פוטנציאל מנוחה של הממברנה (Resting Membrane Potential)
בין פנים התא לחוץ התא קיימים הבדלים בריכוזי יונים:
| יון / חומר | בתוך התא $(-)$ | מחוץ לתא $(+)$ |
|---|---|---|
| נתרן $\ce{Na+}$ | נמוך | גבוה |
| אשלגן $\ce{K+}$ | גבוה | נמוך |
| אניונים אורגניים שליליים $\ce{A^-}$ בעיקר חלבונים התורמים לשליליות התוך־תאית | גבוה | נמוך |
| כלוריד $\ce{Cl-}$ בדרך כלל גבוה יותר מחוץ לתא | נמוך | גבוה |
- נתרן $\ce{Na+}$ - גבוה יותר מחוץ לתא
- אשלגן $\ce{K+}$ - גבוה יותר בתוך התא
- כלוריד $\ce{Cl-}$ - גבוה יותר בדרך כלל מחוץ לתא
בתוך התא יש גם אניונים אורגניים שליליים ($\ce{A^-}$), בעיקר חלבונים, שתורמים לשליליות התוך־תאית
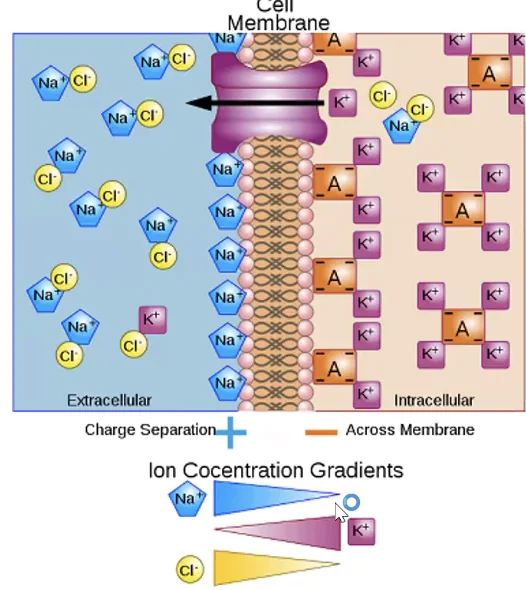
בגלל פיזור היונים נוצר מתח חשמלי על פני הממברנה. בתאי עצב, פוטנציאל המנוחה הוא בקירוב $V_m \approx -70 \text{ mV}$. כלומר, פנים התא שלילי יותר ביחס לחוץ התא. ברקמות אחרות הערך יכול להיות שונה. למשל, בתאי שריר הוא עשוי להיות קרוב יותר ל־$-90 \text{ mV}$.
איך יונים עוברים דרך הממברנה?
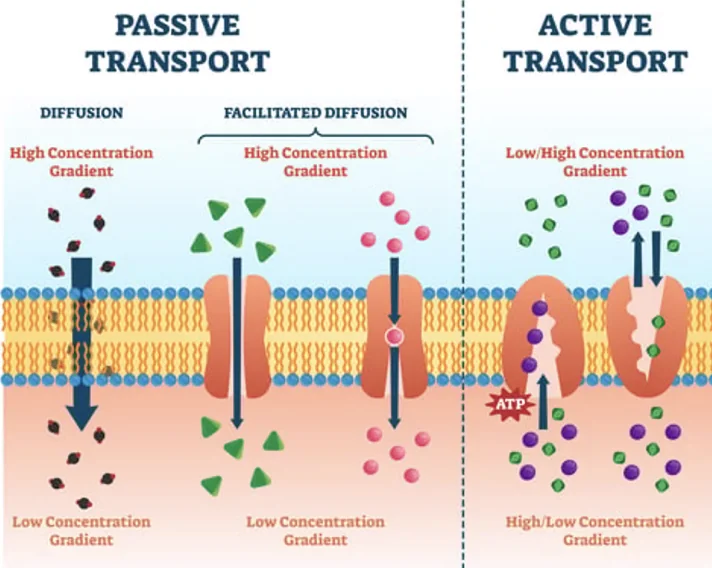
יונים לא חוצים את שכבת הליפידים של הממברנה באופן חופשי, כי הם טעונים חשמלית. הם עוברים בעיקר דרך חלבוני ממברנה (מוזמנים לחזור על ביולוגיה של התא).
