פתיחה - מחיידקים לאאוקריוטים
בשיעור הקודם עסקנו בחיידקים. נזכיר שבחיידק יש RNA polymerase מרכזי אחד, ו־sigma factors שונים מכוונים אותו לפרומוטורים המתאימים לצורכי התא. באאוקריוטים החלוקה מורכבת יותר: בגרעין פועלים שלושה RNA polymerases מרכזיים, שכל אחד מהם מתמחה בקבוצת גנים אחרת.
| פולימראז | תפקיד עיקרי |
|---|---|
| RNA polymerase I | מסנתז את רוב ה־rRNA |
| RNA polymerase II | מסנתז בעיקר mRNA של גנים המקודדים לחלבון |
| RNA polymerase III | מסנתז בעיקר tRNA וגם 5S rRNA |
השיעור מתמקד ב־RNA polymerase II (Pol II), מפני שהוא הפולימראז האחראי על שעתוק הגנים המקודדים לחלבון ועל כן נתון לבקרה רחבה מאוד.
RNA Polymerase II
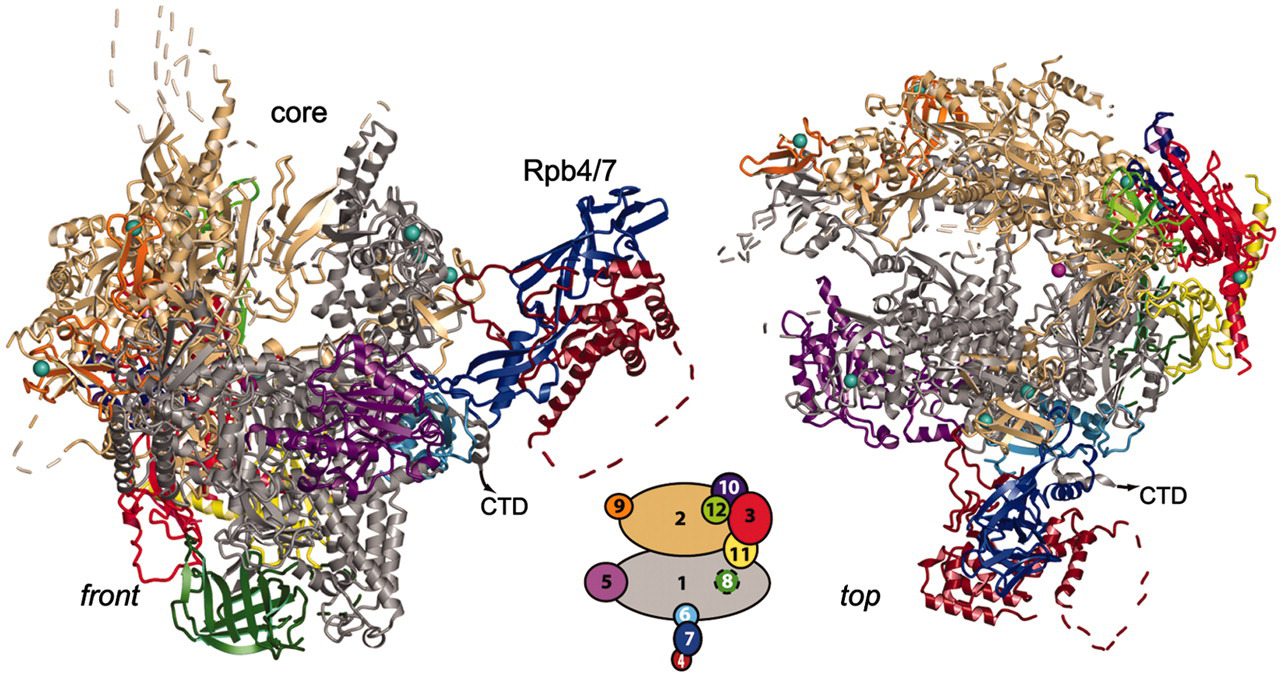
Pol II הוא קומפלקס המורכב מ־12 תת־יחידות, הנקראות Rpb1–Rpb12.
- עשר תת־יחידות, דומות לתת־יחידות המקבילות ב־Pol I וב־Pol III, ויוצרות את הליבה המבנית והקטליטית של הפולימראז.
- Rpb4 ו־Rpb7 יוצרות יחד את ה־stalk (גבעול) - אזור המאפשר אינטראקציות עם חלבונים נוספים המלווים את Pol II.
Rpb1, Rpb2 וה־active-site cleft
Rpb1 היא תת־היחידה הגדולה ביותר. היא מכילה חלק מרכזי מהאזור הקטליטי וגם את זנב ה־CTD, הייחודי ל־Pol II.
Rpb1 ו־Rpb2 יוצרות ביניהן את ה־cleft: סדק טעון חיובית שאליו נכנס ה־DNA הטעון שלילית. המבנה הכללי מזכיר פה של פקמן, כאשר ה־DNA נכנס מהצד ה־downstream אל האתר הפעיל.
Rpb1 יוצרת גם את ה־clamp (צבט), שעוטף את ה־DNA ומחזיק אותו בתוך הפולימראז:
- במהלך
initiationה־clamp פתוח יחסית ומאפשר ל־DNA להיכנס. - כאשר Pol II עובר ל־
elongation, ה־clamp נסגר ומייצב את קומפלקס השעתוק.
בקדמת ה־clamp נמצא ה־shelf, המסייע לכוון את ה־DNA אל תוך הפולימראז.
אזורים מבניים חשובים
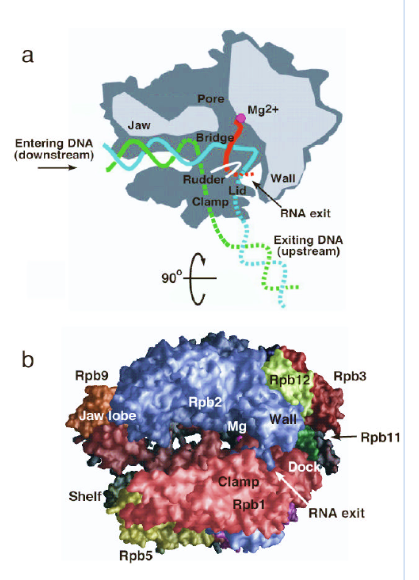
- Cleft - הסדק שאליו נכנס ה־DNA ובתוכו ממוקם האתר הפעיל.
- Clamp - עוטף את ה־DNA הנכנס ומייצב אותו בתוך Pol II.
- Shelf ו־jaw - מכוונים ומייצבים את ה־DNA בדרכו אל האתר הפעיל.
- Bridge helix - פועל בדומה ל־ratchet ומסייע למשוך את ה־DNA פנימה במהלך התקדמות הפולימראז.
- Rudder ו־lid - מסייעים לשמור על בועת השעתוק ולהפריד בין ה־RNA החדש לבין גדיל התבנית.
- Wall - גורם ל־DNA לשנות כיוון בזווית חדה ומסייע בהפרדת ה־RNA מה־DNA.
- RNA exit channel - התעלה שדרכה יוצא ה־RNA החדש.
- Funnel / pore - המסלול שדרכו נכנסים ה־NTPs אל האתר הפעיל.
- Stalk - Rpb4/7 - אזור קישור לחלבונים שבאים באינטראקציה עם Pol II.
