב־$\mathrm{P} _\ce{O2}$ גורם לשינוי גדול בסטורציה ובתכולת החמצן הקשורה להמוגלובין.
מעל לבערך $60\text{–}70\, \mathrm{mmHg}$ העקומה משתטחת. לכן העלאה של $\mathrm{P} _\ce{O2}$ מ־$80$ ל־$100\, \mathrm{mmHg}$ כמעט לא משנה סטורציה (ההמוגלובין כבר כמעט בתפוסה מלאה).
גם אם נותנים 100% חמצן ומגיעים ל־$\mathrm{P} _\ce{O2}$ של בערך $660\, \mathrm{mmHg}$, ההמוגלובין לא יכול לעבור 100% סטורציה. אפשר להוסיף עוד חמצן מומס, אבל בגלל המסיסות הנמוכה שלו, התוספת לתכולת החמצן הכוללת לא גדולה כמו שהמספר אולי נשמע.
למה העקומה סיגמואידית?
אחת הסיבות לצורה הסיגמואידית היא קישור קואופרטיבי. כשמולקולת חמצן אחת נקשרת להמוגלובין, היא מעלה את האפיניות של ההמוגלובין לקשור מולקולת חמצן נוספת. לאחר הקישור השני האפיניות עולה עוד, ולאחר הקישור השלישי היא עולה עוד יותר.
לכן בתחילת העקומה העלייה איטית, אחר כך יש עלייה חדה, ובסוף העקומה משתטחת כי רוב אתרי הקישור כבר תפוסים.
$\mathrm{P_{50}}$ והסטת העקומה
$\mathrm{P_{50}}$ הוא הלחץ החלקי של חמצן מומס שבו ההמוגלובין נמצא ב־50% סטורציה.
כשהאפיניות של ההמוגלובין לחמצן עולה, צריך פחות \(\mathrm{P} _\ce{O2}\) כדי להגיע לאותה סטורציה. לכן העקומה מוסטת שמאלה, ו־$\mathrm{P_{50}}$ יורד.
כשהאפיניות של ההמוגלובין לחמצן יורדת, צריך יותר \(\mathrm{P} _\ce{O2}\) כדי להגיע לאותה סטורציה. לכן העקומה מוסטת ימינה, ו־$\mathrm{P_{50}}$ עולה.
האפיניות יורדת (והעקומה זזה ימין) למשל בפעילות גופנית - יש יותר $\ce{CO2}$ ומכאן יותר pH, האפיניות לחמצן יורדת - יותר בצד השמאלי של המשוואה:
\[\ce{Co2 +H2O <=> HCO3- + H}\]
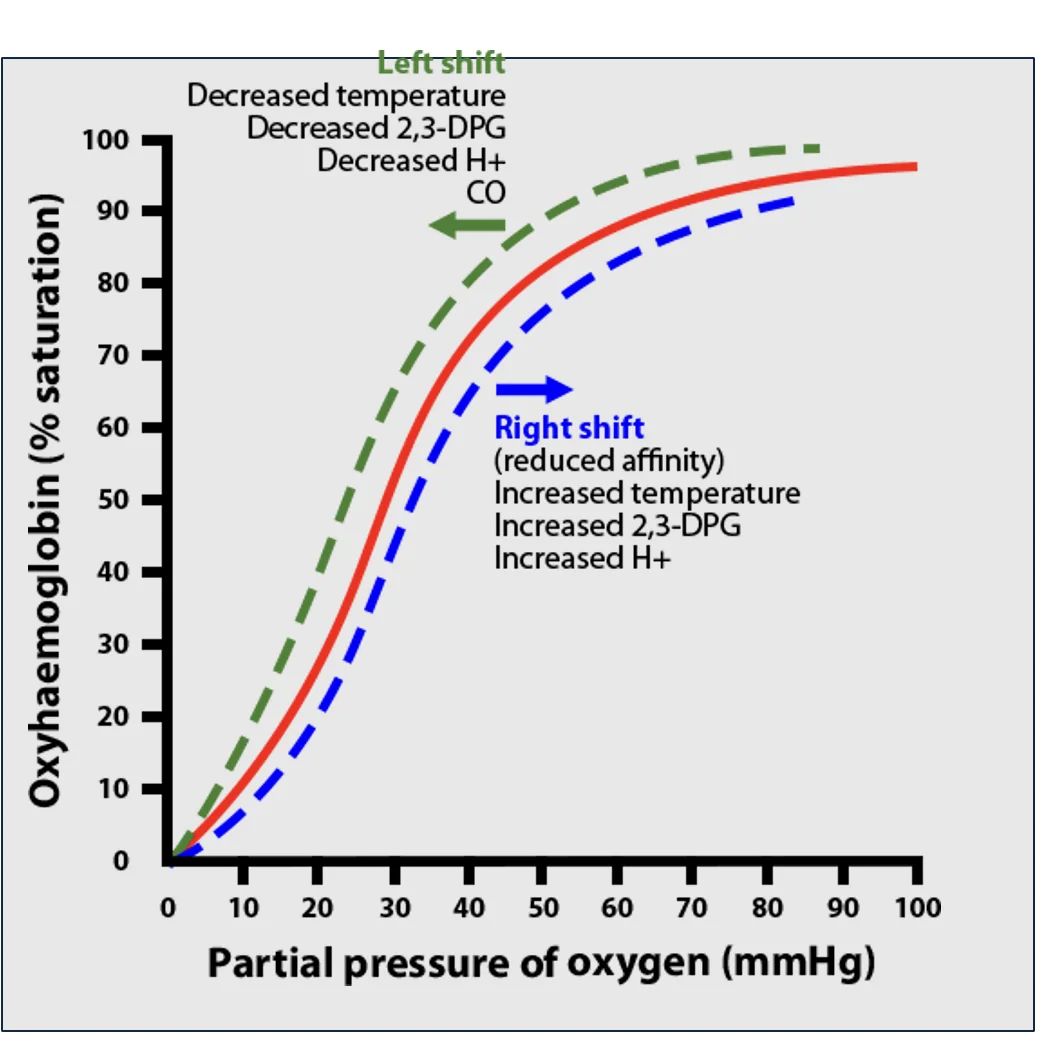
סטייה ימינה
סטייה ימינה פירושה ירידה באפיניות של ההמוגלובין לחמצן. באותו $\mathrm{P} _\ce{O2}$ תהיה סטורציה נמוכה יותר, ולכן ההמוגלובין יפרוק יותר חמצן.