1. תעלות סבילות (Leak Channels)
- פתוחות באופן קבוע
- מאפשרות ליונים לעבור לפי הגרדיאנט האלקטרוכימי
- כל תעלה היא ספציפית ליון מסוים
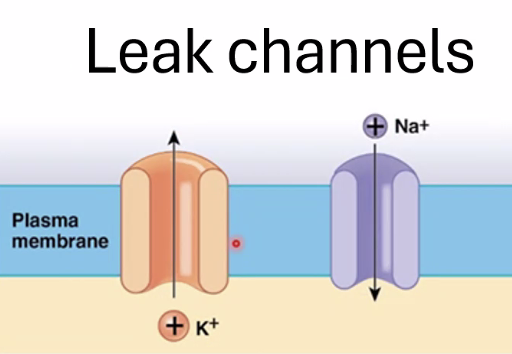
לדוגמה:
- תעלות אשלגן
- תעלות נתרן
- תעלות כלוריד
2. הובלה פעילה - משאבת נתרן-אשלגן
המשאבה החשובה ביותר כאן היא:
משאבת $\ce{Na+/K+ ATPase}$
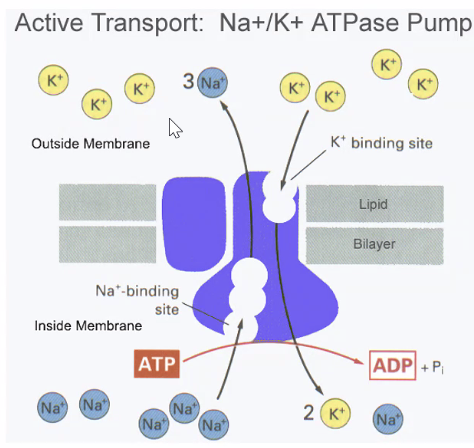
המשאבה:
- מוציאה 3 יוני נתרן $\ce{Na+}$ אל מחוץ לתא
- מכניסה 2 יוני אשלגן $\ce{K+}$ אל תוך התא
- משתמשת ב־ATP
לכן, היא גם תורמת באופן ישיר לכך שפנים התא שלילי יותר.
בנוסף לכך, היא שומרת לאורך זמן על מפלי הריכוזים של נתרן ואשלגן, שבלעדיהם לא ניתן היה לקיים פוטנציאל מנוחה ופוטנציאל פעולה.
בתאי עצב, משאבת $\ce{Na+/K+ ATPase}$ צורכת חלק ניכר מהאנרגיה המטבולית של התא - בערך 40% מהצריכה של הנוירון.
פוטנציאל שיווי משקל (Equilibrium Potential)
לכל יון יש פוטנציאל שיווי משקל משלו. זה המתח החשמלי שבו מאזנים זה את זה:
- הכוח הכימי (שמנסה להזיז את היון לפי מפל הריכוזים)
- ו־הכוח החשמלי (שנובע מהפרש המטענים)
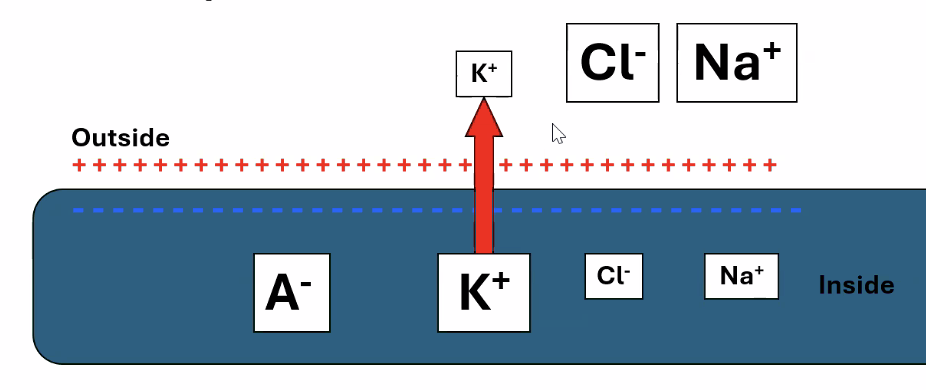
דוגמה: אשלגן
- ריכוז האשלגן גבוה יותר בתוך התא
- לכן הכוח הכימי דוחף אותו החוצה
- אבל פנים התא שלילי, ולכן הכוח החשמלי מושך את האשלגן ($\ce{K+}$) פנימה
כששני הכוחות שווים בגודלם והפוכים בכיוונם מתקבל פוטנציאל שיווי המשקל של אשלגן.