CTD - זנב Pol II שמנהל את מחזור השעתוק
ל־Rpb1 יש זנב ארוך ולא מקופל בשם CTD (C-terminal domain). הזנב משמש כפלטפורמת קישור דינמית: דפוסי פוספורילציה שונים מאפשרים לחלבונים מסוימים להיקשר אליו, לאחרים להתנתק, ול־Pol II לעבור משלב אחד של השעתוק לשלב הבא.
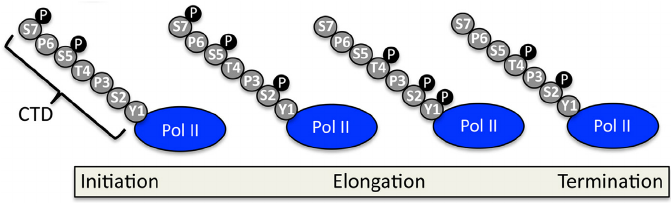
מבנה ה־CTD
ה־CTD בנוי מחזרות של רצף בן שבע חומצות אמינו:
Tyr1-Ser2-Pro3-Thr4-Ser5-Pro6-Ser7
Y S P T S P S
ב־Pol II של אדם יש 52 חזרות. הפוספורילציה משתנה בהדרגה לאורך הגן; יש להתייחס אליה כאל גרדיאנט של סימנים, ולא כאל מתג יחיד.
קוד הפוספורילציה לאורך הגן
| שלב | דפוס הפוספורילציה | משמעות |
|---|---|---|
| לפני initiation | ה־CTD כמעט אינו מזורחן | מאפשר הרכבת קומפלקס ההתחלה באזור הפרומוטור |
| מעבר מ־initiation ל־elongation | עולה הפוספורילציה של Ser5 ושל Ser7 | מאפשרת שינוי בהרכב החלבונים ויציאה של Pol II מאזור ההתחלה |
| תחילת elongation | מתווספת פוספורילציה של Ser2 | מסמנת מעבר להתארכות ומסייעת בגיוס חלבוני עיבוד RNA |
| לאורך גוף הגן | Ser5-P ו־Ser7-P יורדות בהדרגה; Ser2-P נשמרת ועולה גם Tyr1-P | חלבונים שונים נקשרים בהתאם למיקום Pol II לאורך הגן |
| לקראת סוף הגן | בערך 180bp לפני הסיום מתרחשת דה־פוספורילציה של Tyr1, בעוד Ser2-P נשמרת | מאפשרת קישור של פקטורי טרמינציה |
Thr4-P וגם Ser2-P קשורות לגיוס חלבוני splicing בזמן שה־RNA עדיין מסונתז.
פרומוטורים של Pol II
ה־promoter הוא אזור DNA המזוהה על ידי פקטורי שעתוק, וממקם את Pol II בתחילת הגן. נקודת ההתחלה של השעתוק נקראת TSS (Transcription Start Site) ומסומנת כ־+1:
- עמדות לפני ה־TSS הן
upstreamומסומנות במספרים שליליים. - עמדות בתוך הגן הן
downstreamומסומנות במספרים חיוביים.
הפרומוטור אינו רצף יחיד וקבוע. גנים שונים משתמשים בשילובים שונים של אלמנטים, ולעיתים חלק מהאלמנטים חופפים זה לזה.
Core promoter elements
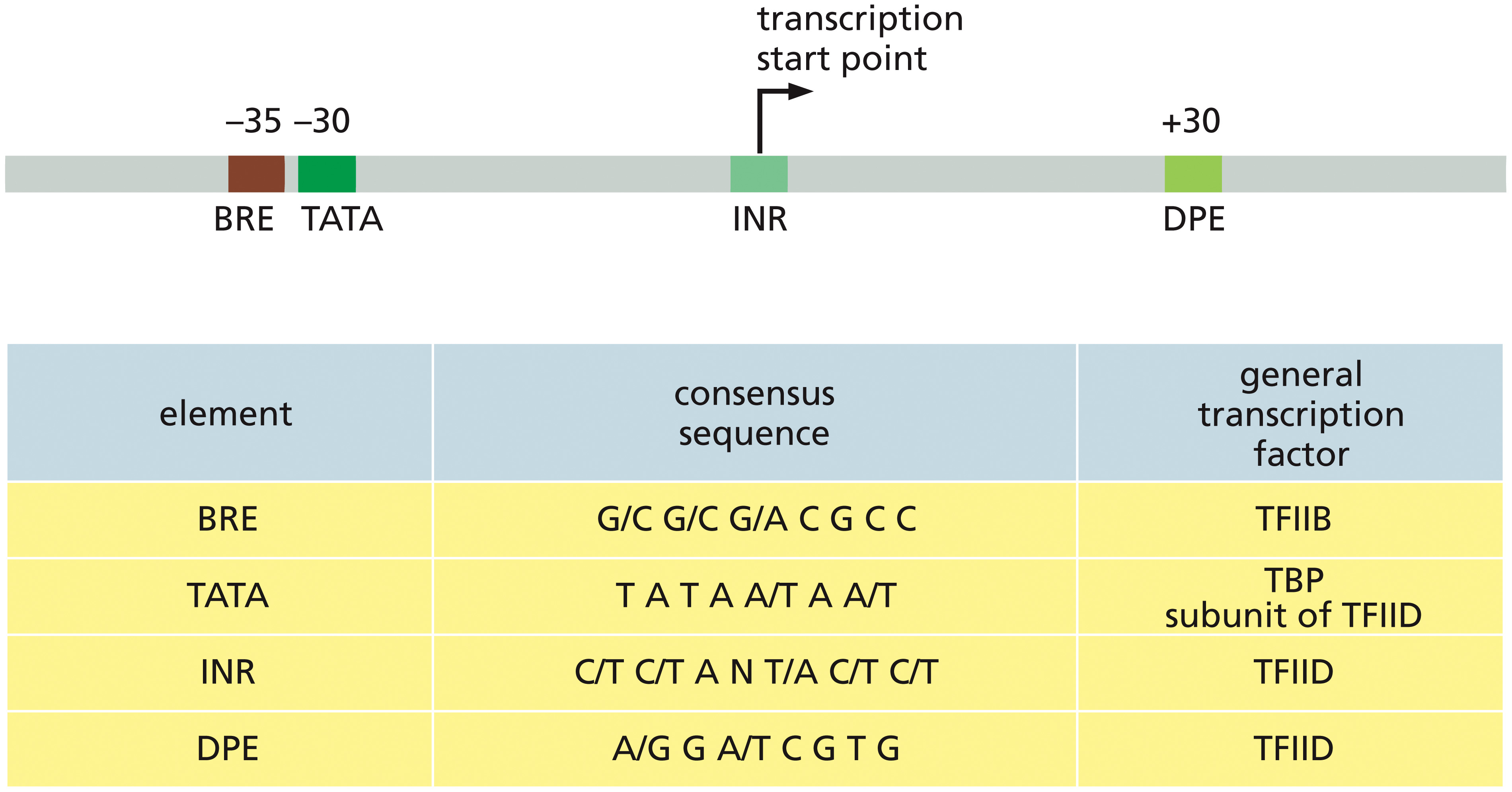
ה־core promoter הוא האזור הקרוב ביותר ל־TSS, שבו מורכב ה־PIC (Pre-Initiation Complex).