זה מתאים לרקמות פעילות, שבהן רוצים לשחרר חמצן.
גורמים שמסיטים ימינה:
- עלייה ב־$\ce{CO2}$
- ירידה ב־pH
- עלייה בטמפרטורה
- עלייה ב־2,3-DPG
ברקמות, כל הגורמים מתקיימים באופן טבעי: יש יותר $\ce{CO2}$, יותר $\ce{H^+}$, טמפרטורה מעט גבוהה יותר, ובמצבים של צורך מוגבר בחמצן יש יותר 2,3-DPG בכדוריות האדומות.
סטייה שמאלה
סטייה שמאלה פירושה עלייה באפיניות של ההמוגלובין לחמצן. באותו $\mathrm{P} _\ce{O2}$ תהיה סטורציה גבוהה יותר, ולכן ההמוגלובין יחזיק חמצן טוב יותר.
זה מתאים לריאות, שבהן רוצים להעמיס חמצן על ההמוגלובין.
גורמים שמסיטים שמאלה:
- ירידה ב־$\ce{CO2}$
- עלייה ב־pH
- ירידה בטמפרטורה
- ירידה בקישור של 2,3-DPG להמוגלובין
בריאות יש פינוי $\ce{CO2}$, ה־pH עולה, הטמפרטורה מעט נמוכה יותר, והריכוז הגבוה של חמצן מעודד את ההמוגלובין לקשור חמצן במקום להישאר קשור ל־2,3-DPG.
2,3-DPG
2,3-DPG הוא תוצר של גליקוליזה בכדוריות האדומות. הוא נקשר בקלות יחסית ל־Deoxyhemoglobin, כלומר להמוגלובין שאינו מחומצן, ופחות מתאים להמוגלובין מחומצן.
כאשר 2,3-DPG נקשר להמוגלובין, הוא מוריד את האפיניות של ההמוגלובין לחמצן. כך הוא עוזר לפרוק חמצן ברקמות, במיוחד במצבים של חמצן נמוך.
Bohr Effect ו־Haldane Effect
שני האפקטים מתארים שני צדדים של אותה מערכת.
Bohr Effect
ברקמות יש הרבה $\ce{CO2}$ ו־pH נמוך יותר. $\ce{CO2}$ וגם $\ce{H^+}$ מפחיתים את האפיניות של ההמוגלובין לחמצן. לכן ההמוגלובין פורק חמצן ברקמות.
כלומר:
\[\ce{CO2 \uparrow},\ \ce{H^+ \uparrow},\ \mathrm{pH} \downarrow \quad \Rightarrow \quad \ce{O2}\ \text{affinity} \downarrow \quad \Rightarrow \quad \ce{O2}\ \text{unloading}\]Haldane Effect
בריאות יש הרבה חמצן. כאשר חמצן נקשר להמוגלובין, האפיניות של ההמוגלובין ל־$\ce{CO2}$ ול־$\ce{H^+}$ יורדת. לכן ההמוגלובין משחרר $\ce{CO2}$ ו־$\ce{H^+}$, והמערכת דוחפת את יצירת $\ce{CO2}$ מחדש מביקרבונט כדי לפנות אותו בנשיפה.
כלומר:
\[\ce{O2}\ \text{binds to Hb} \quad \Rightarrow \quad \ce{CO2}\ \text{affinity} \downarrow \quad \Rightarrow \quad \ce{CO2}\ \text{removal}\]המוגלובין עוברי
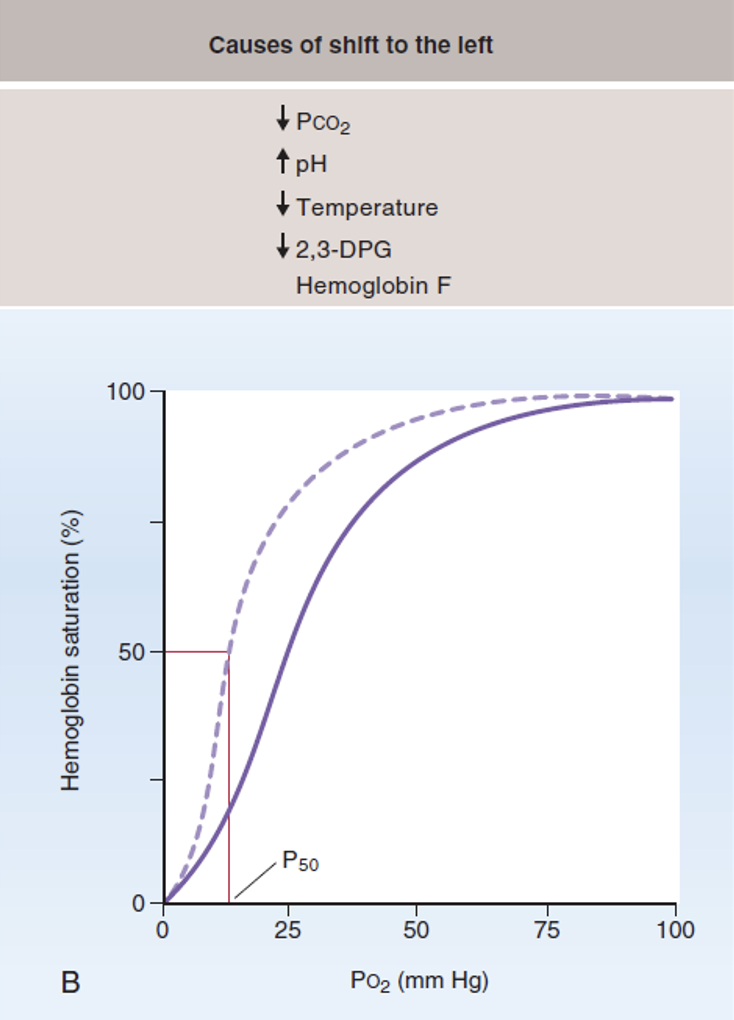
לעובר יש המוגלובין מסוג Hemoglobin F, או המוגלובין עוברי. להמוגלובין הזה יש אפיניות גבוהה יותר לחמצן בהשוואה להמוגלובין רגיל.
הסיבה המרכזית היא שהמוגלובין F לא נקשר היטב ל־2,3-DPG. מכיוון ש־2,3-DPG רגיל להוריד את האפיניות של ההמוגלובין לחמצן, היעדר הקישור אליו משאיר את ההמוגלובין העוברי עם אפיניות גבוהה יותר.