משוואת נרנסט
פוטנציאל שיווי המשקל של יון יחיד מחושב על ידי:
\[E_x = \frac{RT}{zF} \ln \left(\frac{[X]_{out}}{[X]_{in}}\right)\]כאשר:
- $z$ - המטען של היון
- $[X]_{out}$ - ריכוז מחוץ לתא
- $[X]_{in}$ - ריכוז בתוך התא
- $R$ - קבוע הגזים
- $T$ - טמפרטורה בקלווין
- $F$ - קבוע פאראדיי
בטמפרטורת הגוף נהוג להשתמש בקירוב:
\[E_x \approx \frac{61.5}{z} \log \left(\frac{[X]_{out}}{[X]_{in}}\right)\]ערכים אופייניים
\[\begin{aligned} E_{\ce{K}} &\approx -96 \text{ mV} \\ E_{\ce{Na}} &\approx +52 \text{ mV} \\ E_{\ce{Ca}} &\approx +134 \text{ mV} \end{aligned}\]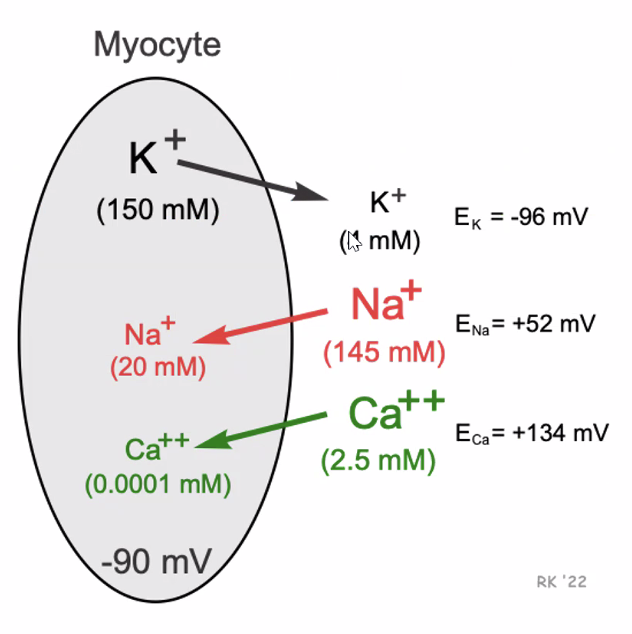
אז למה פוטנציאל המנוחה הוא לא פשוט $E_K$?
כי פוטנציאל המנוחה לא נקבע על ידי יון אחד בלבד. הממברנה חדירה לכמה יונים, וכל אחד מהם תורם לפי:
- מפל הריכוזים שלו
- המטען שלו
- החדירות היחסית של הממברנה אליו (כמה תעלות פתוחות עבור אותו יון - Permeability)
במצב מנוחה, הממברנה חדירה הרבה יותר לאשלגן ($\ce{K+}$) מאשר לנתרן ($\ce{Na+}$), ולכן פוטנציאל המנוחה של הממברנה קרוב לפוטנציאל שיווי המשקל של אשלגן ($E_K$) - אבל לא שווה לו בדיוק.
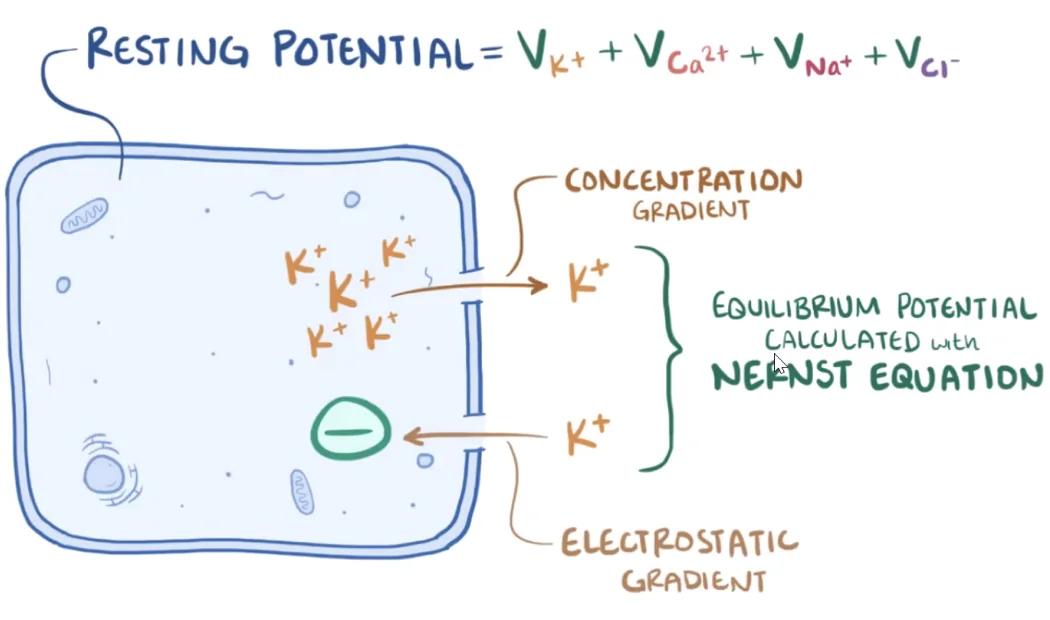
משוואת Goldman-Hodgkin-Katz
כשמתחשבים בכמה יונים יחד, משתמשים עקרונית במשוואת GHK.

הרעיון החשוב מהשיעור:
- פוטנציאל המנוחה נקבע על ידי כמה יונים יחד
- אבל התרומה של כל יון תלויה ב־חדירות הממברנה אליו
- במנוחה, אשלגן הוא הדומיננטי ביותר, משום שיש לו חדירות גבוהה במיוחד
לכן:
\[\boxed{V_m \approx -70 \text{ mV}}\]והוא קרוב יותר ל־$E_K$ מאשר ל־$E_{Na}$.