האלמנטים המרכזיים הם:
- TATA box - מזוהה על ידי TBP (
TATA-binding protein). - BRE (
TFIIB Recognition Element) - מזוהה על ידי TFIIB. - Initiator (Inr) - נמצא סביב
+1. - DPE (
Downstream Promoter Element) - נמצא בתוך הגן ופועל יחד עם Inr.
לא כל פרומוטור מכיל את כל האלמנטים.
BRE - TATA - Inr (+1) - DPE
TATA box
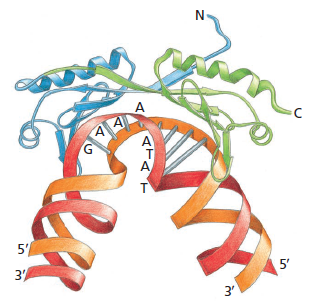
ה־TATA box הוא רצף עשיר ב־A וב־T, הנמצא בדרך כלל סביב -25 עד -30 ביחס ל־TSS. הוא מזוהה על ידי TBP, תת־יחידה של TFIID.
TBP נקשר ל־TATA box ומכופף את ה־DNA. בשיעור הודגש שהכיפוף חושף את אזור הפרומוטור ומקל על קישור יתר פקטורי השעתוק.
למרות שה־TATA box הוא האלמנט המוכר ביותר, הוא קיים רק בכ־30%–20% מהגנים. ברוב הגנים אין TATA box קלאסי, ורבים מהם נשענים על Initiator ואלמנטים נוספים.
BRE
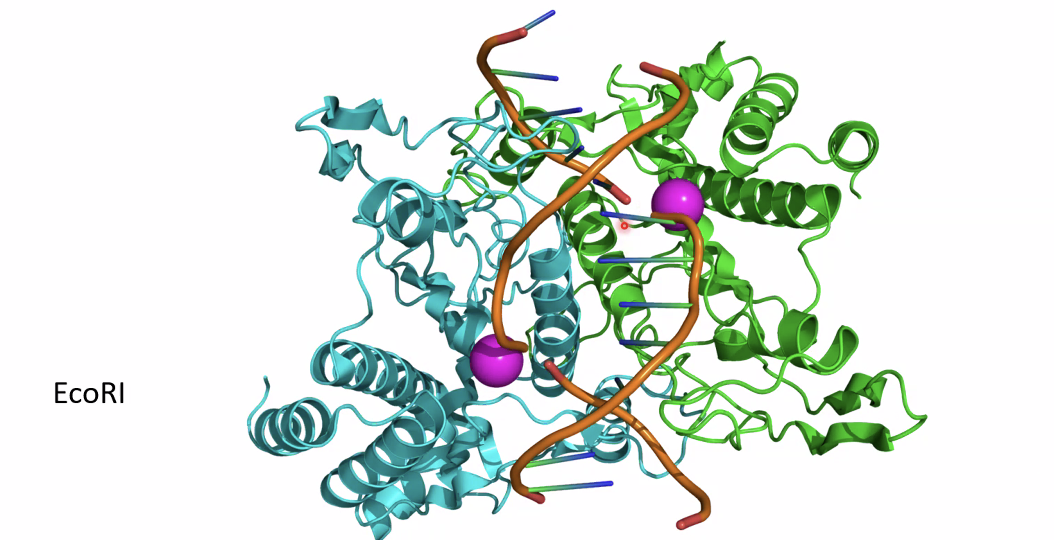
BRE הוא רצף DNA המזוהה על ידי TFIIB.
במצב הקלאסי, סביב TATA box יכולים להופיע שני אלמנטים:
- BREu - בצד ה־upstream ביחס ל־TATA box.
- BREd - בצד ה־downstream ביחס ל־TATA box.
ל־TFIIB יש שני אזורי קישור ל־DNA, ולכן הוא יכול ליצור נקודות מגע משני צדי ה־TATA box. השמות BREu ו־BREd ניתנו משום שהאלמנטים התגלו בתחילה סביב TATA box, אך BRE ו־TATA אינם חייבים להופיע יחד: אפשר למצוא BRE ללא TATA, וגם TATA ללא BRE.
רצפי קונצנזוס וקוד IUPAC
אלמנטי הפרומוטור הם רצפי קונצנזוס: הם מתארים דפוס מועדף של בסיסים, ולא רצף זהה לחלוטין בכל גן. כאשר יותר מבסיס אחד אפשרי באותה עמדה, משתמשים בקוד IUPAC.
| קוד | משמעות | קוד | משמעות |
|---|---|---|---|
Y | פירימידין: C או T | R | פורין: A או G |
W | A או T, קשרים חלשים יותר | S | G או C, קשרים חזקים יותר |
M | A או C | K | G או T |
B | כל בסיס חוץ מ־A | D | כל בסיס חוץ מ־C |
H | כל בסיס חוץ מ־G | V | כל בסיס חוץ מ־T |
N | כל בסיס |
פקטורי השעתוק מזהים את דפוס הקונצנזוס המתאים גם כאשר הרצף המדויק משתנה מעט בין גנים.
Initiator - Inr
ה־Initiator נמצא סביב ה־TSS. בשיעור הודגש שזהו אלמנט מרכזי מאוד: גנים רבים יכולים להסתפק בו גם ללא TATA box.
DPE
DPE נמצא בתוך האזור המשועתק, סביב +28 עד +32. הוא מסייע לייצב את ה־TFIID על הפרומוטור יחד עם ה־Initiator.
- DPE פועל יחד עם Inr ואינו מופיע כאלמנט עצמאי.
- Inr יכול להופיע גם ללא DPE.
Proximal promoter elements
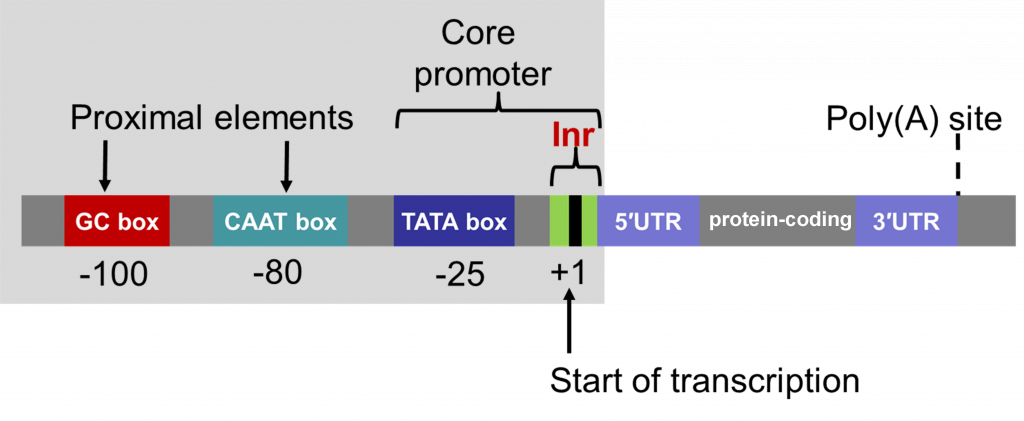
מעבר ל־core promoter, קיימים proximal promoter elements. בשיעור הם תוארו כרצפים קרובים יחסית, בערך מהאזור של -100 ועד סדר גודל של -1,000bp.