זה מתאים לפיזיולוגיה של העובר: העובר לא נושם דרך הריאות; ה”ריאות” הפיזיולוגיות שלו הן השליה. בשליה, ה־$\mathrm{P} _\ce{O2}$ נמוך יותר מה־$\mathrm{P} _\ce{O2}$ בריאה של אדם שנושם אוויר חדר, ולכן העובר צריך המוגלובין שיכול לקשור חמצן גם בלחץ חלקי נמוך יחסית.
למרות האפיניות הגבוהה, המוגלובין F עדיין יכול לפרוק חמצן ברקמות העובר, כי שם $\mathrm{P} _\ce{O2}$ נמוך מאוד. כשהלחץ החלקי מספיק נמוך, גם המוגלובין עם אפיניות גבוהה ישחרר חמצן.
A-a Gradient
A-a Gradient הוא ההפרש בין הלחץ החלקי של חמצן בנאדיות לבין הלחץ החלקי של חמצן בדם העורקי.
\[\mathrm{A\text{-}a\ Gradient} = \mathrm{P\scriptstyle{A}} _\ce{O2} - \mathrm{Pa} _\ce{O2}\]כאשר:
- $\mathrm{P\scriptstyle{A}} _\ce{O2}$ - הלחץ החלקי של חמצן בנאדית (A = Alveolar)
- $\mathrm{Pa} _\ce{O2}$ - הלחץ החלקי של חמצן בדם העורקי (a = arterial)
באידיאל, אם הפעפוע תקין לגמרי, היינו מצפים שהלחץ החלקי של החמצן בדם העורקי יהיה כמעט זהה ללחץ החלקי בנאדית.
בפועל, ה־A-a gradient אינו אפס. יש תמיד הפרש קטן, בערך $15\text{–}20\, \mathrm{mmHg}$, בין היתר בגלל ערבוב קטן עם דם פחות מחומצן. דם שמספק את הברונכוסים עצמם דרך המערכת הסיסטמית יכול לחזור לצד שמאל, ויש גם תרומה של אזורי shunt קטנים בריאה.
המשוואה האלוואולרית
כדי לחשב את $\mathrm{P\scriptstyle{A}} _\ce{O2}$ אפשר להשתמש במשוואת הגזים האלוואולרית:
\[\mathrm{P\scriptstyle{A}} _\ce{O2} = F_{I\ce{O2}}(\mathrm{P_{atm}} - \mathrm{P_{\ce{H2O}}}) - \frac{\mathrm{Pa} _\ce{CO2}}{\mathrm{RR}}\]בשאלות פשוטות, כאשר $\mathrm{Pa} _\ce{CO2}$ תקין, אפשר להעריך מהר את $\mathrm{P\scriptstyle{A}} _\ce{O2}$ לפי ריכוז החמצן הנשאף:
\[\mathrm{P\scriptstyle{A}} _\ce{O2} \approx F_{I\ce{O2}}(\%) \times 5\text{ to }6\]דוגמאות:
| $F_{I\ce{O2}}$ | הערכת $\mathrm{P\scriptstyle{A}} _\ce{O2}$ |
|---|---|
| 21% - אוויר חדר | $100\, \mathrm{mmHg}$ |
| 40% | $200\text{–}240\, \mathrm{mmHg}$ |
| 50% | $250\text{–}300\, \mathrm{mmHg}$ |
| 100% | $660\, \mathrm{mmHg}$ |
דוגמה קלינית
אם אדם מקבל 100% חמצן, הצפי הוא $\mathrm{P\scriptstyle{A}} _\ce{O2}$ של בערך $660\, \mathrm{mmHg}$. אם הסטורציה שלו היא 90%, אפשר להעריך לפי העקומה שה־$\mathrm{Pa} _\ce{O2}$ שלו בערך $60\, \mathrm{mmHg}$.
לכן:
\[\mathrm{A\text{-}a\ Gradient} \approx 660 - 60 = 600\, \mathrm{mmHg}\]זה פער עצום, והוא מצביע על בעיה משמעותית בשחלוף הגזים. המשמעות הקלינית היא שלא מספיק להסתכל רק על הסטורציה. צריך לשאול כמה חמצן האדם מקבל כדי להגיע לסטורציה הזו.
אדם עם סטורציה של 93% על אוויר חדר אינו דומה לאדם עם סטורציה 93% תחת 100% חמצן. במקרה השני, הריאה עובדת הרבה פחות טוב, גם אם המספר על מד הסטורציה עדיין נראה “סביר”.
מד סטורציה
מד סטורציה מודד את ריווי החמצן של ההמוגלובין בדם העורקי. העיקרון הוא שהמוגלובין מחומצן והמוגלובין לא מחומצן סופגים אור באורכי גל שונים.
המכשיר מעביר אור דרך הרקמה, בדרך כלל דרך האצבע, ומזהה את ההבדל בין ספיגת האור של המוגלובין מחומצן לבין המוגלובין לא מחומצן. הוא מתמקד באות הפועם, כלומר בדם העורקי שמגיע בכל פעימה, ומנסה להפחית את רעש הרקע של רקמות, דם ורידי ודם קפילרי.
היחס הבסיסי הוא:
\[\text{Saturation} = \frac{\text{Oxyhemoglobin}}{\text{Oxyhemoglobin} + \text{Deoxyhemoglobin}}\]בנוסף למספר הסטורציה, המכשיר נותן גם גל דופק. איכות הגל חשובה, כי היא עוזרת להבין האם המדידה אמינה.
גורמים שיכולים לפגוע באמינות המדידה:
- לק בציפורניים
- קור או פרפוזיה ירודה לאצבע
- תנועה
- עור כהה יכול להשפיע מעט במצבים מסוימים
- סטורציות נמוכות מאוד, בעיקר מתחת לכ־80% עד 85%, פחות אמינות
בסטורציות נמוכות מאוד, אם המכשיר מציג למשל 70%, המספר כבר פחות מדויק; הסטורציה האמיתית יכולה להיות נמוכה יותר.
מד הסטורציה גם משמיע צליל המשתנה לפי הסטורציה. כאשר הסטורציה גבוהה הצליל גבוה יותר, וכשהסטורציה יורדת הצליל יורד. זה מאפשר לצוות לזהות ירידה גם בלי להביט כל הזמן במסך.