מה קובע את פוטנציאל המנוחה?
- הבדלי ריכוזי יונים בין פנים התא לחוץ התא
- חדירות הממברנה ליונים שונים
- דליפת אשלגן דרך תעלות סבילות
- דליפה מועטה יותר של נתרן וכלוריד
- פעולת משאבת נתרן-אשלגן
מורפולוגיה בסיסית של תא עצב
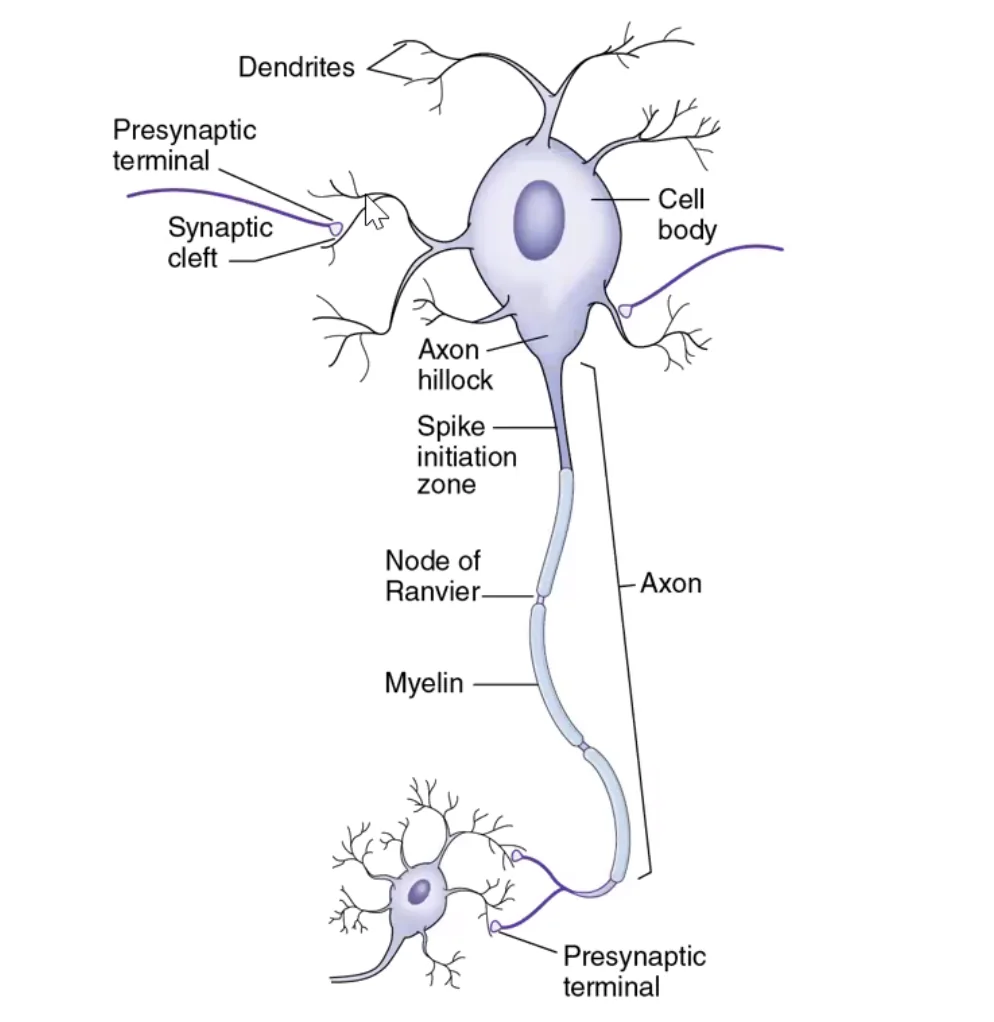
לתא עצב יש שלושה אזורים עיקריים:
- דנדריטים - קולטים מידע מתאים אחרים
- גוף התא (Soma) - מרכז אינטגרציה ועיבוד
- אקסון - מוליך את האות החשמלי לאורך התא
עיקרון חשוב:
- לתא עצב יש בדרך כלל אקסון אחד
- אך יכולים להיות לו דנדריטים רבים
סוגים עיקריים של נוירונים
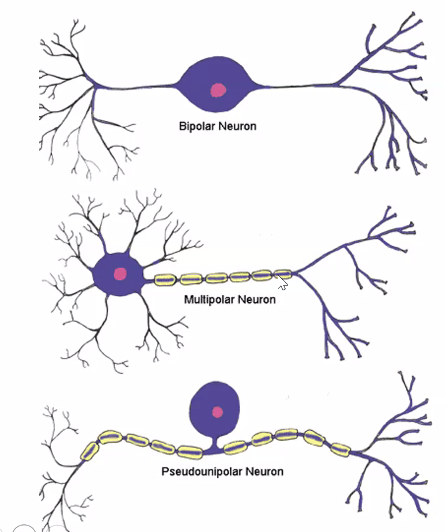
נוירון מולטיפולרי
- אקסון אחד
- הרבה דנדריטים
- הסוג השכיח ביותר במוח
נוירון ביפולרי
- דנדריט אחד ואקסון אחד