אלה עדיין אלמנטים של הפרומוטור הקרוב, ולא enhancers. נקשרים אליהם פקטורי שעתוק ספציפיים, שמסייעים לגיוס או לייצוב של מכונת השעתוק באזור ה־core promoter.
CAAT box
- רצף קונצנזוס שכיח:
CCAAT. - יכול להופיע בכמה עותקים ובשילובים שונים עם TATA box או Inr.
- נקשר בין היתר לפקטור הספציפי
NF-Y. - מסייע לגייס את פקטורי השעתוק הכלליים אל הפרומוטור.
GC box
- רצף עשיר ב־G וב־C, לדוגמה
GGGCGG. - יכול להופיע במספר רב של עותקים לפני ה־core promoter.
- נקשר לפקטור השעתוק הספציפי
Sp1, שיכול לסייע בגיוס TFIID. - נפוץ בפרומוטורים של housekeeping genes, וגם בגנים התפתחותיים ובגנים בעלי ביטוי תלוי-רקמה.
CpG islands
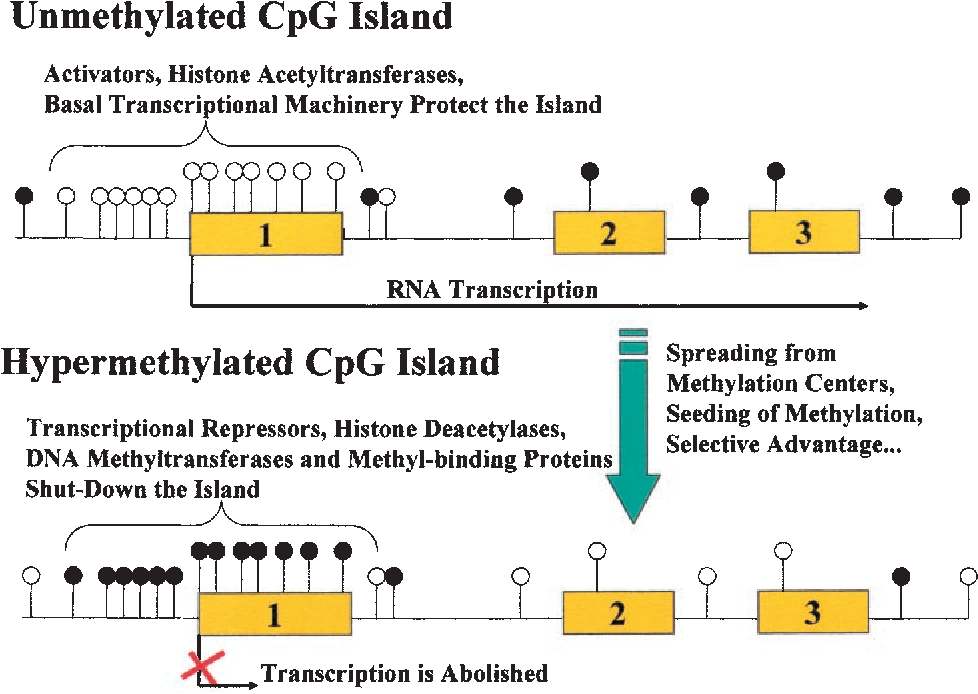
CpG islands הם אזורים ארוכים ועשירים ב־CpG; באורך של כ־1,100bp. בגלל תכולת ה־GC הגבוהה, הם עשויים להכיל רצפים רבים הדומים ל־GC boxes ולהשתתף בבקרת השעתוק.
- כאשר הציטוזינים באזור אינם ממותלים, הכרומטין נוטה להיות פתוח יותר והגן יכול להתבטא.
- מתילציה של ציטוזין ל־5-methylcytosine קשורה לדחיסת האזור להטרוכרומטין, ולהפחתה או מניעה של התחלת השעתוק.
GTFs והרכבת ה־PIC
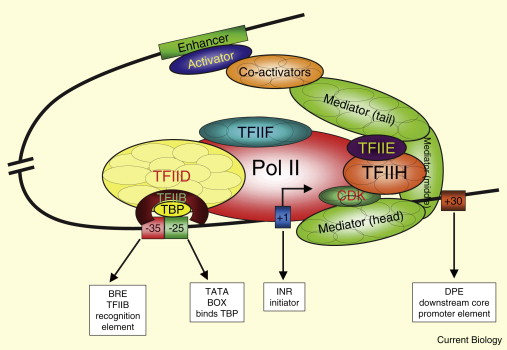
Pol II אינו מזהה את הפרומוטור לבדו. הוא זקוק ל־GTFs (General Transcription Factors), אשר נקשרים ל־core promoter ומרכיבים יחד עם Pol II את ה־PIC (Pre-Initiation Complex).
מבחינה רעיונית, אוסף ה־GTFs מבצע באאוקריוטים חלק מהתפקיד שמבצע sigma factor בחיידקים: זיהוי אזור ההתחלה, גיוס הפולימראז, ומיקומו ביחס ל־TSS. ההבדל הוא שבאאוקריוטים נדרשים כמה קומפלקסים חלבוניים, ולא פקטור יחיד.
מקור השם TFII: TF עבור Transcription Factor, וה־II מציין את Pol II. האות שלאחר מכן ספציפית.
חשוב להבדיל בין:
- GTFs - פקטורים כלליים המשתתפים בהרכבת PIC בפרומוטורים רבים. יכולים עקרונית להיקשר לכל אזורי הפרומוטר הפשוטים.
- פקטורי שעתוק ספציפיים - מזהים רצפים מסוימים, למשל
Sp1ב־GC box אוNF-Yב־CAAT box, ומבקרים קבוצות מסוימות של גנים.
TFIID, TBP ו־TAFs
TFIID הוא קומפלקס של 12 תת־יחידות:
- תת־יחידה אחת היא TBP, שנקשרת ל־TATA box ומכופפת את ה־DNA.
- 11 תת־היחידות האחרות נקראות TAFs (
TBP-associated factors).
השם “TAFs” הוא היסטורי: בפועל, TAFs יכולות לבצע אינטראקציות נוספות מלבד תמיכה ב־TBP.