תכולת חמצן - Oxygen Content
תכולת חמצן היא כמות החמצן הכוללת שיש בדם. היא כוללת שני רכיבים:
- חמצן הקשור להמוגלובין
- חמצן מומס בפלזמה
הנוסחה:
\[\mathrm{Ca} _\ce{O2} = \underbrace{(\mathrm{Hb} \times 1.34 \times \mathrm{Sa} _\ce{O2})}_{\text{bound to hemoglobin}} + \underbrace{(0.003 \times \mathrm{Pa} _\ce{O2})}_{\text{dissolved in plasma}}\]כאשר:
- $\mathrm{Ca} _\ce{O2}$ - תכולת החמצן בדם עורקי, ביחידות של $\left[\mathrm{mL\,\ce{O2}/dL\ blood}\right]$
- $\mathrm{Hb}$ - ריכוז ההמוגלובין, ב־$\left[\mathrm{g/dL}\right]$
- $1.34$ - כמות החמצן שגרם אחד של המוגלובין יכול לקשור כשהוא רווי לגמרי
- $\mathrm{Sa} _\ce{O2}$ - הסטורציה כשבר עשרוני, למשל 97% = 0.97
- $0.003$ - מקדם המסיסות של חמצן בדם
- $\mathrm{Pa} _\ce{O2}$ - הלחץ החלקי של חמצן מומס בדם עורקי
מאיפה מגיע 1.34?
המספר $1.34\, \left[\mathrm{mL\, O2/g\, Hb}\right]$ אומר כמה חמצן יכול להיקשר לגרם אחד של המוגלובין בריווי מלא. זו דרך לכמת מספר מולקולות חמצן לפי הנפח שהיו תופסות במצב גזי בתנאים מוגדרים.
אפשר לחשוב על זה דרך חוק הגזים:
\[\mathrm{PV = nRT}\]כשהלחץ והטמפרטורה קבועים, נפח הגז מייצג את כמות החלקיקים. לכן מדברים על $\left[\mathrm{mL}\,\ce{O2}\right]$ גם כשהחמצן נמצא בפועל קשור להמוגלובין ולא כבועות גז בדם.
דוגמה עם המוגלובין תקין
נניח:
- $\mathrm{Hb} = 15\, \left[\mathrm{g/dL}\right]$
- $\mathrm{Sa} _\ce{O2} = 0.97$
- $\mathrm{Pa} _\ce{O2} = 100\, \mathrm{mmHg}$
הרכיב שקשור להמוגלובין:
\[15 \times 1.34 \times 0.97 \approx 19.5\, \left[\mathrm{mL\, O2/dL}\right]\]הרכיב המומס:
\[0.003 \times 100 = 0.3\, \left[\mathrm{mL\, O2/dL}\right]\]סה”כ:
\[\mathrm{Ca} _\ce{O2} \approx 19.8\, \left[\mathrm{mL\, O2/dL}\right]\]כלומר, במצב רגיל:
- בערך $19.5\, \left[\mathrm{mL\, O2/dL}\right]$ קשורים להמוגלובין
- בערך $0.3\, \left[\mathrm{mL\, O2/dL}\right]$ מומסים בפלזמה
- החמצן המומס הוא בערך 1.5% מסך תכולת החמצן
| Hb | סטורציה | O2 קשור | $\mathrm{Pa} _\ce{O2}$ | O2 מומס | סה״כ O2 | %קשור | %מומס |
|---|---|---|---|---|---|---|---|
| $15\, \left[\mathrm{g/dL}\right]$ | 0.97 | 19.50 | 100 | 0.30 | 19.80 | 98.48% | 1.52% |
| $15\, \left[\mathrm{g/dL}\right]$ | 1.00 | 20.10 | 660 | 1.98 | 22.08 | 91.03% | 8.97% |
| $3\, \left[\mathrm{g/dL}\right]$ | 0.97 | 3.90 | 100 | 0.30 | 4.20 | 92.86% | 7.14% |
| $3\, \left[\mathrm{g/dL}\right]$ | 1.00 | 4.02 | 660 | 1.98 | 6.00 | 67.00% | 33.00% |
מתן 100% חמצן
אם נותנים 100% חמצן לאדם עם המוגלובין תקין ושחלוף גזים תקין, אפשר להגיע ל־$\mathrm{Pa} _\ce{O2}$ של בערך $660\, \mathrm{mmHg}$ וסטורציה של 100%.
אז הרכיב שקשור להמוגלובין:
\[15 \times 1.34 \times 1 = 20.1\, \left[\mathrm{mL\, O2/dL}\right]\]והחמצן המומס:
\[0.003 \times 660 = 1.98\, \left[\mathrm{mL\, O2/dL}\right]\]סה”כ:
\[\mathrm{Ca} _\ce{O2} \approx 22.08\, \left[\mathrm{mL\, O2/dL}\right]\]כלומר, מתן 100% חמצן מעלה את החלק המומס, אבל כשההמוגלובין כבר כמעט רווי, התוספת הכוללת לתכולת החמצן מוגבלת.
אנמיה, סטורציה ותכולת חמצן
אדם יכול להיות עם סטורציה תקינה ועם $\mathrm{Pa} _\ce{O2}$ תקין, ועדיין להיות עם תכולת חמצן נמוכה מאוד אם אין לו מספיק המוגלובין.
לדוגמה, אם יש המוגלובין $3\, \left[\mathrm{g/dL}\right]$ במקום $15\, \left[\mathrm{g/dL}\right]$, הסטורציה יכולה עדיין להיות 97%, כי ההמוגלובין שכן קיים רווי בחמצן. הבעיה היא שיש מעט מאוד המוגלובין, ולכן מעט מאוד אתרי קישור.
במצב כזה אין בהכרח בעיה בשחלוף הגזים בריאה. הבעיה היא בתכולת החמצן ובאספקת החמצן לרקמות.
מתן 100% חמצן יכול להעלות את החמצן המומס, ולכן הוא יכול לעזור מעט. אבל זו תוספת מוגבלת, כי חמצן מסיס מעט מאוד בפלזמה. כאשר חסר המוגלובין, הפתרון העיקרי אינו להעמיס עוד ועוד חמצן מומס, אלא לטפל בחסר ההמוגלובין.