נוירון פסאודו־יוניפולרי
- שלוחה אחת שיוצאת מגוף התא ומתפצלת בהמשך
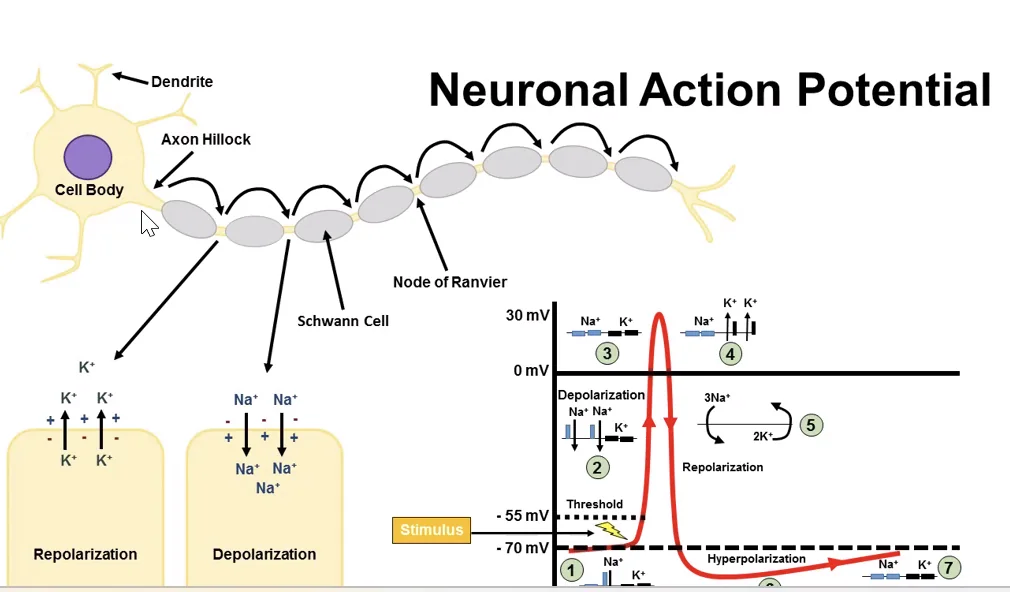
פוטנציאל פעולה (Action Potential)
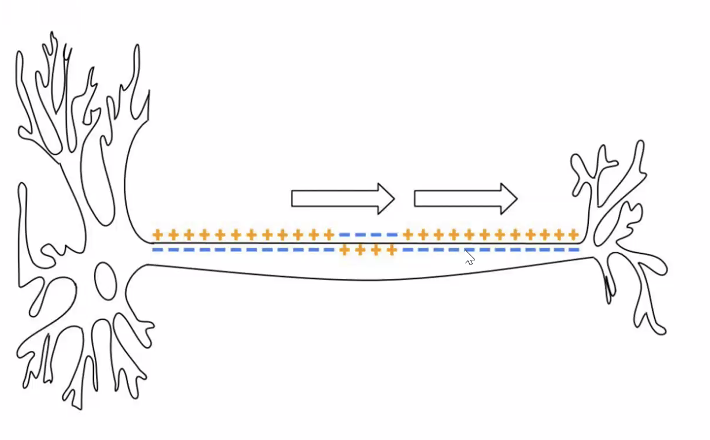
פוטנציאל פעולה הוא שינוי מהיר, חד וחולף במתח על פני הממברנה, שמאפשר הולכת מידע לאורך האקסון.
שלבי פוטנציאל הפעולה
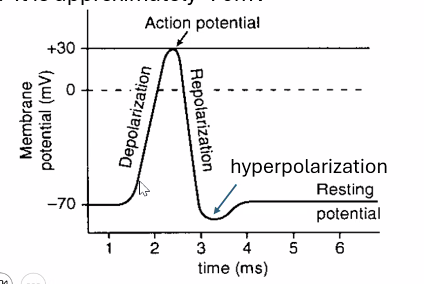 |  |
-
מצב מנוחה - הממברנה נמצאת סביב $\boxed{-70 \text{ mV}}$.
-
הגעה לסף - כשהמתח מגיע לערך סף, בערך $\boxed{-55 \text{ mV}}$, נפתחות תעלות נתרן תלויות מתח.
-
דה־פולריזציה - כניסה מהירה של נתרן לתא:
- תעלות $\ce{Na+}$ תלויות מתח נפתחות
- נתרן נכנס במהירות לתא
- פנים התא נעשה פחות שלילי ואף חיובי
המתח עולה עד בערך $\boxed{+30 \text{ mV}}$.