סדר הרכבת ה־PIC במצב הקלאסי
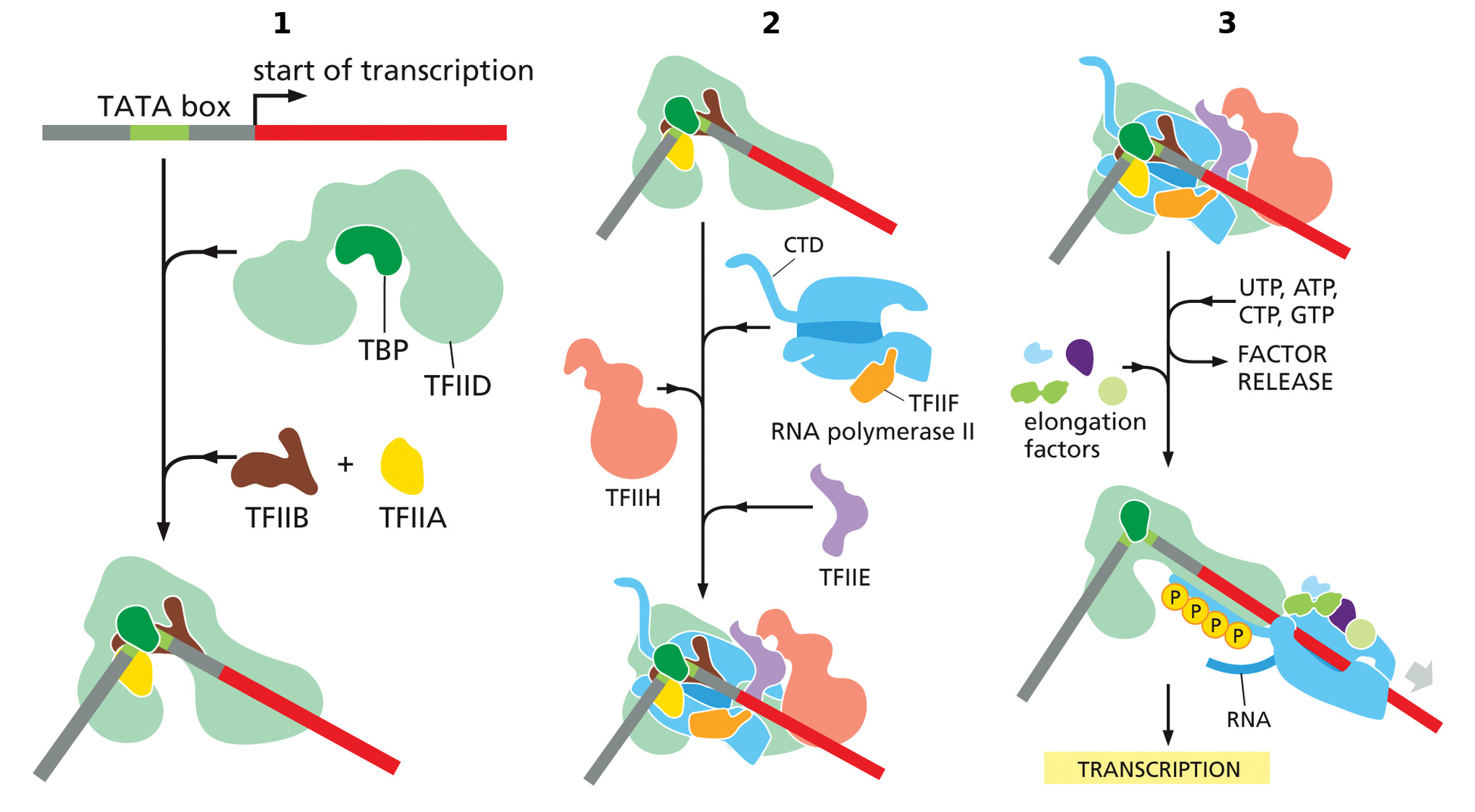
- TFIID נקשר לפרומוטור; בפרומוטור המכיל TATA, TBP נקשר ל־TATA box ומכופף את ה־DNA.
- TFIIB נקשר ל־TFIID ול־BRE. TFIIA יכול להצטרף ולייצב את הקומפלקס, אך הוא אינו תמיד נוכח.
- רק לאחר הרכבת הקומפלקס המוקדם, מגויס Pol II, שמגיע כשהוא קשור ל־TFIIF.
- TFIIE ו־TFIIH מצטרפים אחרונים ומשלימים את ה־PIC.
- TFIIH מזרחן את ה־CTD, הרכב החלבונים סביב Pol II משתנה והפולימראז יכול להתחיל לצאת מהפרומוטור.
העיקר בסדר ההרכבה: Pol II אינו מגיע ראשון. TFIID ו־TFIIB מסמנים ומארגנים את אזור הפרומוטור; Pol II–TFIIF מגיעים אחריהם; TFIIE ו־TFIIH משלימים את הקומפלקס.
תפקידי ה־GTFs
| פקטור | תפקיד | מנמוניקה |
|---|---|---|
| TFIID | הקומפלקס הגדול ביותר; מתחיל את ההרכבה; מכיל TBP ו־11 TAFs | D - Dock הרציף שעליו מתחילה הרכבת הקומפלקס. |
| TFIIB | נקשר ל־TFIID ולרצפי BRE, ממקם את הקומפלקס ומסייע בגיוס Pol II | B - Bridge/BRE-binding יוצר גשר בין TFIID ל־Pol II וממקם את הפולימראז. |
| TFIIA | מייצב את הקומפלקס המוקדם; אינו נדרש בכל תהליך שעתוק | A - Anchor עוגן; מייצב את הקומפלקס אך לא תמיד הכרחי. |
| TFIIF | נקשר מראש ל־Pol II ומגיע איתו ל־PIC | F - Ferry ה״מעבורת״ שמביאה את Pol II לפרומוטור. |
| TFIIE | מצטרף לאחר גיוס Pol II יחד עם TFIIH | E - Escort מגייס ומכוון את H |
| TFIIH | מזרחן את ה־CTD ומאפשר ל־Pol II לצאת משלב האיתחול, לשלב ההתארכות | H - Helix מסייע בפתיחת ה־DNA |
מדיאטורים ואלמנטים רגולטוריים רחוקים
ה־GTFs ממקמים את Pol II בפרומוטור, אך אינם מסבירים כשלעצמם מתי גן מסוים יופעל או יעוכב. הבקרה נעשית גם בעזרת פקטורי שעתוק ספציפיים, ואלמנטים רגולטוריים, שיכולים להימצא אלפי בסיסים מהגן.
להלן נסקור בקצרה ארבעה סוגים:
- Mediator - קומפלקס חלבוני שמקשר בין פקטורי שעתוק ספציפיים לבין Pol II.
- Enhancers - רצפי DNA שאליהם נקשרים אקטיבטורים, שמגבירים את השעתוק.
- Silencers - רצפי DNA שאליהם נקשרים רפרסורים, שמפחיתים את השעתוק.
- Insulators - רצפי DNA שיוצרים גבול פיזי בין אזורים רגולטוריים, ומונעים אינטראקציות לא רצויות.
- Enhancer, silencer ו־insulator הם רצפי DNA.
- אקטיבטורים, רפרסורים ו־CTCF הם חלבונים.
Mediator
Mediator הוא קומפלקס גדול של כ־30 תת־יחידות. הוא מקשר בין חלבונים רגולטוריים, הקשורים לאזורים רחוקים, לבין Pol II וה־PIC בפרומוטור.
למדיאטור יש שלושה חלקים עיקריים:
- ראש - יוצר אינטראקציות עם Pol II ועם אזור הפרומוטור.
- גוף - מחבר בין חלקי הקומפלקס.
- זנב - יוצר אינטראקציות עם חלבונים רגולטוריים הקשורים לאלמנטים רחוקים.
לולאת DNA מקרבת את האלמנט הרחוק אל הפרומוטור. הלולאה עצמה אינה קובעת אם השעתוק יופעל או יעוכב; זה נקבע לפי הרצף הרגולטורי והחלבונים הקשורים אליו. דרך המדיאטור, הם יכולים לקדם את פעילות Pol II או למנוע את גיוסו ופעילותו.
Enhancers
Enhancers הם רצפי DNA שאליהם נקשרים אקטיבטורים.