עקומת תכולת החמצן
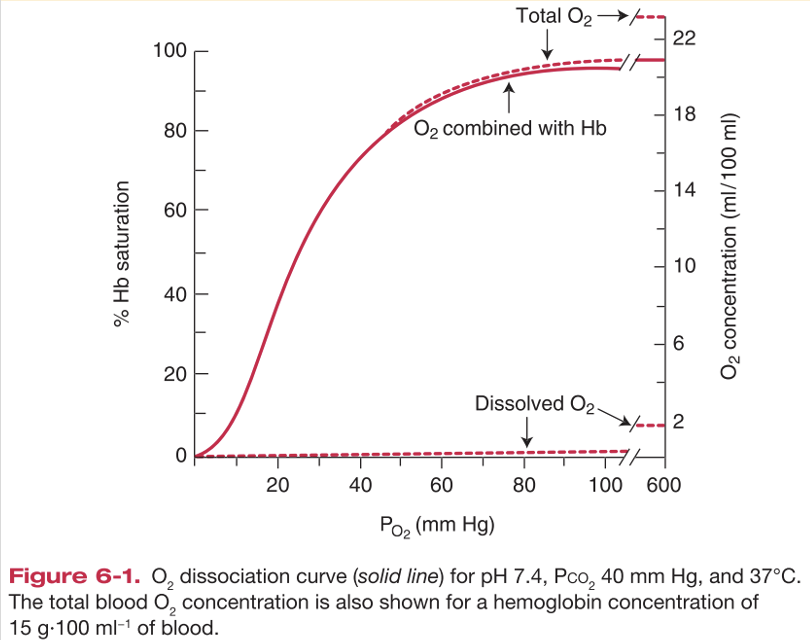
בעקומת תכולת החמצן אפשר להציג שלושה דברים יחד:
- ציר x: $\mathrm{P} _\ce{O2}$ מומס
- ציר y שמאלי: סטורציה
- ציר y ימני: תכולת חמצן ב־$\left[\mathrm{mL\, O2/dL\ blood}\right]$
הקו של הסטורציה והקו של החמצן הקשור להמוגלובין דומים מאוד, כי רוב תכולת החמצן מגיעה מהחמצן שקשור להמוגלובין.
החמצן המומס עולה בצורה כמעט ליניארית עם $\mathrm{P} _\ce{O2}$, אבל הוא נשאר רכיב קטן כל עוד ריכוז ההמוגלובין תקין.
כאשר ההמוגלובין נמוך, עקומת הסטורציה עצמה לא משתנה, כי סטורציה היא יחס. אבל ציר תכולת החמצן כן משתנה, כי יש פחות המוגלובין ולכן פחות קיבולת חמצן.
תכולת חמצן, תכולת CO2 ו־RQ
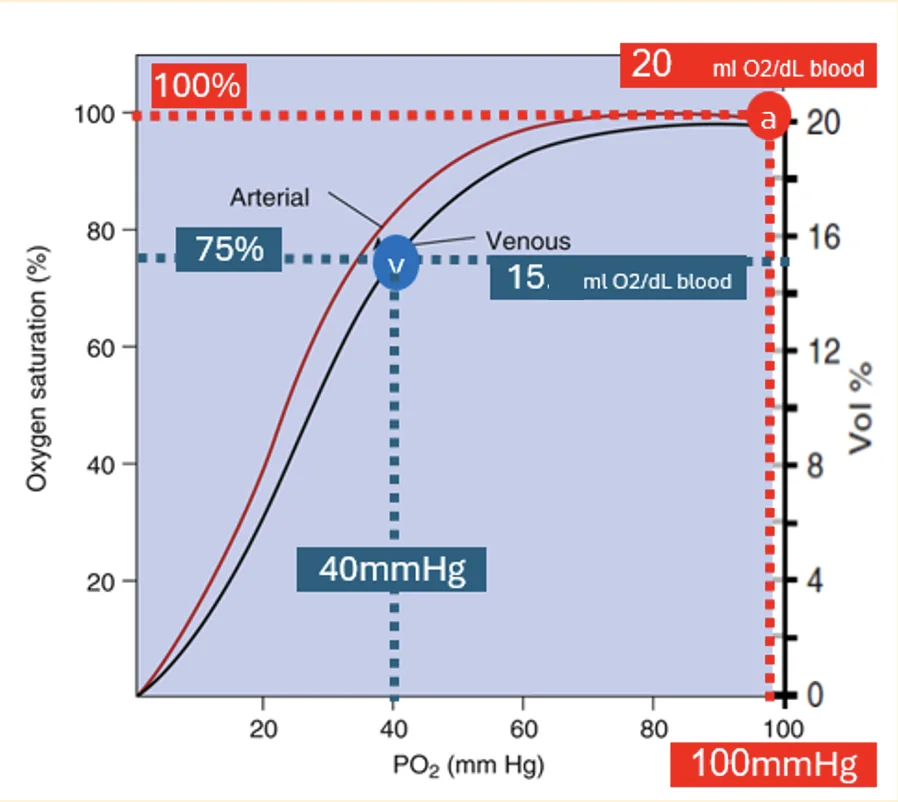
בדם עורקי תקין, כאשר $\mathrm{Hb}$ בערך $15\, \left[\mathrm{g/dL}\right]$, תכולת החמצן היא בערך:
\[20 \left[\mathrm{mL\, O2/dL\, blood}\right]\]בדם ורידי מעורב, אחרי שהרקמות צרכו חמצן, תכולת החמצן היא בערך:
\[15 \left[\mathrm{mL\, O2/dL\, blood}\right]\]כלומר, בכל $100\, \mathrm{mL}$ דם הגוף משתמש בערך ב־$5\, \mathrm{mL}$ חמצן.
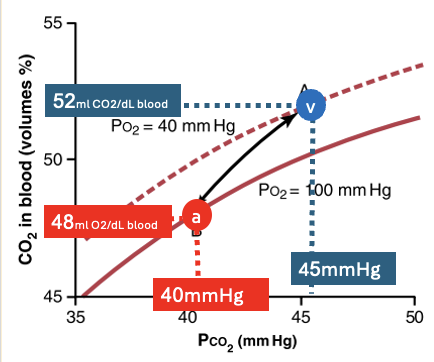
במקביל, תכולת $\ce{CO2}$ בדם עורקי היא בערך:
\[48\, \left[\mathrm{mL\, CO2/dL\ blood}\right]\]ובדם ורידי היא בערך:
\[52\, \left[\mathrm{mL\, CO2/dL\ blood}\right]\]כלומר, בכל $100\, \mathrm{mL}$ דם מתווספים בערך $4\, \mathrm{mL}\, \ce{CO2}$.