-
רה־פולריזציה - חסימת נתרן ויציאת אשלגן:
- תעלות הנתרן עוברות אינאקטיבציה
- תעלות אשלגן תלויות מתח נפתחות
- אשלגן יוצא מן התא
- המתח חוזר לערכים שליליים
-
היפר-פולריזציה - לפעמים יציאת האשלגן נמשכת מעט מעבר לנדרש, ולכן המתח נעשה שלילי יותר מפוטנציאל המנוחה.
-
חזרה לפוטנציאל המנוחה - התא חוזר לערך המנוחה הבסיסי (בדרך כלל סביב $-70 \text{ mV}$).
הבהרה: השלבים לעיל מתארים את פוטנציאל הפעולה. אשלגן לא נכנס לתא בפוטנציאל פעולה, להבנתי, אלא בעיקר יוצא ממנו - במיוחד בשלב הרה־פולריזציה וההיפר-פולריזציה. גם במצב מנוחה קיימת בעיקר דליפה של אשלגן החוצה דרך תעלות דליפה, והדבר תורם לפוטנציאל המנוחה. כניסת אשלגן לתא מתרחשת בעיקר דרך משאבת נתרן-אשלגן (ראו לעיל), ולא דרך תעלות תלויות מתח במהלך פוטנציאל הפעולה.
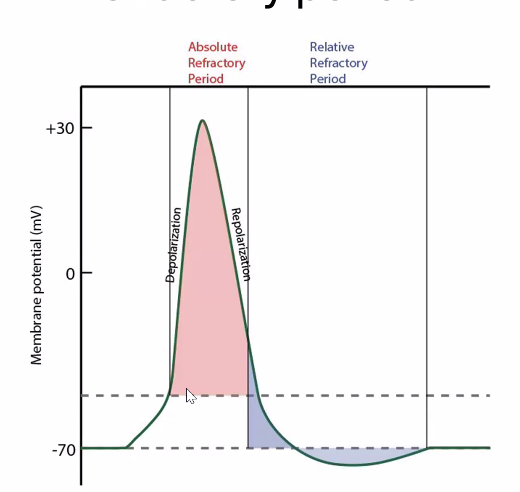