- הם יכולים להימצא אלפי בסיסים מהגן.
- אקטיבטורים מגייסים חלבונים נוספים ויוצרים אינטראקציות עם Mediator.
- קיפול ה־DNA מקרב את ה־enhancer לפרומוטור, ומאפשר לו להגביר את השעתוק.
Silencers
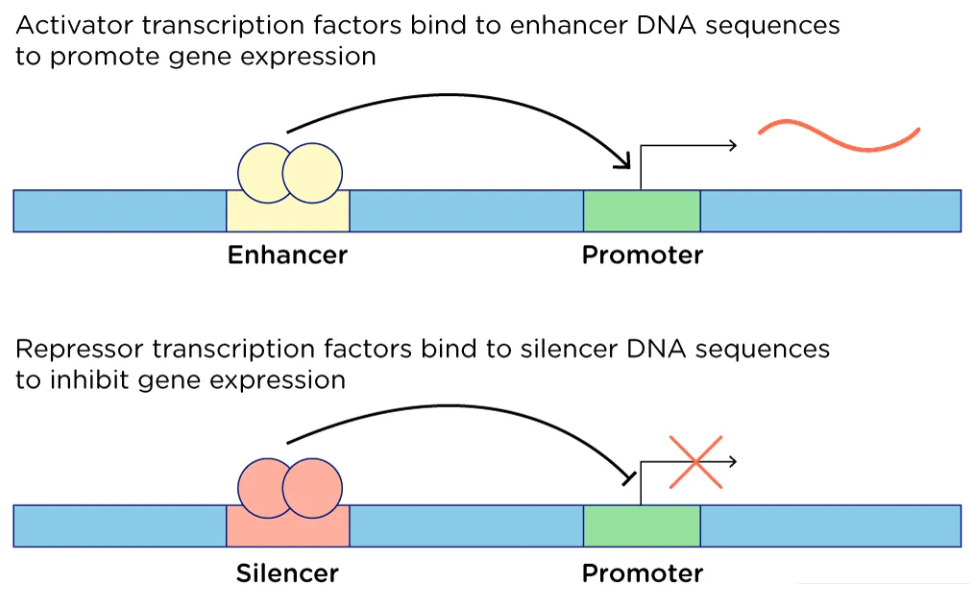
Silencers הם רצפים שאליהם נקשרים רפרסורים. באמצעות לולאות ואינטראקציות עם קומפלקסים רגולטוריים, הם יכולים להפריע להרכבת מכונת השעתוק או להפחית את פעילותה.
Insulators ו־CTCF
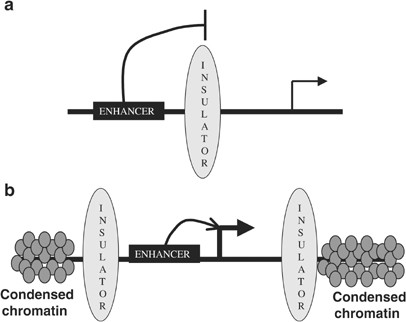
Insulators הם רצפי DNA היוצרים גבול פיזי בין אזורים רגולטוריים. אחד החלבונים המרכזיים שנקשרים אליהם הוא CTCF, שלעיתים פועל יחד עם קוהזין ומשתתף ביצירת לולאות וגבולות של TADs.
שתי פעולות עיקריות שהוזכרו:
- חסימת Enhancer - insulator הנמצא בין enhancer לבין פרומוטור יכול למנוע פיזית יצירת מגע ביניהם.
- Barrier activity - insulator יכול לעצור התפשטות של הטרוכרומטין וסימני דיכוי אל אזור בגן שצריך להישאר פתוח.
לעיתים התפקיד של CTCF הוא לארגן את המרחב הגרעיני, כך שאלמנט רגולטורי ישפיע רק על התחום המתאים. הוא לא חייב לדכא את הפרומוטור ישירות.
שלבי השעתוק של RNA Polymerase II
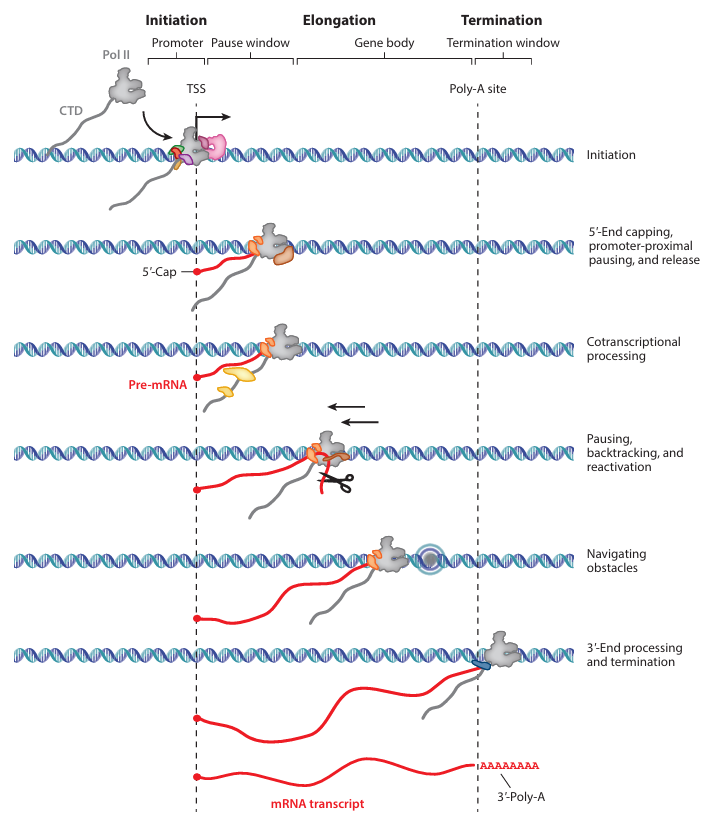
לשעתוק על ידי Pol II יש שלושה שלבים כלליים:
- Initiation - הרכבת PIC, התחלת סינתזת RNA ויציאה מהפרומוטור.
- Elongation - התקדמות לאורך הגן, עצירות בקרה ועיבוד ה־RNA תוך כדי סינתזה.
- Termination - חיתוך קצה ה־RNA, הוספת poly-A ושחרור Pol II מה־DNA.