היחס בין ייצור $\ce{CO2}$ לבין צריכת $\ce{O2}$ הוא Respiratory Quotient, או RQ:
\[\mathrm{RQ} = \frac{\mathrm{\dot V_{\ce{CO2}}}}{\mathrm{\dot V_{\ce{O2}}}}\]במצב מנוחה רגיל:
\[\mathrm{RQ} \approx \frac{4}{5} = 0.8\]אותו יחס מופיע בכמה רמות:
- ברמה המולקולרית: על כל 5 יחידות חמצן שנצרכות, נוצרות בערך 4 יחידות $\ce{CO2}$
- ברמת דציליטר דם: שימוש בכ־$5\, \left[\mathrm{mL\, O2/dL}\right]$ וייצור כ־$4\, \left[\mathrm{mL\, CO2/dL}\right]$
- בזמן אמת: צריכת חמצן וייצור $\ce{CO2}$ לדקה
Oxygen Delivery ו־Oxygen Extraction Ratio
כדי לעבור מתכולת חמצן לזרימה בזמן אמת, צריך להכניס את תפוקת הלב.
תפוקת לב ממוצעת במנוחה היא בערך:
\[5\, \left[\mathrm{L/min}\right] = 50\, \left[\mathrm{dL/min}\right]\]אם בכל דציליטר דם יש בערך $20 \left[\mathrm{mL\, O2}\right]$, אז אספקת החמצן לרקמות היא:
\[20 \left[\frac{\mathrm{mL\, O2}}{\mathrm{dL}}\right] \times 50 \left[\frac{\mathrm{dL}}{\mathrm{min}}\right] = 1000 \left[\mathrm{mL\, O2/min}\right]\]זהו Oxygen Delivery, או $\mathrm{\dot D_{\ce{O2}}}$.
אבל הגוף לא משתמש בכל החמצן שהוא מספק. במנוחה הוא משתמש בערך ב־$5 \left[\mathrm{mL\, O2/dL}\right]$ מתוך $20 \left[\mathrm{mL\, O2/dL}\right]$.
לכן צריכת החמצן היא:
\[5 \left[\frac{\mathrm{mL\, O2}}{\mathrm{dL}}\right] \times 50 \left[\frac{\mathrm{dL}}{\mathrm{min}}\right] = 250 \left[\mathrm{mL\, O2/min}\right]\]אם בכל דציליטר נוצרים בערך $4 \left[\mathrm{mL\, CO2}\right]$, אז ייצור $\ce{CO2}$ הוא:
\[4 \left[\frac{\mathrm{mL\, CO2}}{\mathrm{dL}}\right] \times 50 \left[\frac{\mathrm{dL}}{\mathrm{min}}\right] = 200 \left[\mathrm{mL\, CO2/min}\right]\]ולכן:
\[\mathrm{RQ} = \frac{200}{250} = 0.8\]Oxygen Extraction Ratio הוא היחס בין צריכת החמצן לבין אספקת החמצן:
\[\mathrm{O_2\ Extraction\ Ratio} = \frac{\mathrm{\dot V_{\ce{O2}}}}{\mathrm{\dot D_{\ce{O2}}}}\]במנוחה:
\[\frac{250}{1000} = 0.25 = 25\%\]במאמץ או במצבים שבהם אספקת החמצן מוגבלת, הגוף יכול לשלוף אחוז גבוה יותר מהחמצן שמגיע לרקמות. כאשר השליפה גבוהה מאוד, הגוף מתקרב למגבלה, ואז עלול להידרש יותר מטבוליזם אנאירובי.
במאמץ משמעותי, צריכת החמצן יכולה לעלות מאוד. אצל ספורטאים היא יכולה להגיע לערכים גבוהים בהרבה ממנוחה. במאמץ מתון הגוף מעלה גם את האוורור וגם את תפוקת הלב כדי לשמור על האיזון. במאמץ קשה יותר, ובעיקר כשמשתמשים יותר בגליקוגן ובפחמימות, RQ יכול להתקרב ל־1.
היפוקסמיה לעומת היפוקסיה
היפוקסמיה היא כמות חמצן נמוכה בדם.
היפוקסיה היא כמות חמצן נמוכה ברקמות.
בדרך כלל, אם אדם היפוקסמי, יש סיכוי טוב שגם הרקמות שלו יהיו היפוקסיות. אבל זה לא תמיד אותו דבר.
יכול להיות מצב שבו החמצן בדם תקין, אבל הרקמות לא מקבלות מספיק חמצן בגלל בעיית זרימה. לדוגמה, אם הלב לא מצליח להזרים דם כמו שצריך, תכולת החמצן בדם יכולה להיות תקינה, שחלוף הגזים בריאה יכול להיות תקין, ועדיין תהיה היפוקסיה ברקמות.
לכן צריך להבדיל בין:
- בעיית חמצון בדם
- בעיית אספקת חמצן לרקמות
- בעיית ניצול חמצן ברקמות
אוורור, דילול גזים ושיווי משקל חדש
בכל נשימה רגילה נכנסים בערך $500\, \mathrm{mL}$ אוויר, אבל רק כ־$350\, \mathrm{mL}$ מגיעים לנאדיות. כ־$150\, \mathrm{mL}$ נשארים בחלל המת.
נפח האוויר שנמצא בריאות בסוף נשיפה רגילה גדול בהרבה, בערך כמה ליטרים. לכן נשימה אחת לא מחליפה את כל הגז בנאדיות; השינוי בהרכב הגזים מתרחש בהדרגה דרך דילול.
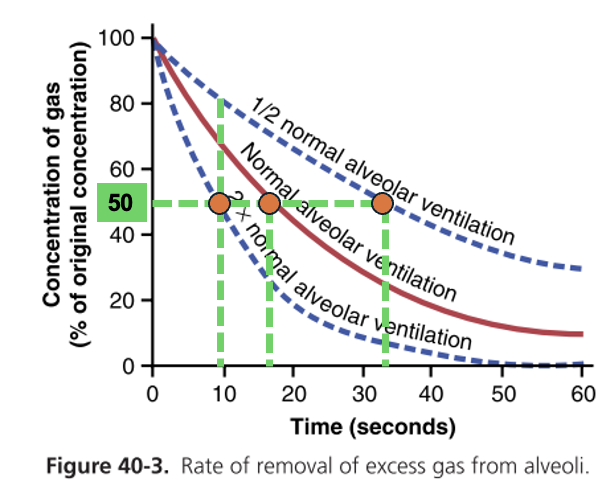
אם היה גז מסוים בנאדיות שאינו נכנס לגוף ואינו מיוצר מחדש, והריאות היו מתחילות לאוורר אותו, הריכוז שלו היה יורד בהדרגה בצורה גיאומטרית. בהתחלה הירידה גדולה יותר, ואז כל ירידה נוספת קטנה יותר.