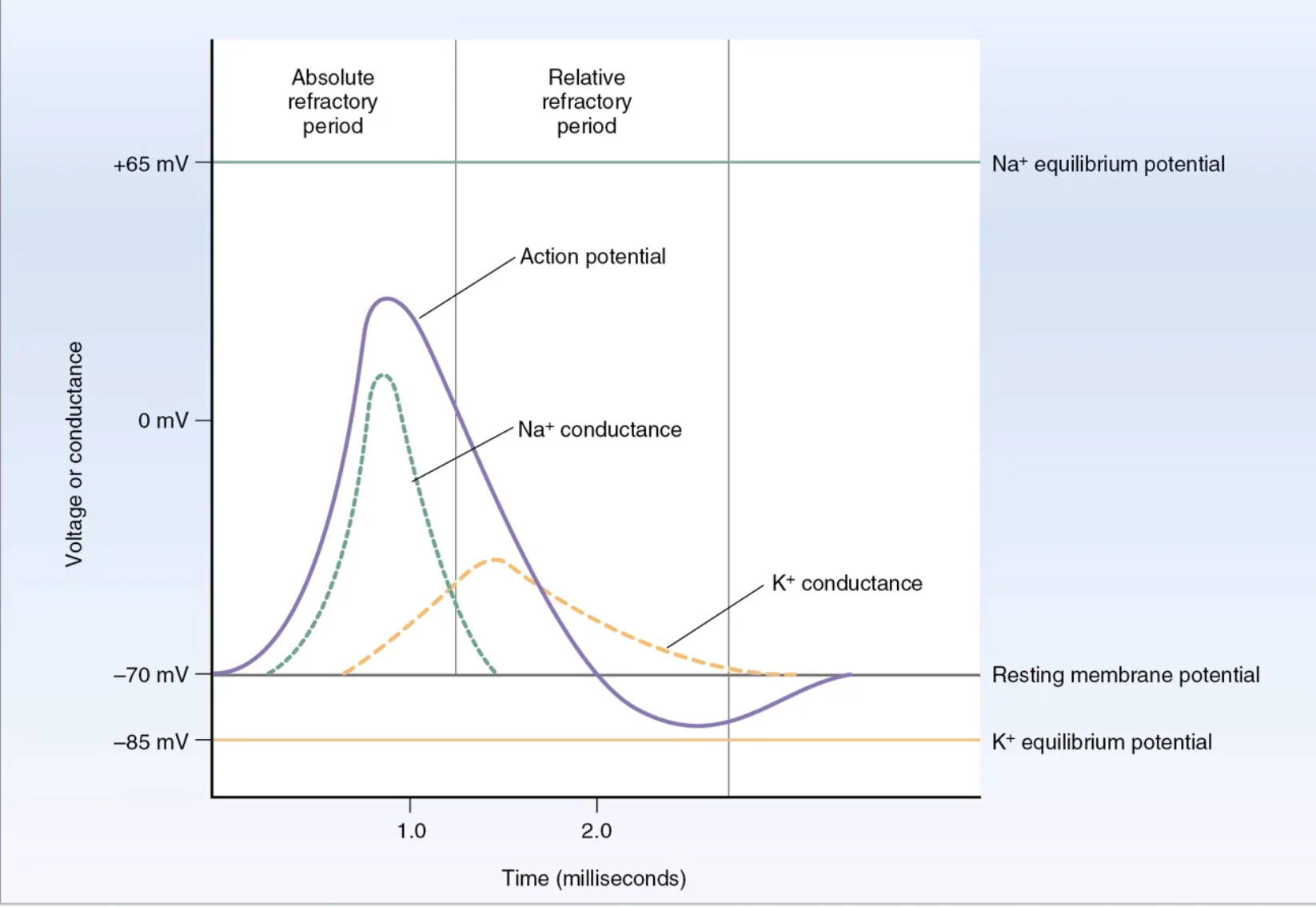
תקופות רפרקטוריות
Absolute Refractory Period
בשלב הזה אי אפשר לייצר פוטנציאל פעולה נוסף. תעלות הנתרן נמצאות במצב אינאקטיבי, ולכן אינן מסוגלות להיפתח מחדש מייד.
Relative Refractory Period
בשלב הזה אפשר, אבל קשה יותר, לייצר פוטנציאל פעולה נוסף. הממברנה עדיין היפר-פולרית יחסית, ולכן נדרש גירוי חזק יותר.
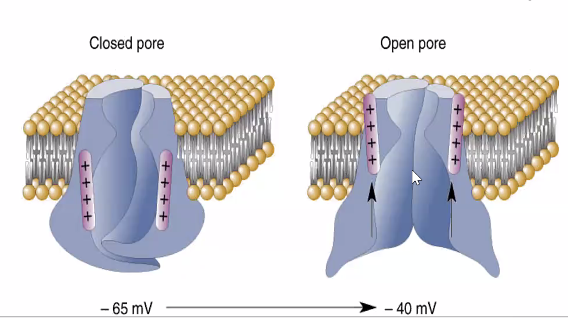
מה פותח את פוטנציאל הפעולה?
השאלה הנכונה היא לא רק ״מה קורה כשהגענו לסף״, אלא גם: איך בכלל הגענו לסף?
הסבר מלא יינתן בהמשך, אבל העיקרון הוא שהמידע שנקלט בדנדריטים ובגוף התא מסוכם באזור תחילת האקסון. האזור החשוב לכך הוא Axon Hillock / Spike Initiation Zone: אם המתח שם מגיע לסף - מתחיל פוטנציאל פעולה.
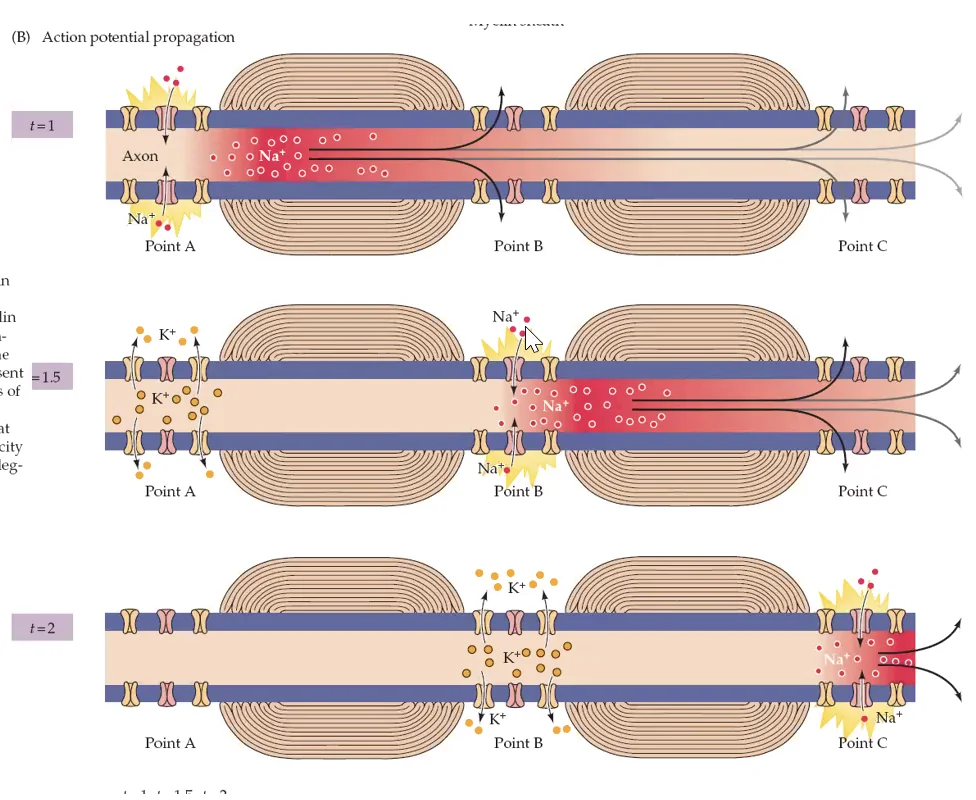
איך פוטנציאל הפעולה מתקדם לאורך האקסון?