1. איתחול (Initiation) ו־Promoter Escape
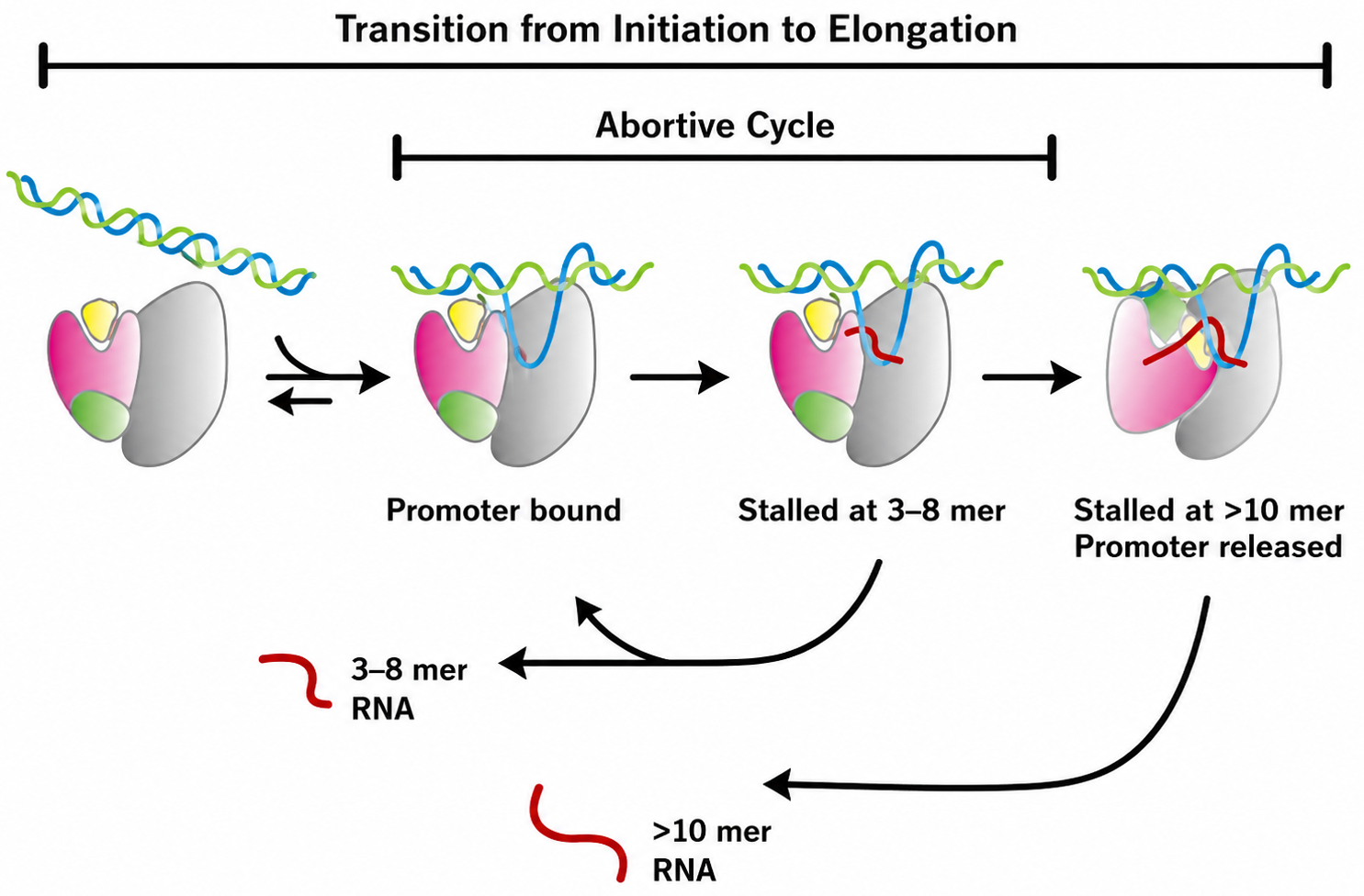
לאחר הרכבת ה־PIC, Pol II מתחיל למשוך את ה־DNA אל האתר הפעיל ולסנתז RNA קצר. בתחילת התהליך הפולימראז עדיין קשור בחוזקה לפרומוטור ולפקטורי ההתחלה, ולכן הוא עובר מספר מחזורים של משיכה וסינתזה לפני שהוא מצליח להתרחק מהפרומוטור.
כאשר התעתיק מגיע לאורך של יותר מכ־10 nt, פוספורילציות על ה־CTD ושינוי באינטראקציות עם ה־GTFs מאפשרים promoter escape.
במהלך היציאה:
- TFIIE ו־TFIIH משתחררים מ־Pol II.
- TFIIF נשאר עם Pol II ומלווה אותו לתחילת ההתארכות.
- TFIID ו־TFIIB יכולים להישאר על הפרומוטור.
הישארות TFIID ו־TFIIB יוצרת מעין scaffold שמאפשר לגייס Pol II נוסף במהירות ולבצע סבב שעתוק נוסף, כאשר התא זקוק לכך.
האחרונים שהגיעו (E ו־H), הם הראשונים לעזוב.
1.5. Promoter-proximal pausing
גם לאחר promoter escape, Pol II אינו נכנס מיד להתארכות רציפה. לאחר סינתזה של בערך 30 nt, הוא נעצר סמוך לפרומוטור. העצירה נקראת PPP (Promoter-Proximal Pausing).
העצירה היא נקודת בקרה מרכזית:
- היא יכולה להימשך שניות, אך גם שעות.
- Pol II ממתין לסיגנל שיקבע אם להמשיך בשעתוק.
- אם הסיגנל המתאים אינו מגיע, Pol II יכול להתנתק והגן לא ישועתק במלואו.
Spt4/Spt5 ו־NELF
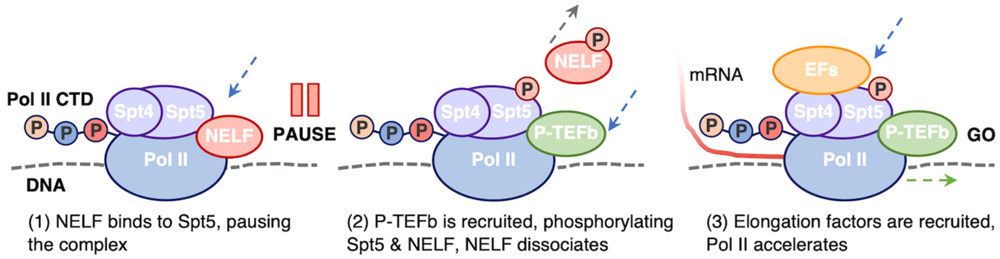
מיד לאחר initiation נקשרים ל־Pol II החלבונים Spt4 ו־Spt5, היוצרים יחד את קומפלקס DSIF.
- Spt5 מסייע בגיוס חלבוני capping אל ה־RNA החדש.
- NELF (
Negative Elongation Factor) מצטרף ויוצר את מצב העצירה. - כל עוד NELF נשאר בקומפלקס, Pol II אינו מתקדם ל־productive elongation.
P-TEFb ושחרור העצירה
כאשר מתקבל הסיגנל המתאים, P-TEFb מזרחן את:
- NELF
- Spt5
בעקבות הפוספורילציה:
- NELF מתנתק.
- Spt5 נשאר קשור ל־Pol II.
- Spt5 המזורחן מגייס elongation factors נוספים.
- Pol II יוצא מה־pause ונכנס ל־productive elongation.
בחלון העצירה מתווסף ה־5′ cap, וגם פקטורי splicing מתחילים להיקשר לתעתיק. לכן PPP הוא checkpoint שמחבר בין ההחלטה להמשיך בשעתוק לבין תחילת עיבוד ה־RNA.
2. התארכות (Elongation) ועיבוד RNA תוך כדי שעתוק
לאחר השחרור מ־PPP, Pol II מתקדם לאורך הגן ומסנתז את ה־pre-mRNA. התהליך אינו אחיד: גורמים שונים יכולים להאיץ את הפולימראז, להאט אותו או לגרום לו לעצור זמנית.