האנלוגיה שניתנה היא כוס עם מיץ פטל:
- יש כוס עם ריכוז מסוים של פטל.
- בכל מחזור מוציאים חלק מהנוזל ומוסיפים מים.
- אם יש גם ייצור קבוע של פטל, למשל הוספת כפית בכל מחזור, אפשר להגיע לשיווי משקל.
- אם מתחילים מריכוז גבוה, הריכוז יורד מהר בהתחלה ואז מתקרב בהדרגה לשיווי המשקל.
- אם קצב הייצור עולה, שיווי המשקל החדש יהיה בריכוז גבוה יותר, אלא אם גם קצב הדילול עולה.
כך גם עם $\ce{CO2}$:
- אם קצב ייצור $\ce{CO2}$ קבוע והאוורור עולה, $\mathrm{P_{\ce{CO2}}}$ יורד.
- אם קצב ייצור $\ce{CO2}$ קבוע והאוורור יורד, $\mathrm{P_{\ce{CO2}}}$ עולה.
- העלייה אינה אינסופית; הגוף מגיע לשיווי משקל חדש רק עם $\mathrm{P_{\ce{CO2}}}$ גבוה יותר.
כלל פשוט:
\[\boxed{\mathrm{P\scriptstyle{A}} _\ce{CO2} \propto \frac{\mathrm{\dot V_ \ce{CO2}}}{\mathrm{\dot V_A}}}\]כלומר, הלחץ החלקי של $\ce{CO2}$ בנאדיות עולה כשייצור $\ce{CO2}$ עולה, ויורד כשהאוורור הנאדיתי (MV) עולה.
אם האוורור הנאדיתי מוכפל וקצב ייצור $\ce{CO2}$ לא משתנה, $\mathrm{P\scriptstyle{A}} _\ce{CO2}$ ירד בערך בחצי. אם האוורור יורד בחצי, $\mathrm{P\scriptstyle{A}} _\ce{CO2}$ יעלה בערך פי שניים.
אוורור ביחס לייצור $\ce{CO2}$ ולצריכת $\ce{O2}$
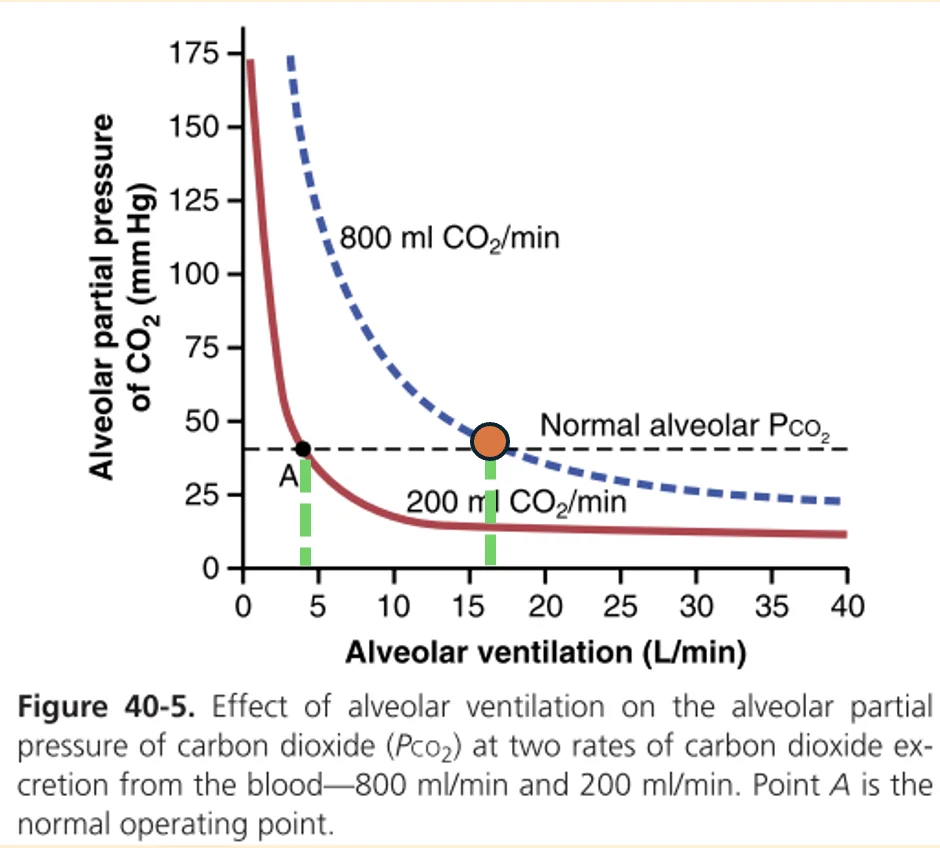
במצב מנוחה, קצב ייצור $\ce{CO2}$ הוא בערך:
\[200\ \left[\mathrm{mL\, CO2/min}\right]\]כדי לשמור על $\mathrm{P\scriptstyle{A}} _\ce{CO2}$ של בערך $40\, \mathrm{mmHg}$, צריך אוורור נאדיתי של בערך:
\[4.2\ \left[\mathrm{L/min}\right]\]אם ייצור $\ce{CO2}$ עולה פי 4, למשל במאמץ, ל־$800 \left[\mathrm{mL\, CO2/min}\right]$, צריך להעלות גם את האוורור פי 4 כדי לשמור על אותו $\mathrm{P\scriptstyle{A}} _\ce{CO2}$:
\[4.2 \times 4 = 16.8\, \left[\mathrm{L/min}\right]\]אותו עיקרון חל גם על חמצן.
במנוחה, צריכת החמצן היא בערך:
\[250\, \left[\mathrm{mL\, O2/min}\right]\]אם צריכת החמצן עולה פי 4 ל־$1000 \left[\mathrm{mL\, O2/min}\right]$, אבל האוורור לא עולה בהתאם, הריאות ימשיכו לספק חמצן לאט מדי ביחס לצריכה. במקרה כזה $\mathrm{P\scriptstyle{A}} _\ce{O2}$ ירד.
כדי לשמור על $\mathrm{P\scriptstyle{A}} _\ce{O2}$ סביב $100\, \mathrm{mmHg}$ כאשר צריכת החמצן עולה פי 4, צריך להעלות גם את האוורור בערך פי 4.
במילים פשוטות: אם המטבוליזם עולה, גם ייצור $\ce{CO2}$ עולה וגם צריכת $\ce{O2}$ עולה. כדי לשמור על הרכב הגזים בנאדיות, האוורור צריך לעלות בהתאם.