אם היינו מסתמכים רק על דיפוזיה פסיבית של יונים, האות היה נחלש עם המרחק. אבל בפועל זה לא קורה. מה שקורה הוא:
- במקטע הראשון נפתחות תעלות נתרן ונכנס $\ce{Na+}$
- הזרם המקומי הזה מעלה את המתח במקטע הסמוך
- כשהמקטע הבא מגיע לסף - גם בו נפתחות תעלות נתרן
- כך מתקבל חידוש רציף של פוטנציאל הפעולה לאורך האקסון
לכן פוטנציאל הפעולה:
- אינו דועך
- נשמר בעוצמה קבועה
- מתקדם באופן אקטיבי לאורך הממברנה
למה ההולכה חד־כיוונית?
כי המקטע שכבר עבר פוטנציאל פעולה נמצא כעת בתקופה רפרקטורית.
לכן:
- הוא לא יכול לעבור מיד דה־פולריזציה נוספת
- האות מתקדם רק קדימה
- אין “פינג-פונג” אחורה
זה אחד הדברים היפים בפיזיולוגיה: לא צריך “חץ” שיגיד לאן להתקדם. הכיווניות נוצרת מתוך המנגנון עצמו.
מיאלין והולכה סלטטורית
ברוב האקסונים קיימת מעטפת מיאלין:
- מבודד את האקסון חשמלית
- מקטין זליגת זרם
- מאפשר הולכה מהירה יותר
בין מקטעי המיאלין יש אזורים קצרים חשופים בשם Nodes of Ranvier, שמרוכזות בהם תעלות יונים תלויות מתח. לכן, פוטנציאל הפעולה לא מתקדם באופן רציף לאורך כל הממברנה, אלא “קופץ” מצומת לצומת. לתופעה הזאת קוראים הולכה סלטטורית (Saltatory Conduction).
למה מיאלין כל כך חשוב?
-
מהירות: האות מתקדם הרבה יותר מהר.
-
חיסכון אנרגטי: אין צורך להפעיל תעלות ומשאבות על פני כל האקסון, אלא בעיקר בצמתים.
-
יעילות ביולוגית: אות מהיר יותר בפחות עלות מטבולית.
מה קורה כשיש פגיעה במיאלין?
כשהמיאלין נפגע, הולכת האות נעשית איטית ופחות יעילה. הסיגנל מתקדם לאט יותר, ולעיתים אפילו נפגע או נכשל. זה בסיס חשוב להבנה של מחלות דה־מיאלינטיביות.
סיכום
-
פוטנציאל מנוחה: נובע מהבדלי ריכוזי יונים, חדירות סלקטיבית של הממברנה, ומשאבת נתרן-אשלגן.
-
פוטנציאל שיווי משקל: מתאר יון יחיד, ומחושב בעזרת נרנסט.
-
פוטנציאל מנוחה אמיתי: נקבע על ידי כמה יונים יחד, ובעיקר לפי החדירות היחסית שלהם.
-
פוטנציאל פעולה: מתחיל בהגעה לסף, כולל דה־פולריזציה, רה־פולריזציה והיפר-פולריזציה.
-
דה־פולריזציה - בעיקר כניסת נתרן.
-
רה־פולריזציה - בעיקר יציאת אשלגן.
-
חד־כיווניות - נובעת מהתקופה הרפרקטורית.
-
מיאלין - מאפשר הולכה סלטטורית מהירה וחסכונית.
טעויות שלא כדאי לעשות
- לבלבל בין פוטנציאל מנוחה לבין פוטנציאל פעולה
- לחשוב שפוטנציאל המנוחה שווה בדיוק ל־$E_K$
- לחשוב שמשוואת נרנסט מתארת את כל הממברנה - היא מתארת יון אחד בלבד
- לשכוח שהתרומה של כל יון תלויה בחדירות הממברנה אליו
- להתבלבל בין פתיחת תעלות נתרן לבין פתיחת תעלות אשלגן בשלבים השונים של פוטנציאל הפעולה
בהקשר אחר, כתבתי פעם על מודל חישובי של נוירונים במסגרת תואר במדעי המחשב. מוזמנים לקרוא באתר האישי (האחר) שלי.
דור פסקל