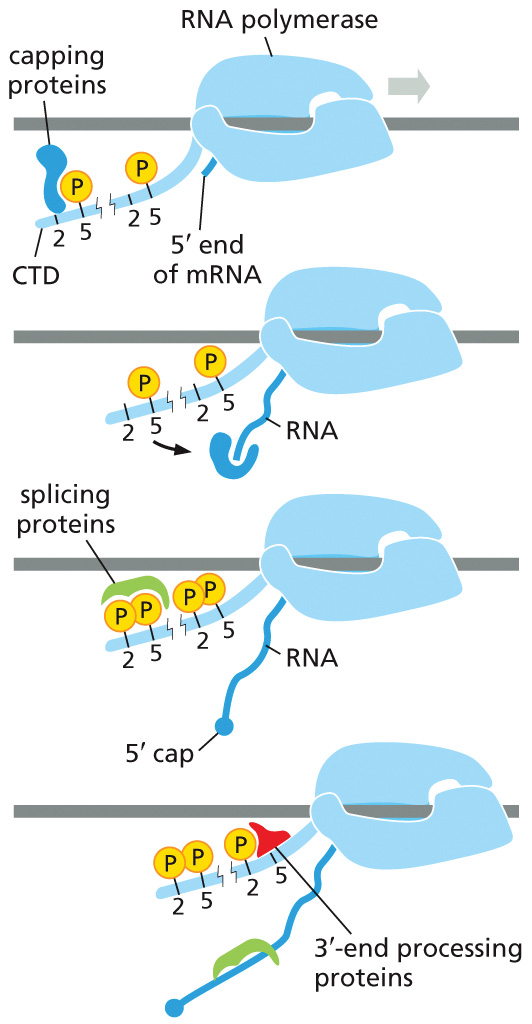
עיבוד ה־RNA מתרחש co-transcriptionally, כלומר בזמן שהתעתיק עדיין נוצר. שלושת תהליכי העיבוד המרכזיים הם:
- 5′ capping - לתחילת ה־RNA, כדי למנוע דגרדציה.
- Splicing - הוצאת אינטרונים וחיבור אקסונים.
- זנב פולי A - 3′-end processing ו־polyadenylation.
Capping
חלבוני capping נקשרים מוקדם, באזור ה־promoter-proximal pause, ומוסיפים 5′ cap לקצה ה־RNA. ה־cap מגן על התעתיק מפירוק מהיר.
Splicing
פקטורי splicing נקשרים ל־CTD ול־RNA בזמן שה־RNA מסונתז. בשיעור הודגשו הפוספורילציות של Ser2 ושל Thr4 כסימנים המסייעים לגיוס חלבוני splicing.
לכן splicing אינו אירוע שמתחיל רק לאחר סיום השעתוק: אינטרונים יכולים לצאת ואקסונים יכולים להתחבר כבר במהלך elongation.
Proofreading של Pol II
כאשר Pol II מכניס נוקלאוטיד שגוי, ההתאמה בין ה־RNA ל־DNA נפגעת. הפולימראז יכול לבצע backtracking:
- Pol II נע לאחור על גבי ה־DNA.
- הקצה המכיל את הנוקלאוטיד השגוי בולט אל מחוץ לאתר הפעיל.
- פקטור cleavage חותך את מקטע ה־RNA הקצר והשגוי.
- נוצר קצה RNA תקין חדש, ו־Pol II חוזר לסינתזה קדימה.
מה משפיע על מהירות ההתארכות?
- חלבונים הקשורים ל־DNA.
- נוקלאוזומים ומצב אריזת הכרומטין.
- מודיפיקציות של histones.
- מבנים שניוניים ב־DNA.
- elongation factors הקשורים ל־Pol II.
כל אחד מהגורמים האלה יכול לגרום ל־Pol II להתקדם מהר יותר, לאט יותר או לעצור. לכן גם קצב ההתארכות הוא שכבת בקרה על ביטוי גנים.
3. טרמינציה ו־Polyadenylation (או: PAS-CPA-XRN2)
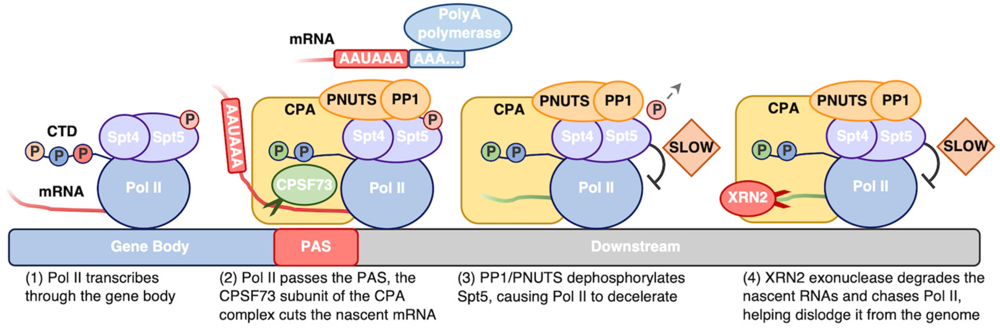
לקראת סוף הגן, Pol II משעתק את אזור ה־PAS (Polyadenylation Site). ב־RNA, השעתוק שלו הוא בדרך כלל: AAUAAA.
הסיגנל מאפשר לגייס את הקומפלקס CPA (Cleavage & Polyadenylation Complex), שחותך את ה־RNA ומוסיף לו זנב poly(A) (להלן).
CPA - חיתוך והוספת poly-A
CPA מבצע שתי פעולות מרכזיות, כפי שמרמז שמו:
- Cleavage - חיתוך ה־RNA החדש.
- Polyadenylation - סיוע ל־Poly(A) polymerase בהוספת זנב poly-A, לקצה 3׳ של התעתיק.
החיתוך יוצר את קצה ה־3׳ של ה־mRNA. לאחר הוספת poly-A, התעתיק יציב יותר ומוכן להמשך הבשלה ויציאה מהגרעין.
חשוב: חיתוך ה־RNA אינו גורם ל־Pol II לעצור מיד. הפולימראז ממשיך לשעתק downstream עוד בערך 2,000-500bp.
האטת Pol II ושחרור
לאחר החיתוך נקשרים פקטורי טרמינציה, ובהם PNUTS ו־PP1. הם מסירים את הפוספורילציה מ־Spt5, ולכן Pol II מאט. זה גם מאפשר חיבור של אנזים אקסונוקלאז בשם XRN2 (אקסונוקלאז - אוכל RNA, השם “XRN2” הופיע במצגת אבל למיטב זכרוני לא הוזכר בהרצאה).
ה־RNA שנוצר downstream מנקודת החיתוך חסר 5′ cap, ולכן חשוף לפירוק. האקסונוקלאז 5′←3′ מתחיל לפרק אותו ורודף אחרי Pol II. האקסונוקלאז משיג את הפולימראז המואט ומערער את קומפלקס השעתוק, עד ש־Pol II משתחרר מה־DNA.
לסיכום:
- PAS משועתק
- CPA חותך את ה־RNA ומסייע בהוספת poly-A
- PNUTS/PP1 מסירים פוספט מ־Spt5, ו־Pol II מאט
- אקסונוקלאז מפרק את ה־RNA הלא מוגן בכיוון 5′←3′
- ההאקסונוקלאז משיג את Pol II, מה שמוביל לטרמינציה.