למה שינויים ב־$\mathrm{P} _\ce{O2}$ פחות דרמטיים בהתחלה?
ההמוגלובין נותן סוג של בופר לחמצן. כאשר $\mathrm{P} _\ce{O2}$ עולה מעל 100 ל־130, למשל בגלל היפרוונטילציה שמורידה $\ce{CO2}$, תכולת החמצן הכוללת כמעט לא משתפרת, כי ההמוגלובין כבר כמעט רווי.
גם בכיוון ההפוך, אם $\mathrm{P} _\ce{O2}$ יורד עד בערך 60, הסטורציה עדיין סביב 90% עד 93%, ולכן תכולת החמצן לא נופלת מיד בצורה קיצונית. אבל מתחת לאזור הזה נכנסים לחלק התלול של העקומה, ואז ירידה נוספת ב־$\mathrm{P} _\ce{O2}$ גורמת לנפילה חדה בסטורציה ובתכולת החמצן.
היפרוונטילציה ותחושת נימול
בהיפרוונטילציה, למשל בזמן התקף חרדה, מפנים יותר $\ce{CO2}$. זה לא גורם להרעלת חמצן. הבעיה העיקרית היא ירידה ב־$\ce{CO2}$ ועלייה ב־pH, כלומר בססת נשימתית.
בססת יכולה לשנות את מצב הקלציום בדם: יותר קלציום נקשר לאלבומין, ולכן יש פחות קלציום יוני חופשי. זה יכול לגרום לתחושות כמו נימול, כיווץ שרירים או טטניה. זה לא נובע מעודף חמצן.
בקרת הנשימה
מערכת הנשימה צריכה להתאים את האוורור לצרכים המטבוליים. הבקרה העיקרית נעשית לפי $\ce{CO2}$, חמצן ו־pH.
הבקרה המרכזית - $\ce{CO2}$ ו־pH במוח
גזע המוח רגיש בעיקר לשינויים ב־$\ce{CO2}$, אבל בפועל הוא מגיב להשפעה של $\ce{CO2}$ על ה־pH.
הנקודה החשובה היא ש־$\ce{CO2}$ חוצה את מחסום הדם-מוח בקלות, ואילו $\ce{H^+}$ לא חוצה אותו בקלות. לכן, כאשר $\ce{CO2}$ עולה בדם, הוא חוצה אל הנוזל סביב המוח, הופך שם לחומצה פחמתית, מעלה $\ce{H^+}$, מוריד pH, ומגרה את מרכזי הנשימה להגביר אוורור.
אם יש ירידת pH מסיבה מטבולית שאינה $\ce{CO2}$, גם היא יכולה להגביר אוורור, אבל התגובה ל־$\ce{CO2}$ חזקה יותר, כי $\ce{CO2}$ חוצה את ה־BBB בקלות.
שינויים קטנים יחסית ב־$\mathrm{P_{\ce{CO2}}}$, אפילו סביב $3\, \mathrm{mmHg}$, יכולים לגרום לשינוי משמעותי באוורור, במיוחד בטווח של בערך $35\text{–}75\, \mathrm{mmHg}$.
הבקרה ההיקפית - חמצן
קולטני חמצן חשובים נמצאים ב־Carotid Bodies וב־Aortic Bodies. הם רגישים בעיקר לירידה משמעותית ב־$\mathrm{P} _\ce{O2}$.
התגובה לחמצן נעשית משמעותית בעיקר כאשר:
\[\mathrm{Pa} _\ce{O2} < 60\, \mathrm{mmHg}\]זה מתאים בערך לסטורציה של 90%. זה גם האזור שבו עקומת ההמוגלובין־חמצן מתחילה להיות תלולה יותר, ולכן הגוף מתחיל להיות בסכנה של ירידה מהירה בתכולת החמצן.
גובה והסתגלות
בגובה רב, אחוז החמצן באוויר עדיין בערך 21%, אבל הלחץ האטמוספרי נמוך יותר. לכן הלחץ החלקי של החמצן שנשאף נמוך יותר.
כאשר $\mathrm{P} _\ce{O2}$ יורד, קולטני החמצן מעודדים עלייה באוורור. אבל היפרוונטילציה מורידה $\ce{CO2}$, וירידת $\ce{CO2}$ גורמת לעליית pH, כלומר בססת נשימתית. ה־pH הגבוה יכול לדכא את הדחף לנשום.
לכן בטיפוס מהיר לגובה יש התנגשות בין שני אותות:
- חמצן נמוך אומר: לנשום יותר
- $\ce{CO2}$ נמוך ו־pH גבוה אומרים: להאט את הנשימה
בטיפוס הדרגתי הגוף מסתגל. הכליות מפרישות יותר ביקרבונט בשתן, וכך הן עוזרות לתקן את הבססת הנשימתית. כאשר ה־pH מתקרב חזרה לנורמה, המוח פחות “נלחם” בהיפרוונטילציה, ואפשר לשמר אוורור גבוה יותר בגובה.
שילוב בין $\ce{CO2}$, חמצן ו־pH
התגובה לאוורור לא נקבעת מגורם אחד בלבד: כאשר $\ce{CO2}$ עולה, האוורור עולה; כאשר חמצן יורד; האוורור עולה; כאשר pH יורד, האוורור עולה.
השילובים חשובים:
- אם $\mathrm{Pa} _\ce{O2}$ תקין, עלייה ב־$\mathrm{Pa} _\ce{CO2}$ תעלה את האוורור לפי עקומת תגובה מסוימת.
- אם $\mathrm{Pa} _\ce{O2}$ נמוך, אותה עלייה ב־$\mathrm{Pa} _\ce{CO2}$ תגרום לתגובה חזקה יותר באוורור.
- אם pH נמוך מסיבה שאינה $\ce{CO2}$, הגוף משנה את נקודת העבודה שלו ומנסה לשמר $\mathrm{P_{\ce{CO2}}}$ נמוך יותר כדי להעלות pH.
- אם גם pH נמוך וגם חמצן נמוך, התגובה ל־$\ce{CO2}$ נעשית חזקה עוד יותר.
במילים פשוטות: אם יש חמצן נמוך, רוצים לנשום יותר. אם יש pH נמוך, רוצים לנשום יותר. אם יש $\ce{CO2}$ גבוה, רוצים לנשום יותר. כשכמה מהגורמים הללו מופיעים יחד, תגובת האוורור חזקה יותר.
דור פסקל